УДФ-глюкозо-4-эпимераза
| УДФ-глюкозо-4-эпимераза | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | УДФгалактозо-4-эпимераза-4-эпимераза-дифосфат-глюкоза-4-эпимеразаУДФ-4-эпимеразаУДФ-4-эпимеразаУДФ-галактоза-4-эпимеразеуридин-дифосфоглюкоза эпимеразеуридин-дифосфо-галактозо-4-эпимеразаUDP-D-галактозо-4-эпимеразаUDP-глюкоза-эпимеразеуридин-дифосфоглюкоза 4-эпимеразеуридин-дифосфат-галактоза 4-эпимераза | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Генные карты : [1] ; ОМА : - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| УДФ-глюкозо-4-эпимераза | |||
|---|---|---|---|
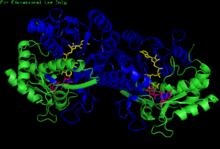 Гомодимер УДФ-глюкозо-4-эпимеразы H. sapiens, связанный с НАДН и УДФ-глюкозой . Домены: N-концевой и C-концевой . | |||
| Идентификаторы | |||
| Номер ЕС. | 5.1.3.2 | ||
| Номер CAS. | 9032-89-7 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| УДФ-галактозо-4-эпимераза | |||
|---|---|---|---|
 Человеческий GALE связан с NAD+ и UDP-GlcNAc , с выделенными N- и C-концевыми доменами. Asn 207 искажается для размещения UDP-GlcNAc внутри активного сайта. | |||
| Идентификаторы | |||
| Символ | ГЕЙЛ | ||
| ген NCBI | 2582 | ||
| HGNC | 4116 | ||
| МОЙ БОГ | 606953 | ||
| RefSeq | НМ_000403 | ||
| ЮниПрот | Q14376 | ||
| Другие данные | |||
| Номер ЕС | 5.1.3.2 | ||
| Локус | Хр. 1 п36-п35 | ||
| |||
| НАД-зависимая эпимераза/дегидратаза | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ? | ||
| Пфам | PF01370 | ||
| ИнтерПро | ИПР001509 | ||
| Мембраном | 330 | ||
| |||
Фермент GALE УДФ-глюкозо-4-эпимераза ( EC 5.1.3.2 ), также известный как УДФ-галактозо-4-эпимераза или , представляет собой гомодимерную эпимеразу, обнаруженную в клетках бактерий, грибов, растений и млекопитающих. Этот фермент выполняет заключительный этап Лелуара метаболизма галактозы пути , катализируя обратимое превращение УДФ-галактозы в УДФ-глюкозу . [1] GALE прочно связывает никотинамидадениндинуклеотид (НАД+), кофактор, необходимый для каталитической активности. [2]
Кроме того, человеческие и некоторые бактериальные изоформы GALE обратимо катализируют образование UDP -N -ацетилгалактозамина (UDP-GalNAc) из UDP -N -ацетилглюкозамина ( UDP-GlcNAc ) в присутствии НАД+, что является начальной стадией синтеза гликопротеинов или гликолипидов . [3]
Историческое значение
[ редактировать ]Доктор Луис Лелуар установил роль GALE в метаболизме галактозы во время своей работы в Институте биохимических исследований Фонда Кампомар, первоначально уничтожив фермент вальденазу. [4] Доктор Лелуар был удостоен Нобелевской премии по химии 1970 года за открытие сахарных нуклеотидов и их роли в биосинтезе углеводов. [5]
Структура
[ редактировать ]GALE принадлежит к суперсемейству белков короткоцепочечных дегидрогеназ/редуктаз (SDR). [6] Это семейство характеризуется консервативным мотивом Tyr-XXX-Lys, необходимым для ферментативной активности; одна или несколько складных подмостей Россмана ; и способность связывать НАД + . [6]
Третичная структура
[ редактировать ]Структура GALE определена для ряда видов, включая E. coli. [7] и люди. [8] GALE существует в виде гомодимера у различных видов. [8]
Хотя размер субъединицы варьируется от 68 аминокислот (Enterococcus faecalis) до 564 аминокислот (Rhodococcus jostii) , длина большинства субъединиц GALE составляет около 330 аминокислот. [6] Каждая субъединица содержит два отдельных домена. N-концевой домен содержит 7-нитевой параллельный β-складчатый лист, окруженный α-спиралями. [1] Парные складки Россмана внутри этого домена позволяют GALE прочно связывать один НАД. + кофактор на субъединицу. [2] 6-нитевой β-лист и 5 α-спиралей составляют С-концевой домен GALE. [1] С-концевые остатки связывают UDP, так что субъединица отвечает за правильное расположение UDP-глюкозы или UDP-галактозы для катализа. [1]
Активный сайт
[ редактировать ]фермента Щель между N- и C-концевыми доменами GALE составляет активный центр . Консервативный мотив Tyr-XXX Lys необходим для каталитической активности GALE; у человека этот мотив представлен Tyr 157-Gly-Lys-Ser-Lys 161, [6] в то время как E. coli GALE содержит Tyr 149-Gly-Lys-Ser-Lys 153. [8] Размер и форма активного сайта GALE различаются у разных видов, что позволяет варьировать специфичность субстрата GALE. [3] Кроме того, конформация активного сайта внутри видоспецифичного GALE податлива; например, объемная 2'-N-ацетильная группа UDP-GlcNAc размещается внутри активного сайта GALE человека за счет вращения карбоксамидной боковой цепи Asn 207. [3]
| Остаток | Функция |
|---|---|
| Ала 216, Фе 218 | Закрепите урациловое кольцо на ферменте. |
| Асп 295 | Взаимодействует с 2'-гидроксильной группой рибозы. |
| Асн 179, Арг 231, Арг 292 | Взаимодействуют с фосфатными группами УДФ. |
| Бык 299, Асн 179 | Взаимодействовать с 2'-гидроксильной группой галактозы или 6'-гидроксильной группой глюкозы; правильно расположить сахар в активном центре. |
| Тир 177, Фе 178 | Взаимодействовать с 3'-гидроксильной группой галактозы или 6'-гидроксильной группой глюкозы; правильно расположить сахар в активном центре. |
| Свет 153 | Снижает pKa Tyr 149, позволяет отщеплять или отдавать атом водорода к 4'-гидроксильной группе сахара или от нее. |
| Тир 149 | Абстрагирует или отдает атом водорода к 4'-гидроксильной группе сахара или от нее, катализируя образование промежуточного соединения 4-кетопиранозы. |
Механизм
[ редактировать ]Превращение УДФ-галактозы в УДФ-глюкозу
[ редактировать ]GALE инвертирует конфигурацию 4'-гидроксильной группы УДФ-галактозы посредством серии из 4 стадий. При связывании УДФ-галактозы консервативный остаток тирозина в активном центре отрывает протон от 4'-гидроксильной группы. [7] [10]
присоединяется 4'-гидрид , образуя НАДН и промежуточное соединение 4-кетопиранозы. Одновременно с этим к si -стороне НАД+ [1] Промежуточное соединение 4-кетопиранозы поворачивается на 180 ° вокруг пирофосфорильной связи между гликозильным кислородом и атомом β-фосфора, представляя противоположную сторону промежуточного соединения кетопиранозы по отношению к НАДН. [10] Перенос гидрида от НАДН к этой противоположной стороне инвертирует стереохимию 4'-центра. Консервативный остаток тирозина затем отдает свой протон, регенерируя 4'-гидроксильную группу. [1]
Преобразование UDP-GlcNAc в UDP-GalNAc
[ редактировать ]Человеческие и некоторые бактериальные изоформы GALE обратимо катализируют превращение UDP-GlcNAc в UDP-GalNAc по идентичному механизму, инвертируя стереохимическую конфигурацию 4'-гидроксильной группы сахара. [3] [11]
Биологическая функция
[ редактировать ]
Метаболизм галактозы
[ редактировать ]Прямых катаболических путей метаболизма галактозы не существует. Поэтому галактоза преимущественно превращается в глюкозо-1-фосфат , который может быть направлен на гликолиз или путь синтеза инозитола . [12]
GALE действует как один из четырех ферментов пути Лелуара превращения галактозы в глюкозо-1-фосфат. Сначала галактозомутаротаза превращает β-D-галактозу в α-D-галактозу. [1] Затем галактокиназа фосфорилирует α-D-галактозу по 1'-гидроксильной группе, образуя галактозо-1-фосфат . [1] На третьем этапе галактозо-1-фосфатуридилтрансфераза катализирует обратимый перенос фрагмента UMP от УДФ-глюкозы к галактозо-1-фосфату, образуя УДФ-галактозу и глюкозо-1-фосфат. [1] На заключительном этапе Лелуара УДФ-глюкоза регенерируется из УДФ-галактозы с помощью GALE; УДФ-глюкоза возвращается к третьему этапу пути. [1] Таким образом, GALE регенерирует субстрат, необходимый для продолжения цикла пути Лелуара.
Глюкозо-1-фосфат, образующийся на этапе 3 пути Лелуара, может быть изомеризован в глюкозо-6-фосфат под действием фосфоглюкомутазы . Глюкозо-6-фосфат легко вступает в гликолиз, приводя к образованию АТФ и пирувата. [13] Кроме того, глюкозо-6-фосфат может превращаться в инозитол-1-фосфат под действием инозитол-3-фосфатсинтазы , образуя предшественник, необходимый для биосинтеза инозитола . [14]
Синтез UDP-GalNAc
[ редактировать ]Человеческие и отдельные бактериальные изоформы GALE связывают UDP-GlcNAc, обратимо катализируя его превращение в UDP-GalNAc. Семейство гликозилтрансфераз, известное как UDP -N -ацетилгалактозамин:полипептидные N-ацетилгалактозаминтрансферазы (ppGaNTases), переносит GalNAc от UDP-GalNAc к остаткам серина и треонина гликопротеина. [15] Гликозилирование, опосредованное ppGaNTase, регулирует сортировку белков, [16] [17] [18] [19] [20] лигандная сигнализация, [21] [22] [23] устойчивость к протеолитической атаке, [24] [25] и представляет собой первый решительный шаг в биосинтезе муцина. [15]
Роль в болезни
[ редактировать ]Дефицит или дисфункция GALE у человека приводит к галактоземии III типа , которая может существовать в легкой (периферической) или более тяжелой (генерализованной) форме. [12]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к Холден Х.М., Рэймент I, Тоден Дж.Б. (ноябрь 2003 г.). «Структура и функции ферментов пути Лелуара метаболизма галактозы» . Ж. Биол. Хим . 278 (45): 43885–8. дои : 10.1074/jbc.R300025200 . ПМИД 12923184 .
- ^ Перейти обратно: а б Лю Ю., Ванхук Дж.Л., Фрей П.А. (июнь 1996 г.). «UDP-галактозо-4-эпимераза: содержание НАД + и полоса переноса заряда, связанная с конформационным переходом, индуцированным субстратом». Биохимия . 35 (23): 7615–20. дои : 10.1021/bi960102v . ПМИД 8652544 .
- ^ Перейти обратно: а б с д Тоден Дж.Б., Волерс Т.М., Фридович-Кейл Дж.Л., Холден Х.М. (май 2001 г.). «Человеческая УДФ-галактозо-4-эпимераза. Размещение УДФ-N-ацетилглюкозамина в активном центре» . Ж. Биол. Хим . 276 (18): 15131–6. дои : 10.1074/jbc.M100220200 . ПМИД 11279032 .
- ^ ЛЕЛУАР Л.Ф. (сентябрь 1951 г.). «Ферментативное превращение уридиндифосфатглюкозы в производное галактозы». Арх Биохим . 33 (2): 186–90. дои : 10.1016/0003-9861(51)90096-3 . HDL : 11336/140700 . ПМИД 14885999 .
- ^ «Нобелевская премия по химии 1970 г.» (Пресс-релиз). Шведская королевская академия наук. 1970 год . Проверено 17 мая 2010 г.
- ^ Перейти обратно: а б с д Кавана К.Л., Йорнвалль Х., Перссон Б., Опперманн У. (декабрь 2008 г.). «Семейства генов и белков дегидрогеназы / редуктазы со средней и короткой цепью: суперсемейство SDR: функциональное и структурное разнообразие внутри семейства метаболических и регуляторных ферментов» . Клетка. Мол. Наука о жизни . 65 (24): 3895–906. дои : 10.1007/s00018-008-8588-y . ПМЦ 2792337 . ПМИД 19011750 .
- ^ Перейти обратно: а б PDB : 1EK5 ; Тоден Дж.Б., Волерс Т.М., Фридович-Кейл Дж.Л., Холден Х.М. (май 2000 г.). «Кристаллографические доказательства того, что Tyr 157 функционирует как основа активного центра в UDP-галактозо-4-эпимеразе человека». Биохимия . 39 (19): 5691–701. дои : 10.1021/bi000215l . ПМИД 10801319 .
- ^ Перейти обратно: а б с PDB : 1XEL ; Тоден Дж.Б., Фрей П.А., Холден Х.М. (апрель 1996 г.). «Молекулярная структура НАДН/УДФ-глюкоза абортивного комплекса УДФ-галактозо-4-эпимеразы из Escherichia coli: значение для каталитического механизма». Биохимия . 35 (16): 5137–44. дои : 10.1021/bi9601114 . ПМИД 8611497 .
- ^ PDB : 1A9Z ; Тоден Дж. Б., Холден Х. М. (август 1998 г.). «Драматические различия в связывании УДФ-галактозы и УДФ-глюкозы с УДФ-галактозо-4-эпимеразой из Escherichia coli». Биохимия . 37 (33): 11469–77. дои : 10.1021/bi9808969 . ПМИД 9708982 .
- ^ Перейти обратно: а б Лю Ю, Тоден Дж. Б., Ким Дж., Бергер Э., Гулик А. М., Ружичка Ф. Дж., Холден Х. М., Фрей П. А. (сентябрь 1997 г.). «Механистическая роль тирозина 149 и серина 124 в УДФ-галактозо-4-эпимеразе Escherichia coli». Биохимия . 36 (35): 10675–84. дои : 10.1021/bi970430a . ПМИД 9271498 .
- ^ Кингсли Д.М., Козарский К.Ф., Хобби Л., Кригер М. (март 1986 г.). «Обратимые дефекты O-связанного гликозилирования и экспрессии рецептора ЛПНП у мутанта с дефицитом 4-эпимеразы UDP-Gal/UDP-GalNAc». Клетка . 44 (5): 749–59. дои : 10.1016/0092-8674(86)90841-X . ПМИД 3948246 . S2CID 28293937 .
- ^ Перейти обратно: а б Лай К., Эльзас Л.Дж., Виеренга К.Дж. (ноябрь 2009 г.). «Галактозная токсичность у животных» . ИУБМБ Жизнь . 61 (11): 1063–74. дои : 10.1002/iub.262 . ПМК 2788023 . ПМИД 19859980 .
- ^ Страйер, Люберт; Берг, Джереми Марк; Тимочко, Джон Л. (2008). Биохимия (вкладыш) . Сан-Франциско: WH Freeman. стр. 443–58 . ISBN 9780716718437 .
- ^ Мичелл Р.Х. (февраль 2008 г.). «Производные инозитола: эволюция и функции». Нат. Преподобный мол. Клеточная Биол . 9 (2): 151–61. дои : 10.1038/nrm2334 . ПМИД 18216771 . S2CID 3245927 .
- ^ Перейти обратно: а б Тен Хаген К.Г., Фриц Т.А., Табак Л.А. (январь 2003 г.). «Все в семье: UDP-GalNAc:полипептид N-ацетилгалактозаминилтрансферазы» . Гликобиология . 13 (1): 1П–16П. дои : 10.1093/гликоб/cwg007 . ПМИД 12634319 .
- ^ Альфала М., Джейкоб Р., Пройсс У., Циммер К.П., Наим Х., Наим Х.Ю. (июнь 1999 г.). «О-связанные гликаны опосредуют апикальную сортировку сахарозы-изомальтазы кишечника человека посредством ассоциации с липидными рафтами» . Курс. Биол . 9 (11): 593–6. дои : 10.1016/S0960-9822(99)80263-2 . ПМИД 10359703 . S2CID 16866875 .
- ^ Альтшулер Ю., Кинлоу К.Л., Польша, Пенсильвания, Брунс Дж.Б., Аподака Г., Вайс О.А., Хьюи Р.П. (март 2000 г.). «Клатрин-опосредованный эндоцитоз MUC1 модулируется состоянием его гликозилирования» . Мол. Биол. Клетка . 11 (3): 819–31. дои : 10.1091/mbc.11.3.819 . ПМК 14813 . ПМИД 10712502 .
- ^ Бреуза Л., Гарсия М., Дельгросси М.Х., Ле Бивик А. (февраль 2002 г.). «Роль мембранно-проксимального сайта O-гликозилирования в сортировке человеческого рецептора нейротрофинов на апикальной мембране клеток MDCK». Эксп. Сотовый Res . 273 (2): 178–86. дои : 10.1006/excr.2001.5442 . ПМИД 11822873 .
- ^ Наим Х.И., Джоберти Дж., Альфала М., Джейкоб Р. (июнь 1999 г.). «Временная ассоциация событий N- и O-связанного гликозилирования и их влияние на поляризованную сортировку кишечной щеточной границы сахаразы-изомальтазы, аминопептидазы N и дипептидилпептидазы IV» . Ж. Биол. Хим . 274 (25): 17961–7. дои : 10.1074/jbc.274.25.17961 . PMID 10364244 .
- ^ Чжэн X, Сэдлер Дж. Э. (март 2002 г.). «Муцин-подобный домен энтеропептидазы направляет апикальное нацеливание в клетки почек собак Мадина-Дарби» . Ж. Биол. Хим . 277 (9): 6858–63. дои : 10.1074/jbc.M109857200 . ПМИД 11878264 .
- ^ Хупер Л.В., Гордон Дж.И. (февраль 2001 г.). «Гликаны как законодатели взаимодействий хозяин-микроб: от симбиоза до патогенности» . Гликобиология . 11 (2): 1П–10П. дои : 10.1093/гликоб/11.2.1R . ПМИД 11287395 .
- ^ Йе Дж.С., Хираока Н., Петриняк Б., Накаяма Дж., Эллис Л.Г., Рабука Д., Хиндсгаул О., Март Дж.Д., Лоу Дж.Б., Фукуда М. (июнь 2001 г.). «Новые сульфатированные рецепторы самонаведения лимфоцитов и их контроль с помощью бета-1,3-N-ацетилглюкозаминилтрансферазы расширения Core1» . Клетка . 105 (7): 957–69. дои : 10.1016/S0092-8674(01)00394-4 . ПМИД 11439191 . S2CID 18674112 .
- ^ Сомерс В.С., Тан Дж., Шоу Г.Д., Кампхаузен Р.Т. (октябрь 2000 г.). «Понимание молекулярной основы связывания и свертывания лейкоцитов, выявленное с помощью структур P- и E-селектина, связанных с SLe (X) и PSGL-1» . Клетка . 103 (3): 467–79. дои : 10.1016/S0092-8674(00)00138-0 . ПМИД 11081633 . S2CID 12719907 .
- ^ Зауэр Дж., Сигуршельд Б.В., Кристенсен У., Франдсен Т.П., Миргородская Е., Харрисон М., Ропсторф П., Свенссон Б. (декабрь 2000 г.). «Глюкоамилаза: взаимосвязи структура/функция и белковая инженерия». Биохим. Биофиз. Акта . 1543 (2): 275–293. дои : 10.1016/s0167-4838(00)00232-6 . ПМИД 11150611 .
- ^ Гарнер Б., Мерри А.Х., Ройл Л., Харви DJ, Радд П.М., Тиллет Дж. (июнь 2001 г.). «Структурное выяснение N- и O-гликанов аполипопротеина (а) человека: роль о-гликанов в обеспечении устойчивости к протеазам» . Ж. Биол. Хим . 276 (25): 22200–8. дои : 10.1074/jbc.M102150200 . ПМИД 11294842 .
Дальнейшее чтение
[ редактировать ]- Лелуар Л.Ф. (1953). «Ферментативная изомеризация и родственные процессы». Достижения энзимологии и смежных областей молекулярной биологии . Достижения в энзимологии и смежных областях молекулярной биологии. Том. 14. стр. 193–218. дои : 10.1002/9780470122594.ch6 . ISBN 9780470122594 . ПМИД 13057717 .
{{cite book}}:|journal=игнорируется ( помогите ) - Максвелл Э.С., де Робишон-Сульмайстер Х (1960). «Очистка уридиндифосфатгалактозо-4-эпимеразы из дрожжей и идентификация связанного с белком дифосфопиридинового нуклеотида» . Ж. Биол. Хим . 235 (2): 308–312. дои : 10.1016/S0021-9258(18)69520-1 .
- Уилсон Д.Б., Хогнесс Д.С. (август 1964 г.). «Ферменты галактозного оперона Escherichia coli. I Очистка и характеристика уридиндифосфогалактозо-4-эпимеразы» . Ж. Биол. Хим . 239 : 2469–81. дои : 10.1016/S0021-9258(18)93876-7 . ПМИД 14235524 .
Внешние ссылки
[ редактировать ]- Запись GeneReviews/NCBI/NIH/UW о галактоземии, вызванной дефицитом эпимеразы
- Записи OMIM о галактоземии с дефицитом эпимеразы
- УДФгалактоза + 4-эпимераза в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- ^ Биби, Джейн А.; Фрей, Перри А. (1 октября 1998 г.). «Галактозомутаротаза: очистка, характеристика и исследование двух важных остатков гистидина» . Биохимия . 37 (42): 14989–14997. дои : 10.1021/bi9816047 . ISSN 0006-2960 .