Кооперативность
Эта статья нуждается в дополнительных цитатах для проверки . ( декабрь 2009 г. ) |
Кооперативность — это явление, проявляющееся в системах, включающих идентичные или почти идентичные элементы, которые действуют независимо друг от друга, по сравнению с гипотетической стандартной невзаимодействующей системой, в которой отдельные элементы действуют независимо. Одним из проявлений этого являются ферменты или рецепторы , которые имеют несколько сайтов связывания , где сродство сайтов связывания к лиганду очевидно увеличивается ( положительная кооперативность ) или снижается ( отрицательная кооперативность ) при связывании лиганда с сайтом связывания. Например, когда атом кислорода связывается с одним из четырех сайтов связывания гемоглобина, сродство к кислороду трех оставшихся доступных сайтов связывания увеличивается; т.е. кислород с большей вероятностью связывается с гемоглобином, связанным с одним кислородом, чем с несвязанным гемоглобином. Это называется кооперативным связыванием . [1]
Мы также наблюдаем кооперативность в молекулах с большой цепью, состоящих из множества идентичных (или почти идентичных) субъединиц (таких как ДНК , белки и фосфолипиды ), когда такие молекулы претерпевают фазовые переходы, такие как плавление, разворачивание или раскручивание. Это называется кооперативностью субъединиц. Однако определение кооперативности, основанное на кажущемся увеличении или уменьшении сродства к последовательным этапам связывания лиганда, проблематично, поскольку понятие «энергия» всегда должно определяться относительно стандартного состояния. Когда мы говорим, что аффинность увеличивается при связывании одного лиганда, эмпирически неясно, что мы имеем в виду, поскольку для строгого определения энергии связывания и, следовательно, также аффинности требуется кривая некооперативного связывания. Гораздо более общее и полезное определение позитивной кооперативности таково: процесс, включающий множество идентичных дополнительных шагов, в котором промежуточные состояния статистически недостаточно представлены по сравнению с гипотетической стандартной системой (нулевая гипотеза), где шаги происходят независимо друг от друга.
Аналогичным образом, определение негативной кооперативности будет означать процесс, включающий множество идентичных дополнительных шагов, в котором промежуточные состояния преобладают по сравнению с гипотетическим стандартным состоянием, в котором отдельные шаги происходят независимо. [2] Эти последние определения положительной и отрицательной кооперативности легко охватывают все процессы, которые мы называем «кооперативными», включая конформационные переходы в больших молекулах (таких как белки) и даже психологические явления большого числа людей (которые могут действовать независимо друг от друга или в кооперативный режим).
Кооперативное связывание [ править ]
Когда субстрат связывается с одной ферментативной субъединицей, остальные субъединицы стимулируются и становятся активными. Лиганды могут иметь положительную кооперативность, отрицательную кооперативность или некооперативность.
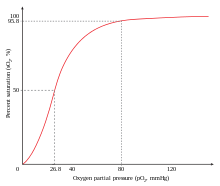
Примером положительной кооперативности является связывание кислорода с гемоглобином . Одна молекула кислорода может связываться с двухвалентным железом молекулы гема в каждой из четырех цепей молекулы гемоглобина . Дезоксигемоглобин имеет относительно низкое сродство к кислороду , но когда одна молекула связывается с одним гемом, сродство к кислороду увеличивается, позволяя второй молекуле связываться легче, а третьей и четвертой еще легче. Сродство к кислороду 3-оксигемоглобина примерно в 300 раз больше, чем у дезоксигемоглобина. Такое поведение приводит к тому, что кривая сродства гемоглобина становится сигмоидальной , а не гиперболической, как у мономерного миоглобина . По тому же процессу способность гемоглобина терять кислород увеличивается по мере того, как связывается меньше молекул кислорода. [1] См. также кривую диссоциации кислорода и гемоглобина .
Отрицательная кооперативность означает, что будет верно обратное; по мере того как лиганды связываются с белком , сродство белка к лиганду снижается, т. е. становится менее вероятным связывание лиганда с белком. Примером этого является взаимосвязь между глицеральдегид-3-фосфатом и ферментом глицеральдегид-3-фосфатдегидрогеназой.
Гомотропная кооперативность означает, что молекула, вызывающая кооперативность, будет затронута ею. Гетеротропная кооперативность – это когда третье вещество вызывает изменение сродства. Гомотропная или гетеротропная кооперативность может быть как положительной, так и отрицательной, в зависимости от того, поддерживает или препятствует она дальнейшему связыванию молекул лиганда с ферментами. [3]
Кооперативность подразделений [ править ]
Кооперативность — это не только явление связывания лигандов, но она также применяется в любое время, когда энергетические взаимодействия облегчают или затрудняют выполнение чего-либо с участием нескольких единиц, а не с использованием отдельных единиц. (То есть проще или сложнее по сравнению с тем, что ожидается, если учитывать только добавление нескольких единиц). Например, раскручивание ДНК предполагает кооперативность: части ДНК должны раскручиваться, чтобы ДНК могла осуществлять репликацию , транскрипцию и рекомбинацию . Положительная кооперативность между соседними нуклеотидами ДНК облегчает раскручивание целой группы соседних нуклеотидов, чем раскручивание того же числа нуклеотидов, разбросанных по цепи ДНК. Размер кооперативного подразделения — это количество соседних баз, которые имеют тенденцию распадаться как единое целое из-за эффектов положительного сотрудничества. Это явление применимо и к другим типам цепных молекул, например, к сворачиванию и разворачиванию белков, а также к «плавлению» фосфолипидных цепей, составляющих мембраны клеток . Кооперативность субъединиц измеряется по относительной шкале, известной как константа Хилла.
Уравнение Хилла [ править ]
Простой и широко используемой моделью молекулярных взаимодействий является уравнение Хилла , которое позволяет количественно оценить кооперативное связывание путем описания доли насыщенных сайтов связывания лигандов как функции концентрации лиганда.
Хилла Коэффициент
Коэффициент Хилла является мерой сверхчувствительности (т.е. насколько крутой является кривая отклика).
С эксплуатационной точки зрения коэффициент Хилла можно оценить как:
- .

где и — это входные значения, необходимые для получения 10% и 90% максимального ответа соответственно.


отклика Коэффициент
Меры глобальной чувствительности, такие как коэффициент Хилла, не характеризуют локальное поведение s-образных кривых. Вместо этого эти особенности хорошо фиксируются показателем коэффициента отклика. [4] определяется как:

В системной биологии такие реакции называются эластичностью .
между коэффициентом Хилла и отклика коэффициентом Связь
Альтшилер и др. (2017) показали, что эти показатели сверхчувствительности можно связать следующим уравнением: [5]

где обозначает среднее значение переменной x в диапазоне [a,b].

в составе функциональном Сверхчувствительность
Рассмотрим два связанных сверхчувствительных модуля, не учитывая эффекты секвестрации молекулярных компонентов между слоями. В этом случае выражение для кривой доза-реакция системы F является результатом математического состава функций: , которые описывают взаимосвязь ввода/вывода изолированных модулей. :



Браун и др. (1997) [6] [5] показали, что локальная сверхчувствительность разных слоев мультипликативно объединяется:
- .

В связи с этим результатом Феррелл и др. (1997) [7] показал для модулей типа Хилла, что общая глобальная сверхчувствительность каскада должна быть меньше или равна произведению оценок глобальной сверхчувствительности каждого слоя каскада, [5]
- ,

где и – коэффициент Хилла модулей 1 и 2 соответственно.


Альтшилер и др. (2017) [5] показали, что глобальную сверхчувствительность каскада можно рассчитать аналитически:

где и ограничил рабочий диапазон входа Хилла составной системы, т. е. входные значения для i-го слоя так, чтобы последний слой (соответствующий в данном случае) достигли 10% и 90% от максимального уровня выпуска. Следуя этому уравнению, коэффициент Хилла системы n можно было записать как произведение двух факторов: и , который характеризовал локальную среднюю чувствительность в соответствующей входной области для каждого слоя: , с в этом случае.





![{\displaystyle [X10_{i},X90_{i}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/2a6a9f701cc2d5bd5754f6206af5ce8e2b644e8f)
Для более общего случая каскада из N модулей коэффициент Хилла можно выразить как:
- ,

Супрамультипликативность [ править ]
Некоторые авторы сообщили о существовании супрамультипликативного поведения в сигнальных каскадах. [8] [9] (т.е. сверхчувствительность комбинации слоев выше, чем произведение индивидуальных сверхчувствительность), но во многих случаях окончательное происхождение супрамультипликативности оставалось неясным. Альтшилер и др. (2017) [5] Фреймворк естественным образом предложил общий сценарий, в котором может иметь место супрамультипликативное поведение. Это могло произойти, когда для данного модуля соответствующий входной рабочий диапазон Хилла находился во входной области с локальной сверхчувствительностью, превышающей глобальную ультрачувствительность соответствующей кривой зависимости доза-реакция.
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б Уитфорд Д. (2005). Белки: строение и функции . Джон Уайли и сыновья. стр. 66–74.
- ^ Абелиович Х (июль 2005 г.). «Эмпирический принцип экстремума для коэффициента Хилла во взаимодействиях лиганд-белок, показывающий отрицательную кооперативность» . Биофизический журнал . 89 (1): 76–9. Бибкод : 2005BpJ....89...76A . doi : 10.1529/biophysj.105.060194 . ПМК 1366580 . ПМИД 15834004 .
- ^ Хусейн Р., Кумари И., Шарма С., Ахмед М., Хан Т.А., Ахтер Ю. (декабрь 2017 г.). «Каталитическое разнообразие и гомотропная аллостерия двух монооксигеназоподобных белков цитохрома P450 из Trichoderma brevicompactum». Журнал биологической неорганической химии . 22 (8): 1197–1209. дои : 10.1007/s00775-017-1496-6 . ПМИД 29018974 . S2CID 25685603 .
- ^ Холоденко Б.Н., Хук Дж.Б., Вестерхофф Х.В., Браун Г.К. (сентябрь 1997 г.). «Количественная оценка передачи информации через клеточные пути передачи сигналов» . Письма ФЭБС . 414 (2): 430–4. дои : 10.1016/S0014-5793(97)01018-1 . ПМИД 9315734 . S2CID 19466336 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Альтшилер Э., Вентура АС, Колман-Лернер А, Черноморец А (29 июня 2017 г.). «Возврат к сверхчувствительности в сигнальных каскадах: объединение оценок локальной и глобальной сверхчувствительности» . ПЛОС ОДИН . 12 (6): e0180083. arXiv : 1608.08007 . Бибкод : 2017PLoSO..1280083A . дои : 10.1371/journal.pone.0180083 . ПМЦ 5491127 . ПМИД 28662096 .
 Эта статья содержит цитаты из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0) .
Эта статья содержит цитаты из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0) . - ^ Браун Г.К., Хук Дж.Б., Холоденко Б.Н. (август 1997 г.). «Почему протеинкиназные каскады имеют более одного уровня?». Тенденции биохимических наук . 22 (8): 288. дои : 10.1016/s0968-0004(97)82216-5 . ПМИД 9270298 .
- ^ Феррелл Дж. Э. (август 1997 г.). «Как реакции становятся более похожими на переключатели по мере продвижения вниз по каскаду протеинкиназы». Тенденции биохимических наук . 22 (8): 288–9. дои : 10.1016/s0968-0004(97)82217-7 . ПМИД 9270299 .
- ^ Альтшилер Э., Вентура А., Колман-Лернер А., Черноморец А. (октябрь 2014 г.). «Влияние ограничений восходящего и нисходящего потока на сверхчувствительность модуля сигнализации» . Физическая биология . 11 (6): 066003. Бибкод : 2014PhBio..11f6003A . дои : 10.1088/1478-3975/11/6/066003 . ПМЦ 4233326 . ПМИД 25313165 .
- ^ Рац Э., Слепченко Б.М. (июль 2008 г.). «Об усилении чувствительности внутриклеточных сигнальных каскадов» . Физическая биология . 5 (3): 036004. Бибкод : 2008PhBio...5c6004R . дои : 10.1088/1478-3975/5/3/036004 . ПМК 2675913 . ПМИД 18663279 .