Энойл-КоА-изомераза
| Д 3 -Д 2 -Эноил-КоА-изомераза | |||
|---|---|---|---|
 Тример 3,2-транс-еноил-КоА-изомеразы, человека | |||
| Идентификаторы | |||
| Номер ЕС. | 5.3.3.8 | ||
| Номер CAS. | 62213-29-0 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Эноил-КоА-(∆) изомераза ( EC 5.3.3.8 , также известная как додеценоил-КоА-(∆) изомераза , 3,2-транс-еноил-КоА-изомераза , ∆3(цис),∆2(транс)-еноил -КоА-изомераза , или ацетилен-аллен-изомераза , [1] представляет собой фермент , который катализирует превращение цис- или транс - двойных связей , связанных с коферментом А (КоА), жирных кислот у гамма- углерода (положение 3) в транс- двойные связи у бета- углерода (положение 2), как показано ниже:
![]()
Этот фермент играет важную роль в метаболизме ненасыщенных жирных кислот при бета-окислении .
Механизм
[ редактировать ]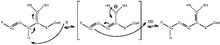
Эноил-КоА- изомераза участвует в бета-окислении , одном из наиболее часто используемых путей деградации жирных кислот , ненасыщенных жирных кислот с двойными связями в нечетных положениях углерода . [2] Это достигается путем смещения положения двойных связей ацил-КоА в промежуточных соединениях и превращения 3-цис- или транс-еноил-КоА в 2-транс-еноил-КоА. Поскольку ключевой этап деградации жирных кислот с двойными связями в четных положениях углерода также приводит к образованию 3-транс-еноил-КоА у млекопитающих и дрожжей , еноил-КоА- изомераза технически необходима для их метаболизма . также [3] Механизм реакции подробно показан на рисунке 1. [4] а основание , инициирующее изомеризацию , и группы NH, стабилизирующие интермедиат, расположены в активном центре еноил-КоА -изомеразы .
функционирует на стадии, непосредственно предшествующей фактическому бета-окислению , и образует двойную связь, идущую от бета - углерода (положение 2), Поскольку еноил-КоА- изомераза она участвует как в НАДФН -зависимом, так и в НАДФН -независимом путях бета-окисления. . [5] Двойная связь служит мишенью окисления и углерод - углерод разрыва связи , тем самым укорачивая цепь жирной кислоты .
Подклассификация
[ редактировать ]Эноил-КоА -изомеразы можно разделить на три класса:
- монофункциональный митохондриальный
- монофункциональный пероксисомальный
- многофункциональный
Монофункциональные митохондриальные и пероксисомальные ферменты обнаружены в и пероксисомах эукариот соответственно митохондриях . Многофункциональные ферменты обнаружены у бактерий и в пероксисомах некоторых эукариот , но они выполняют две функции: N-концевой домен действует так же, как другие классы еноил-КоА -изомераз , а С-концевой домен действует как дегидрогеназа , в частности , до 3-гидроксиактил-КоА. [4] еноил-Со-А-изомеразы выделяют два отдела Среди митохондриальной : короткоцепочечный и длинноцепочечный [4]. [6] При иммуноблоте антитела были протестированы против всех еноил-КоА-изомеразы. Однако две из этих изомераз имели присоединение антител : изомераза с короткой цепью и пероксисомальный многофункциональный фермент. [6] Был один фермент , который не обладал специфичностью связывания с этим антителом : митохондриальная длинноцепочечная изомераза. Длинноцепочечная изомераза была обнаружена, когда она элюировалась при более низкой концентрации фосфата калия в градиенте. [6] [7] Таким образом, было сделано открытие трех подклассов еноил-КоА-изомеразы.
Хотя все три класса ферментов последовательности мало перекрываются выполняют одну и ту же функцию, их аминокислотные . Например, только 40 из 302 аминокислотных одинаковы у монофункциональных пероксисомальных и митохондриальных ферментов человека последовательностей (13% ) . [4] Фактически, у млекопитающих пероксисомальный домен , фермент имеет дополнительный N-концевой которого нет в митохондриальном аналоге. [8] Кроме того, было обнаружено, что он является субъединицей пероксисомального трифункционального фермента (пТФЭ) и способствует лишь незначительному расщеплению цепи жирной кислоты . В этом смысле для многих высших организмов митохондриальный фермент необходим для получения максимальной энергии из липидов и питания мышц . [9]

Митохондрии (как короткоцепочечные, так и длинноцепочечные) печени крысы содержат более одной еноил-Ко-А-изомеразы. [10] Чтобы еще раз поддержать идею о том, что изомеразы с короткой и длинной цепью элюируются при разных концентрациях фосфата калия, следует отметить, что они не имеют схожей первичной полипептидной структуры, следовательно, они не должны быть эволюционно родственными. [6] [11] Пероксисомы растений и крысы печени сильно различаются по принципу действия. Несмотря на сходство первичной структуры , между разными экземплярами существуют различия. Начнем с того, что пероксисомы крыс печени представляют собой многофункциональный фермент , включающий еноил-КоА-изомеразу, еноил-КоА -гидратазу и L-(-)-3-гидроксиацил-КоА- дегидрогеназу . [12] три разных фермента, В этом объекте (многофункциональном белке) находятся позволяющие этому ферменту выполнять изомеризацию, гидратацию и дегидратацию. [13] [14] Изомеразная активность многофункционального фермента проявляется на аминоконцевой каталитической половине белка наряду с гидратазной активностью. [15] Дегидрогеназная активность еноил - КоА проявляется на карбоксильном конце. [15] При дальнейшем исследовании сайта связывания КоА на аминоконцевой половине многофункционального белка КоА субстрат не переносится через водную фазу из фазы изомеризации в место гидратации или не имеет объемной фазы. [11] [16] Это устраняет необходимость в ферменте , переносящем субстрат . [17] С другой стороны, семядоли превращают длинноцепочечный 3-транс-еноил-КоА, длинноцепочечный 3-цис-еноил-КоА и короткоцепочечный 3-цис-еноил-КоА в их 2-транс-еноил-КоА. Соответствующие формы CoA. [13] Как упоминалось ранее, растительная исключительно 2-транс -изомер еноил-КоА-изомераза образует в качестве продукта . Он не действует на виды 4-цис-еноил-КоА или виды 2-транс-4-транс-диеноил-КоА. [13] При сравнении продуктов растительной пероксисомы и многофункционального фермента печени крысы растение не обладает гидратазной активностью. [13] Растительная форма не образовывала 2-цис-изомер (из еноил-КоА-гидратазы) или D- или L-3-гидроксипроизводное (L-(-)-3-гидроксиацил-КоА-дегидрогеназа): продукты многофункционального фермента печени крысы. . [13] Скорость оборота этих двух подразделений пероксисом очень различна. Соотношение Kcat/Km в семядолях составляет 10^6 М-1с-1, что превосходит соотношение 0,07 * 10^6 М-1с-1. [13] Из-за высокой скорости оборота пероксисомы растений содержат меньшее количество еноил-КоА-изомеразы, чем их аналоги в печени крыс . [13]
В печени крысы митохондриальная еноил-КоА-изомераза и пероксисомальная еноил-КоА-изомераза, встроенные в многофункциональный фермент, имеют сходство в последовательности первичной структуры. [15] При сравнении аминоконцевой половины E. coli с аминоконцевой половиной печени крысы наблюдалось сходство первичной и вторичной структуры ближе к середине аминоконцевого конца. [15] Эта консервативная область должна быть важна для структуры и функции этого специфического фермента, поскольку она одинаково проявляется как в E. coli крыс , так и в печени . [15] [18]
Структура
[ редактировать ]Все классы еноил-КоА -изомераз принадлежат к семейству ферментов , суперсемейству гидратазы / изомеразы или кротоназы , и при исследовании с помощью рентгеновской кристаллографии обнаруживают общую структурную особенность семейства - N-концевое ядро со спиральной складкой, состоящей из состоит из четырех витков, каждый виток состоит из двух бета-листов и одной альфа-спирали . [19]
В еноил-КоА - изомеразе два бета-листа являются частью каталитического сайта , поскольку группы остатков NH , следующие за бета-листами, присоединяются к карбонильному кислороду ацил-КоА промежуточного соединения . Образование этой оксианионной дырки стабилизирует переходное состояние реакции , катализируемой ферментом . [4]

Более того, глутамата остаток , расположенный рядом с полостями тела, заполненными молекулами воды и выстланными гидрофобными или аполярными боковыми цепями, также был идентифицирован как часть каталитического сайта . В своей депротонированной форме глутамат может действовать как основание и удалять протон ацил-КоА из промежуточного соединения . Полости тела помогают перестроить глутамата боковую цепь , чтобы сохранить протон и позже доставить его обратно в ацил-КоА в другом положении углерода . [4]

NH-содержащие остатки были идентифицированы как Ala70 и Leu126, а - как Glu158 в пероксисомальных ферментах вида дрожжевого глутамат Saccharomyces cerevisiae . Их относительное расположение на ферменте можно сравнить на рисунке 2. [4]
Ферменты собой суперсемейства гидратазы кротоназы / изомеразы или , обычно представляют тримерные диски димеризованные в гексамеры . Широкий диапазон их субстратно - ферментной специфичности обусловлен вариациями расстояний между тримерными дисками и их ориентацией. [20] Однако человека митохондриальная еноил-КоА- изомераза представляет собой тример и ориентирует хвост жирной кислоты в совершенно другом направлении, чем это наблюдается в гексамерах . [8] Тримерный . диск пероксисомальных ферментов Saccharomyces cerevisiae представлен на рисунке 3 [20]
История
[ редактировать ]Эноил-КоА -изомераза была впервые идентифицирована и очищена из крыс печени митохондрий в 1960-х и 1970-х годах с помощью гель-фильтрации и ионообменной хроматографии . [21] С тех пор все классы еноил-КоА- изомеразы , митохондриальные , пероксисомальные и многофункциональные, были идентифицированы у различных организмов, включая большее количество млекопитающих , растений и одноклеточных организмов.
К 1994 году, используя крысы еноил-СоА -изомеразы кДНК в качестве зонда гибридизации , человека еноил-КоА -изомеразы кДНК можно было секвенировать и клонировать . [2] В том же году был выделен сам белок, не по сродству к крысиным антителам или кДНК зондам . [3] но путем совместной очистки с трансферазой и глутатион S-трансферазами человека. [22]
В попытках человека изучить еноил-КоА -изомеразу детально митохондриальный фермент в печени млекопитающих был идентифицирован как потенциальный биологический маркер метаболических заболеваний из-за его повышенных уровней в дефектных клетках и связанных с дефектами кислот бета-окисления жирных человека , болезни [22] будет указано в следующем разделе.
Клиническое значение
[ редактировать ]У людей дефекты в механизме бета-окисления приводят к гипокетотической гипергликемии , симптому голодания , из -за неэффективного использования жирных кислот в качестве основного источника энергии . [9] генетическом уровне Установлено, что метаболическое заболевание находится на : у крыс без генов еноил-КоА -изомеразы также наблюдался высокий уровень глюкозы в крови . Более того, биологический маркер этого состояния мог быть идентифицирован, поскольку моча этих крыс содержала высокие концентрации ненасыщенных дикарбоновых кислот со средней длиной цепи , состояние, называемое дикарбоновой ацидурией. [9]
Более поздние исследования связывают инфекцию вируса гепатита С (ВГС) с дефектами деградации жирных кислот , в частности, с дефектами еноил-КоА- изомеразы . [23] ВГС является основной причиной хронического гепатита , цирроза печени и рака печени , им во всем мире страдают более 180 миллионов человек. [24] Из-за длительного латентного периода и вируса отсутствия существующих лекарств для избавления от вируса , [25] ВГС является серьезной проблемой, вызывающей больше смертей, чем ВИЧ/СПИД в Соединенных Штатах . [26] но ее угроза до сих пор не получает должного внимания. Необходимость в лечении гепатита С крайне важна, и, по словам Джона Уорда, директора отдела борьбы с гепатитами Центра по контролю и профилактике заболеваний (CDC) , оно может спасти до 120 000 жизней. [26]
Согласно профилированию белков в печени с биоптатах пациентов ВГС первоначально была обнаружена корреляция между дисфункциональными митохондриальными процессами, в том числе бета-окислением , и ВГС . [27] Фактически, липиды играют важную роль в репликации цикле ВГС , и в образцах « in vivo » от с ВГС пациентов множество липидов было обнаружено , которые помогают ВГС в поглощении вируса , репликации РНК и секреции из организма хозяина. клетки . Ферменты , которые регулируют метаболизм жирных кислот , включая еноил-КоА- изомеразу , также были активизированы аналогичным образом . [23] Методы подавления генов показали, что еноил-КоА -изомераза необходима для ВГС репликации РНК , и открыли пути остановки инфекции ВГС на внутриклеточном уровне. [23]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «ФЕРМЕНТ, запись 5.3.3.8» . Проверено 1 марта 2012 г.
- ^ Jump up to: а б Янссен У., Финк Т., Лихтер П., Стоффель В. (сентябрь 1994 г.). «Митохондриальная 3,2-транс-еноил-КоА-изомераза человека (DCI): структура гена и локализация на хромосоме 16p13.3». Геномика . 23 (1): 223–8. дои : 10.1006/geno.1994.1480 . ПМИД 7829074 .
- ^ Jump up to: а б Килпонен Ю.М., Хайринен Х.М., Рен М., Хилтунен Ю.К. (май 1994 г.). «Клонирование кДНК и аминокислотная последовательность митохондриальной дельта-3-дельта-2-еноил-КоА-изомеразы человека: сравнение человеческого фермента с его крысиным аналогом, митохондриальной короткоцепочечной изомеразой» . Биохимический журнал . 300 (1): 1–5. дои : 10.1042/bj3000001 . ПМЦ 1138113 . ПМИД 8198519 .
- ^ Jump up to: а б с д и ж Мурсула А.М., ван Аалтен Д.М., Хилтунен Дж.К., Виренга Р.К. (июнь 2001 г.). «Кристаллическая структура дельта (3)-дельта (2)-еноил-КоА-изомеразы». Дж. Мол. Биол . 309 (4): 845–53. дои : 10.1006/jmbi.2001.4671 . ПМИД 11399063 . S2CID 69172923 .
- ^ Луо М.Дж., Смеланд Т.Э., Шукри К., Шульц Х. (январь 1994 г.). «Дельта-3,5, дельта-2,4-диеноил-КоА-изомераза из митохондрий печени крысы. Очистка и характеристика нового фермента, участвующего в бета-окислении ненасыщенных жирных кислот» . Ж. Биол. Хим . 269 (4): 2384–8. дои : 10.1016/S0021-9258(17)41957-0 . ПМИД 8300563 .
- ^ Jump up to: а б с д Килпонен, Дж. М.; Палосаари, премьер-министр; Хилтунен, Дж. К. (1990). «Появление длинноцепочечной дельта-3,дельта-2-еноил-КоА-изомеразы в печени крыс» . Биохимический журнал . 269 (1): 223–226. дои : 10.1042/bj2690223 . ПМЦ 1131556 . ПМИД 2375752 .
- ^ Брайан В. Гейсбрехт; Дай Чжу; Керстин Шульц; Катя Нау; Джеймс С. Моррелл; Майкл Джерати; Хорст Шульц; Ральф Эрдманн; Стивен Дж. Гулд (1998). «Молекулярная характеристика дельта3, дельта2-еноил-КоА-изомеразы Saccharomyces cerevisiae» . Журнал биологической химии . 273 (50): 33184–33191. дои : 10.1074/jbc.273.50.33184 . ПМИД 9837886 .
- ^ Jump up to: а б Партанен С.Т., Новиков Д.К., Попов А.Н., Мурсула А.М., Хилтунен Ю.К., Веренга Р.К. (сентябрь 2004 г.). «Кристаллическая структура 1,3 А митохондриальной дельта3-дельта2-еноил-КоА-изомеразы человека демонстрирует новый способ связывания жирной ацильной группы». Дж. Мол. Биол . 342 (4): 1197–208. дои : 10.1016/j.jmb.2004.07.039 . ПМИД 15351645 .
- ^ Jump up to: а б с Янссен Ю, Стоффель В (май 2002 г.). «Нарушение митохондриального бета-окисления ненасыщенных жирных кислот у мышей с дефицитом 3,2-транс-еноил-КоА-изомеразы» . Ж. Биол. Хим . 277 (22): 19579–84. дои : 10.1074/jbc.M110993200 . ПМИД 11916962 .
- ^ Jump up to: а б Палосаари Премьер-министр; Хилтунен, Дж. К. (1991). «Очистка и характеристика пероксисомальной дельта2, дельта3-еноил-КоА-изомеразы растений, действующей на 3-цис-еноил-КоА и 3-транс-еноил-КоА» (PDF) . Евро. Дж. Биохим . 196 (3): 699–705. дои : 10.1111/j.1432-1033.1991.tb15868.x . ПМИД 2013292 .
- ^ Jump up to: а б Птиив М. Палосаари; Йоханна М. Килпонен; Райя Т. Сормуненн; Ильмо Э. Хассин; Дж. Калерво Хилтунен (1989). «Характеристика митохондриального изозима у крысы» (PDF) . Журнал биологической химии . 265 (6): 3347–3353. ПМИД 2154476 .
- ^ Герхард Мюллер-Ньюэн; Уве Янссен; Вильгельм Штоффель (1995). «Эноил-КоА-гидратаза и изомераза образуют суперсемейство с общим остатком глутамата в активном центре» . Евро. Дж. Биохим . 228 (1): 68–73. doi : 10.1111/j.1432-1033.1995.0068o.x . ПМИД 7883013 .
- ^ Jump up to: а б с д и ж г Палосаари П.М., Килпонен Дж.М., Сормунен Р.Т., Хасинен Э., Хилтунен Дж.К. (1990). «Дельта-3,дельта-2-еноил-КоА-изомеразы. Характеристика митохондриального изофермента крысы» . Ж. Биол. Хим . 265 (6): 3347–53. дои : 10.1016/S0021-9258(19)39773-X . ПМИД 2154476 .
- ^ Дунъянь Чжан; Вэньфэн Юй; Брайан В. Гейсбрехт; Стивен Дж. Гулд; Говард Спречер; Хорст Шульц (2002). «Функциональная характеристика дельта3, дельта2-еноил-КоА-изомераз из печени крысы» . Журнал биологической химии . 277 (11): 9127–9132. дои : 10.1074/jbc.m112228200 . ПМИД 11781327 .
- ^ Jump up to: а б с д и Паиви М. Палосаари; Мауно Вихинен; Пекка 1. Манцалаг; Стефан Э. Х. Алексонилл; Тайна Пихлажаниеми; Дж. Калерво Хилтунен (1991). «Сходство аминокислотных последовательностей митохондриальной короткоцепочечной дельта3, дельта2-еноил-КоА-изомеразы и пероксисомальной многофункциональной дельта3, дельта2-еноил-КоА-изомеразы, 2-еноил-КоА-гидратазы, 3-гидроксиацил-КоА-дегидрогеназы в печени крысы» (PDF ) ) . Журнал биологической химии . 266 (17): 10750–10753. дои : 10.1016/S0021-9258(18)99081-2 . ПМИД 2040594 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Патрисия К. Бэббит; Джордж Л. Кеньон (1992). «Происхождение 4-хлорбензоатдегалогеназы: анализ идентичности аминокислотных последовательностей среди семейств ацил: адениллигаз, еноил-КоА-гидратаз/изомераз и ацил-КоА-тиоэстераз». Биохимия . 31 (24): 5594–5604. дои : 10.1021/bi00139a024 . ПМИД 1351742 .
- ^ Только г-н Мурсула; Затем М.Ф. ван Аалтен; Дж. Калерво Хилтунен; Рик К. Веренга (2001). «Кристаллическая структура дельта3-дельта2-еноил-КоА-изомеразы». Молекулярная биология . 309 (4): 845–853. дои : 10.1006/jmbi.2001.4671 . ПМИД 11399063 . S2CID 69172923 .
- ^ Анер Гурвиц; Ану М. Мурсула; Андреас Фирцингер; Барбара Гамильтон; Сеппо Х. Килпела инен; Андреас Хартиг; Хельмут Руис; Й. Калерво Хилтунен; Ханспетер Роттенштайнер (1998). «Пероксисомальная дельта3-цис-дельта2-транс-еноил-КоА-изомераза, кодируемая ECI1, необходима для роста дрожжей Saccharomyces cerevisiae на ненасыщенных жирных кислотах» . Журнал биологической химии . 273 (47): 31366–31374. дои : 10.1074/jbc.273.47.31366 . ПМИД 9813046 .
- ^ Гурвиц А., Мурсула А.М., Фирцингер А. и др. (ноябрь 1998 г.). «Пероксисомальная дельта3-цис-дельта2-транс-еноил-КоА-изомераза, кодируемая ECI1, необходима для роста дрожжей Saccharomyces cerevisiae на ненасыщенных жирных кислотах» . Ж. Биол. Хим . 273 (47): 31366–74. дои : 10.1074/jbc.273.47.31366 . ПМИД 9813046 .
- ^ Jump up to: а б Мурсула А.М., Хилтунен Ю.К., Виеренга Р.К. (январь 2004 г.). «Структурные исследования дельта (3)-дельта (2)-еноил-КоА-изомеразы: переменный способ сборки тримерных дисков суперсемейства кротоназ» . ФЭБС Летт . 557 (1–3): 81–7. дои : 10.1016/S0014-5793(03)01450-9 . ПМИД 14741345 .
- ^ Стоффель В., Грол М. (декабрь 1978 г.). «Очистка и свойства 3-цис-2-транс-еноил-КоА-изомеразы (додеценоил-КоА-дельта-изомеразы) из митохондрий печени крысы». З. Физиол Хоппе-Зейлера. Хим . 359 (12): 1777–82. дои : 10.1515/bchm2.1978.359.2.1777 . ПМИД 738702 .
- ^ Jump up to: а б Такахаси Ю., Хирата Ю., Бурштейн Ю., Листовски И. (декабрь 1994 г.). «Дельта-3, дельта-2-еноил-КоА-изомераза представляет собой белок, который очищается совместно с глутатион-S-трансферазами человека из аффинных матриц S-гексилглутатион» . Биохимический журнал . 304 (3): 849–52. дои : 10.1042/bj3040849 . ПМК 1137411 . ПМИД 7818490 .
- ^ Jump up to: а б с Расмуссен А.Л., Даймонд Д.Л., МакДермотт Дж.Э. и др. (ноябрь 2011 г.). «Системная вирусология идентифицирует митохондриальный фермент окисления жирных кислот, додеценоил-коэнзим А дельта-изомеразу, необходимый для репликации вируса гепатита С и вероятного патогенеза» . Дж. Вирол . 85 (22): 11646–54. дои : 10.1128/JVI.05605-11 . ПМК 3209311 . ПМИД 21917952 .
- ^ Розен, Хьюго Р. (июнь 2011 г.). «Хронический гепатит С». Медицинский журнал Новой Англии . 364 (25): 2429–2438. дои : 10.1056/NEJMcp1006613 . ПМИД 21696309 . S2CID 19755395 .
- ^ Амемия Ф., Маэкава С., Итакура Ю. и др. (февраль 2008 г.). «Нацеливание на липидный обмен при лечении инфекции, вызванной вирусом гепатита С» . Дж. Заразить. Дис . 197 (3): 361–70. дои : 10.1086/525287 . PMID 18248300 .
- ^ Jump up to: а б «Гепатит С убивает больше американцев, чем ВИЧ/СПИД» . Голос Америки, Здоровье . 27 февраля 2012 года . Проверено 3 марта 2012 г.
- ^ Даймонд Д.Л., Джейкобс Дж.М., Папер Б. и др. (сентябрь 2007 г.). «Протеомное профилирование биопсии печени человека: фиброз, индуцированный вирусом гепатита С, и митохондриальная дисфункция» . Гепатология . 46 (3): 649–57. дои : 10.1002/hep.21751 . ПМИД 17654742 .