На представленных схемах показано, как синтезируются жирные кислоты у микроорганизмов, и перечислены ферменты, обнаруженные в кишечной палочке . [2] Эти реакции выполняются синтетазой жирных кислот II (FASII), которая обычно содержит несколько ферментов, действующих как один комплекс. FASII присутствует в прокариотах , растениях, грибах и паразитах, а также в митохондриях . [4]
У животных, а также у некоторых грибов, таких как дрожжи, те же реакции происходят с синтазой жирных кислот I (FASI), большим димерным белком, который обладает всей ферментативной активностью, необходимой для создания жирной кислоты. FASII менее эффективен, чем FASI; однако это позволяет образовывать больше молекул, включая жирные кислоты со «средней цепью», за счет раннего обрыва цепи. [4]
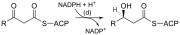
После образования жирная кислота с соотношением углерода 16:0 может претерпевать ряд модификаций, приводящих к десатурации и/или элонгации . Элонгация до стеарата (18:0) в основном происходит в ЭР под действием нескольких мембраносвязанных ферментов. Шаги, участвующие в процессе элонгации, в основном такие же, как и те, которые выполняются с помощью FAS, но четыре основных последовательных этапа элонгации выполняются отдельными белками, которые могут быть физически связаны. [5] [6]
При синтезе жиров восстановителем является НАДФН , тогда как НАД является окислителем при бета-окислении (расщеплении жирных кислот до ацетил-КоА). Это различие иллюстрирует общий принцип, согласно которому НАДФН расходуется во время реакций биосинтеза, тогда как НАДН генерируется в реакциях с выделением энергии. [7] (Таким образом, НАДФН также необходим для синтеза холестерина из ацетил-КоА; тогда как НАДН образуется во время гликолиза .) Источник НАДФН двоякий. Когда малат окислительно декарбоксилируется «НАДФ + -связанный яблочный фермент» с образованием пирувата CO 2 , образуются и НАДФН. НАДФН также образуется по пентозофосфатному пути , который превращает глюкозу в рибозу, которая может использоваться в синтезе нуклеотидов и нуклеиновых кислот , или может катаболизироваться до пирувата. . [7]
Пируват, образующийся в результате гликолиза, является важным посредником в превращении углеводов в жирные кислоты и холестерин. [7] Это происходит путем превращения пирувата в ацетил-КоА в митохондриях. Однако этот ацетил-КоА необходимо транспортировать в цитозоль, где происходит синтез жирных кислот и холестерина. Это не может произойти напрямую. Чтобы получить цитозольный ацетил-КоА, цитрат (полученный конденсацией ацетил-КоА с оксалоацетатом) удаляется из цикла лимонной кислоты и переносится через внутреннюю митохондриальную мембрану в цитозоль. [7] Там он расщепляется АТФ-цитратлиазой на ацетил-КоА и оксалоацетат. Оксалоацетат может использоваться для глюконеогенеза (в печени) или может возвращаться в митохондрии в виде малата. [8] Цитозольный ацетил-КоА карбоксилируется ацетил-КоА-карбоксилазой с образованием малонил-КоА , что является первой стадией синтеза жирных кислот. [8] [9]
Животные не могут повторно синтезировать углеводы из жирных кислот.
Основным топливом, запасаемым в организме животных, является жир. Запасы жира у молодого взрослого человека в среднем составляют примерно 15–20 кг (33–44 фунта), но сильно варьируются в зависимости от возраста, пола и индивидуального предрасположения. [10] Напротив, в организме человека хранится всего около 400 г (0,9 фунта) гликогена , из которых 300 г (0,7 фунта) заперты внутри скелетных мышц и недоступны для организма в целом. Примерно 100 г (0,2 фунта) гликогена, хранящегося в печени, истощаются в течение одного дня голодания. [11] После этого глюкоза, которая выделяется в кровь печенью для общего использования тканями организма, должна синтезироваться из глюкогенных аминокислот и некоторых других глюконеогенных субстратов , которые не включают жирные кислоты. [12]
Жирные кислоты расщепляются до ацетил-КоА посредством бета-окисления внутри митохондрий, тогда как жирные кислоты синтезируются из ацетил-КоА вне митохондрий, в цитозоле. Эти два пути различаются не только по месту их возникновения, но также по протекающим реакциям и используемым субстратам. Эти два пути являются взаимно ингибирующими, предотвращая попадание ацетил-КоА, полученного в результате бета-окисления, в синтетический путь через реакцию ацетил-КоА-карбоксилазы . [12] Его также невозможно превратить в пируват , поскольку реакция декарбоксилирования пирувата необратима. [11] Вместо этого он конденсируется с оксалоацетатом и вступает в цикл лимонной кислоты . Во время каждого витка цикла два атома углерода покидают цикл в виде CO 2 в реакциях декарбоксилирования, катализируемых изоцитратдегидрогеназой и альфа-кетоглутаратдегидрогеназой . Таким образом, каждый виток цикла лимонной кислоты окисляет единицу ацетил-КоА, одновременно регенерируя молекулу оксалоацетата, с которой ацетил-КоА первоначально соединился с образованием лимонной кислоты . Реакции декарбоксилирования происходят до образования малата в цикле. Малат — единственное вещество, которое можно удалить из митохондрий и ввести в глюконеогенный путь с образованием глюкозы или гликогена в печени или любой другой ткани. [12] Поэтому не может быть чистого превращения жирных кислот в глюкозу.
Ацетил-КоА превращается в малонил-КоА под действием ацетил-КоА-карбоксилазы , после чего малонил-КоА предназначен для участия в пути синтеза жирных кислот. Ацетил-КоА-карбоксилаза является точкой регуляции синтеза насыщенных жирных кислот с прямой цепью и подлежит как фосфорилированию , так и аллостерическому регулированию . Регуляция посредством фосфорилирования происходит в основном у млекопитающих, тогда как аллостерическая регуляция происходит у большинства организмов. Аллостерический контроль осуществляется посредством ингибирования по принципу обратной связи пальмитоил-КоА и активации цитратом. При высоком уровне пальмитоил-КоА, конечного продукта синтеза насыщенных жирных кислот, он аллостерически инактивирует ацетил-КоА-карбоксилазу, предотвращая накопление жирных кислот в клетках. Цитрат активирует ацетил-КоА-карбоксилазу при высоких уровнях, поскольку высокие уровни указывают на то, что ацетил-КоА достаточно для участия в цикле Кребса и сохранения энергии. [13]
Высокие уровни инсулина в плазме крови (например, после еды) вызывают дефосфорилирование ацетил-КоА-карбоксилазы, способствуя тем самым образованию малонил-КоА из ацетил-КоА и, как следствие, превращению углеводов в жирные кислоты, тогда как адреналин и глюкагон (высвобождаемые в кровь во время голодания и физических упражнений) вызывают фосфорилирование этого фермента, подавляя липогенез в пользу окисления жирных кислот посредством бета-окисления . [7] [9]
Многие бактерии используют анаэробный путь синтеза ненасыщенных жирных кислот. Этот путь не использует кислород и зависит от ферментов, которые встраивают двойную связь перед элонгацией с использованием обычного механизма синтеза жирных кислот. У Escherichia coli этот путь хорошо изучен.
Синтез ненасыщенных жирных кислот путем анаэробной десатурации.
FabA представляет собой дегидразу β-гидроксидеканоил-ACP, специфичную для промежуточного продукта синтеза 10-углеродных насыщенных жирных кислот (β-гидроксидеканоил-ACP).
FabA катализирует дегидратацию β-гидроксидеканоил-ACP, вызывая высвобождение воды и внедрение двойной связи между C7 и C8, считая от метильного конца. Это создает промежуточный транс-2-деценоил.
Либо промежуточный транс-2-деценоил может быть переведен на нормальный путь синтеза насыщенных жирных кислот с помощью FabB, где двойная связь будет гидролизована и конечным продуктом будет насыщенная жирная кислота, либо FabA будет катализировать изомеризацию в цис- 3-деценоил-промежуточный продукт.
FabB представляет собой β-кетоацил-АСР-синтазу, которая удлиняет и направляет промежуточные продукты в основной путь синтеза жирных кислот. Когда FabB реагирует с промежуточным цис-деценоилом, конечным продуктом после элонгации будет ненасыщенная жирная кислота. [14]
Двумя основными ненасыщенными жирными кислотами являются пальмитолеоил-ACP (16:1ω7) и цис-вакценоил-ACP (18:1ω7). [15]
Большинство бактерий, подвергающихся анаэробной десатурации, содержат гомологи FabA и FabB. [16] Клостридии являются основным исключением; у них есть новый фермент, который еще предстоит идентифицировать, который катализирует образование двойной цис-связи. [15]
Регулирование
Этот путь подвергается транскрипционной регуляции с помощью FadR и FabR. FadR является наиболее изученным белком, которому приписаны бифункциональные характеристики. Он действует как активатор транскрипции fabA и fabB и как репрессор β-окисления регулона . Напротив, FabR действует как репрессор транскрипции fabA и fabB. [14]
Аэробная десатурация — наиболее распространенный путь синтеза ненасыщенных жирных кислот. Он используется у всех эукариот и некоторых прокариот. Этот путь использует десатуразы для синтеза ненасыщенных жирных кислот из полноразмерных субстратов насыщенных жирных кислот. [17] Все десатуразы требуют кислорода и в конечном итоге потребляют НАДН, хотя десатурация является окислительным процессом. Десатуразы специфичны в отношении двойной связи, которую они индуцируют в субстрате. У Bacillus subtilis десатураза Δ 5 -Des, специфичен для индукции двойной цис-связи в Δ 5 позиция. [8] [17] Saccharomyces cerevisiae содержит одну десатуразу, Ole1p, которая индуцирует двойную цис-связь при Δ 9 . [8]
У млекопитающих аэробная десатурация катализируется комплексом трех мембраносвязанных ферментов ( НАДН-цитохром b 5 редуктаза, цитохром b 5 и десатураза ). Эти ферменты выделяют молекулярный кислород, O 2 , взаимодействовать с цепью насыщенного жирного ацил-КоА, образуя двойную связь и две молекулы воды, H 2 О. Два электрона принадлежат НАДН + Н. + и два из одинарной связи в цепи жирной кислоты. [7] Однако эти ферменты млекопитающих не способны создавать двойные связи у атомов углерода за пределами C-9 в цепи жирных кислот. [номер 1] .) Следовательно, млекопитающие не могут синтезировать линолеат или линоленат (которые имеют двойные связи у C-12 (= Δ 12 ), или C-12 и C-15 (= Δ 12 и Δ 15 ) положения соответственно, а также на позиции ∆ 9 положение), ни полиненасыщенной 20-углеродной арахидоновой кислоты , полученной из линолеата. Все они называются незаменимыми жирными кислотами , что означает, что они необходимы организму, но могут быть получены только с пищей. (Арахидоновая кислота является предшественником простагландинов , которые выполняют широкий спектр функций местных гормонов .) [7]
У B. subtilis этот путь регулируется двухкомпонентной системой : DesK и DesR. DesK представляет собой мембраносвязанную киназу, а DesR представляет собой регулятор транскрипции гена des . [8] [17] Регулирование реагирует на температуру; при понижении температуры этот ген активируется. Ненасыщенные жирные кислоты повышают текучесть мембраны и стабилизируют ее при более низких температурах. DesK – сенсорный белок, который при понижении температуры будет аутофосфорилироваться. DesK-P перенесет свою фосфорильную группу на DesR. Два белка DesR-P будут димеризоваться и связываться с промоторами ДНК гена des и рекрутировать РНК-полимеразу для начала транскрипции. [8] [17]
синегнойная палочка
Как правило, в одной и той же системе не происходит как анаэробного, так и аэробного синтеза ненасыщенных жирных кислот, однако Pseudomonas aeruginosa и Vibrio ABE-1. исключениями являются [20] [21] [22] Хотя P. aeruginosa претерпевает главным образом анаэробную десатурацию, она также подвергается двум аэробным путям. Один путь использует Δ 9 -десатураза (DesA), катализирующая образование двойной связи в мембранных липидах. Другой путь использует два белка, DesC и DesB, которые вместе действуют как Δ 9 -десатураза, которая вставляет двойную связь в молекулу насыщенной жирной кислоты-КоА. Этот второй путь регулируется белком-репрессором DesT. DesT также является репрессором экспрессии fabAB при анаэробной десатурации в присутствии экзогенных ненасыщенных жирных кислот. Эта функция предназначена для координации экспрессии двух путей внутри организма. [21] [23]
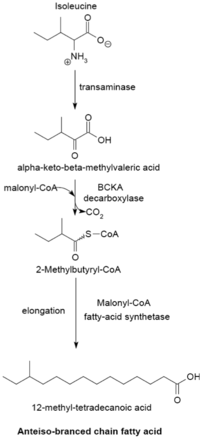
Жирные кислоты с разветвленной цепью обычно являются насыщенными и встречаются в двух различных семействах: изо-серии и антеизо-серии. Было обнаружено, что Actinomycetales содержат уникальные механизмы синтеза жирных кислот с разветвленной цепью, в том числе образующие туберкулостериновую кислоту.
Система синтеза жирных кислот с разветвленной цепью
Пути синтеза системы синтеза жирных кислот с разветвленной цепью с учетом разных праймеров
Система синтеза жирных кислот с разветвленной цепью использует α-кетокислоты в качестве праймеров. Эта система отличается от синтетазы жирных кислот с разветвленной цепью, которая использует сложные эфиры ацил-КоА с короткой цепью в качестве праймеров. [24] Праймеры α-кетокислот получаются в результате переаминирования и декарбоксилирования валина . , лейцина и изолейцина с образованием 2-метилпропанил-КоА, 3-метилбутирил-КоА и 2-метилбутирил-КоА соответственно [25] Праймеры 2-метилпропанил-КоА, полученные из валина, удлиняются для образования жирных кислот изо-ряда с четными номерами, таких как 14-метилпентадекановая (изопальмитиновая) кислота, а праймеры 3-метилбутирил-КоА из лейцина могут использоваться для образования нечетных жирные кислоты изо-ряда, такие как 13-метилтетрадекановая кислота. Праймеры 2-метилбутирил-КоА из изолейцина удлиняются с образованием жирных кислот антиизо-ряда, содержащих нечетное число атомов углерода, таких как 12-метилтетрадекановая кислота. [26] Декарбоксилирование предшественников праймера происходит с помощью фермента декарбоксилазы α-кетокислот с разветвленной цепью (BCKA). Удлинение жирной кислоты происходит по тому же пути биосинтеза в Escherichia coli, который используется для производства жирных кислот с прямой цепью, где малонил-КоА используется в качестве удлинителя цепи. [27] Основными конечными продуктами являются 12–17-углеродные жирные кислоты с разветвленной цепью, а их состав обычно однороден и характерен для многих видов бактерий. [26]
Декарбоксилаза BCKA и относительная активность субстратов α-кетокислот
Фермент декарбоксилаза BCKA состоит из двух субъединиц тетрамерной структуры (A 2 B 2 ) и необходим для синтеза жирных кислот с разветвленной цепью. Он отвечает за декарбоксилирование α-кетокислот, образующихся в результате переаминирования валина, лейцина и изолейцина, и образует праймеры, используемые для синтеза жирных кислот с разветвленной цепью. Активность этого фермента намного выше с субстратами α-кетокислот с разветвленной цепью, чем с субстратами с прямой цепью, а у видов Bacillus его специфичность наиболее высока для α-кето-β-метилвалериановой кислоты, полученной из изолейцина, за которой следует α- кетоизокапроат и α-кетоизовалерат. [26] [27] Высокое сродство фермента к α-кетокислотам с разветвленной цепью позволяет ему функционировать в качестве системы-донора праймера для синтетазы жирных кислот с разветвленной цепью. [27]
Субстрат
Деятельность БКАА
CO 2 Произведенный (нмоль/мин мг)
Км (мкМ)
Vmax (нмоль/мин мг)
L-α-кето-β-метилвалерат
100%
19.7
<1
17.8
α-кетоизовалерат
63%
12.4
<1
13.3
α-Кетоизокапроат
38%
7.4
<1
5.6
Пируват
25%
4.9
51.1
15.2
Факторы, влияющие на длину цепочки и распределение рисунка
Праймеры α-кетокислот используются для производства жирных кислот с разветвленной цепью, длина которых обычно составляет от 12 до 17 атомов углерода. Пропорции этих жирных кислот с разветвленной цепью, как правило, одинаковы и постоянны среди конкретных видов бактерий, но могут быть изменены из-за изменений концентрации малонил-КоА, температуры или присутствия термостабильных факторов (HSF). [26] Все эти факторы могут влиять на длину цепи, и было продемонстрировано, что HSF изменяют специфичность декарбоксилазы BCKA в отношении определенного субстрата α-кетокислоты, тем самым изменяя соотношение образующихся жирных кислот с разветвленной цепью. [26] Было показано, что увеличение концентрации малонил-КоА приводит к увеличению количества вырабатываемых жирных кислот C17 до тех пор, пока не будет достигнута оптимальная концентрация (≈20 мкм) малонил-КоА. Снижение температуры также имеет тенденцию слегка смещать распределение жирных кислот в сторону жирных кислот C17 у Bacillus . видов [24] [26]
Эта система функционирует аналогично системе синтеза жирных кислот с разветвленной цепью, однако в ней в качестве праймеров используются карбоновые кислоты с короткой цепью вместо альфа-кетокислот. Как правило, этот метод используется бактериями, которые не обладают способностью создавать систему жирных кислот с разветвленной цепью с использованием альфа-кето-праймеров. Типичные праймеры с короткой цепью включают изовалерат, изобутират и 2-метилбутират. Обычно кислоты, необходимые для этих праймеров, берутся из окружающей среды; это часто наблюдается у бактерий рубца. [28]
Общая реакция такая:
Изобутирил-КоА + 6 малонил-КоА + 12 НАДФН + 12 H + → Изопальмитиновая кислота + 6 CO 2 12 НАДФ + 5 Н 2 О + 7 КоА [24]
Разница между синтазой жирных кислот (с прямой цепью) и синтазой жирных кислот с разветвленной цепью заключается в субстратной специфичности фермента, который катализирует реакцию ацил-КоА в ацил-АСР. [24]
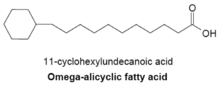
Омега-алициклические жирные кислоты обычно содержат омега-концевую пропильную или бутирильную циклическую группу и являются одними из основных мембранных жирных кислот, обнаруженных у нескольких видов бактерий. Синтетаза жирных кислот, используемая для производства омега-алициклических жирных кислот, также используется для производства мембранных жирных кислот с разветвленной цепью. У бактерий с мембранами, состоящими в основном из омега-алициклических жирных кислот, запас эфиров циклической карбоновой кислоты-КоА намного выше, чем у праймеров с разветвленной цепью. [24] Синтез циклических праймеров недостаточно изучен, но было высказано предположение, что механизм включает превращение сахаров в шикимовую кислоту , которая затем преобразуется в сложные эфиры циклогексилкарбоновой кислоты и КоА, которые служат праймерами для синтеза омега-алициклических жирных кислот. [28]
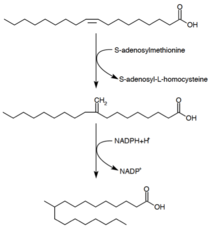
Туберкулостеариновая кислота (D-10-метилстеариновая кислота) представляет собой насыщенную жирную кислоту, которая, как известно, продуцируется Mycobacterium spp. и два вида Streptomyces . Он образуется из предшественника олеиновой кислоты (мононенасыщенной жирной кислоты). [29] После этерификации олеиновой кислоты до фосфолипида S-аденозилметионин отдает метильную группу двойной связи олеиновой кислоты. [30] Эта реакция метилирования образует промежуточный 10-метилен-октадеканоил. Последовательное восстановление остатка с НАДФН в качестве кофактора приводит к образованию 10-метилстеариновой кислоты. [25]
Помимо синтеза жирных кислот в цитозоле, митохондрии также осуществляют собственный синтез жирных кислот (mtFASII). Синтез митохондриальных жирных кислот необходим для клеточного дыхания и митохондриального биогенеза . [31] Предполагается также роль медиатора во внутриклеточной передаче сигнала , поскольку уровни биоактивных липидов, таких как лизофосфолипиды и сфинголипиды , коррелируют с mtFASII. [32]
^ Нумерация атомов углерода Положение атомов углерода в жирной кислоте может указываться по СООН- (или карбоксильному) концу или по -СН. 3 (или метильный) конец. Если указано с конца -COOH, то используются обозначения C-1, C-2, C-3,... .(и т.д.) (синие цифры на схеме справа, где C-1 — это – углерод СООН). Если позиция отсчитывается от другой, -CH 3 , в конце, то положение обозначено обозначением ω-n (цифры красного цвета, где ω-1 относится к метиловому углероду).
Таким образом, положения двойных связей в цепи жирной кислоты можно указать двумя способами: с использованием обозначений Cn или ω-n. Таким образом, в 18-углеродной жирной кислоте двойная связь между C-12 (или ω-7) и C-13 (или ω-6) обозначается либо как Δ 12 если считать от конца –COOH (что указывает только на «начало» двойной связи) или как ω-6 (или омега-6), если считать от -CH 3 конец. «Δ» — это греческая буква «дельта», которая . в латинском алфавите переводится как «D» (двойная связь) Омега (ω) — последняя буква греческого алфавита, поэтому она используется для обозначения «последнего» атома углерода в цепи жирных кислот. Поскольку обозначение ω-n используется почти исключительно для обозначения положений двойных связей, близких к -CH 3 заканчиваются незаменимыми жирными кислотами , нет необходимости в эквивалентном обозначении типа «Δ» - использование обозначения «ω-n» всегда относится к положению двойной связи.
^ Слоан, AW; Коэслаг, Дж. Х.; Бределл, ГАГ (1973). «Состав тела, работоспособность и работоспособность активных и малоподвижных юношей». Европейский журнал прикладной физиологии . 32 : 17–24. дои : 10.1007/bf00422426 . S2CID 39812342 .
^ Jump up to: а б Страйер, Люберт (1995). Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. стр. 581–602, 613, 775–778. ISBN 0-7167-2009-4 .
^ Jump up to: а б с д Страйер, Люберт (1995). «Обмен жирных кислот». Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. стр. 603–628. ISBN 0-7167-2009-4 .
^ Jump up to: а б с д и ж Наик Д.Н., Канеда Т. (декабрь 1974 г.). «Биосинтез разветвленных длинноцепочечных жирных кислот видами Bacillus: относительная активность трех субстратов альфа-кетокислот и факторов, влияющих на длину цепи». Может J Микробиол . 20 (12): 1701–8. дои : 10.1139/m74-263 . ПМИД 4155346 .
^ Рэтледж, Колин; Стэнфорд, Джон (1982). Физиология, идентификация и классификация . Биология микобактерий. Академический. ISBN 9780125823012 . OCLC 248050385 .
^ Кубица, Джордж П.; Уэйн, Лоуренс Г. (1984). Микобактерии: справочник . Деккер. ISBN 9780824719173 .
Arc.Ask3.Ru Номер скриншота №: 6defd986150fa7ffda306e17c6063be0__1721643540 URL1:https://arc.ask3.ru/arc/aa/6d/e0/6defd986150fa7ffda306e17c6063be0.html Заголовок, (Title) документа по адресу, URL1: Fatty acid synthesis - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)