Ацетил-КоА-карбоксилаза
| ацетил-КоА-карбоксилаза | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Номер ЕС. | 6.4.1.2 | ||
| Номер CAS. | 9023-93-2 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| Ацетил-КоА-карбоксилаза альфа | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | акация | ||
| Альт. символы | АСАС, АСС1, АССА | ||
| ген NCBI | 31 | ||
| HGNC | 84 | ||
| МОЙ БОГ | 601557 | ||
| RefSeq | НМ_198839 | ||
| ЮниПрот | Q13085 | ||
| Другие данные | |||
| Номер ЕС | 6.4.1.2 | ||
| Локус | Хр. 17 q21 | ||
| |||
| Ацетил-КоА-карбоксилаза бета | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | АКАСВ | ||
| Альт. символы | АСС2, АККБ | ||
| ген NCBI | 32 | ||
| HGNC | 85 | ||
| МОЙ БОГ | 200350 | ||
| RefSeq | НМ_001093 | ||
| ЮниПрот | О00763 | ||
| Другие данные | |||
| Номер ЕС | 6.4.1.2 | ||
| Локус | Хр. 12 q24.1 | ||
| |||
Ацетил-КоА-карбоксилаза ( АСС ) представляет собой биотин -зависимый фермент ( EC 6.4.1.2 ), который катализирует необратимое карбоксилирование ацетил -КоА с образованием малонил-КоА посредством двух его каталитических активностей: биотинкарбоксилазы (BC) и карбоксилтрансферазы (CT). АСС представляет собой многосубъединичный фермент у большинства прокариот и в хлоропластах большинства растений и водорослей, тогда как это крупный многодоменный фермент в цитоплазме большинства эукариот . Наиболее важной функцией АСС является обеспечение субстратом малонил-КоА для биосинтеза жирных кислот . [1] Активность АСС можно контролировать на уровне транскрипции, а также с помощью модуляторов малых молекул и ковалентной модификации . Геном человека содержит гены двух разных АСС. [2] - ТАМ [3] и АКАСБ . [4]
Структура
[ редактировать ]Прокариоты и растения имеют многосубъединичные АСС, состоящие из нескольких полипептидов. Активность биотинкарбоксилазы (BC), белка-переносчика биотинкарбоксила (BCCP) и активность карбоксилтрансферазы (CT) содержатся в разных субъединицах. Стехиометрия этих субъединиц холофермента АСС различается у разных организмов. [1] У людей и большинства эукариот развился ACC с каталитическими доменами CT и BC и доменами BCCP на одном полипептиде. Большинство растений также имеют эту гомомерную форму в цитозоле. [5] Функциональными областями АСС, начиная с N-конца до С-конца, являются биотинкарбоксилаза (BC), биотинсвязывающая (BB), карбоксилтрансфераза (CT) и АТФ-связывающая (AB). АВ лежит внутри ВС. Биотин ковалентно присоединен через амидную связь к длинной боковой цепи лизина, находящегося в BB. Поскольку BB находится между областями BC и CT, биотин может легко перемещаться в оба активных центра, где он необходим.
У млекопитающих, у которых экспрессируются две изоформы АСС, основным структурным различием между этими изоформами является расширенный N-конец АСС2, содержащий последовательность, нацеленную на митохондрии . [1]
 Субъединица биотинкарбоксилазы E. coli ацетил-КоА-карбоксилазы
Субъединица биотинкарбоксилазы E. coli ацетил-КоА-карбоксилазы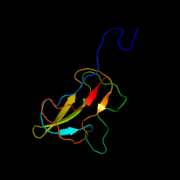 Субъединица белка-переносчика биотина E. coli ацетил-КоА-карбоксилазы
Субъединица белка-переносчика биотина E. coli ацетил-КоА-карбоксилазы Карбоксилтрансферазная субъединица E. coli ацетил-КоА-карбоксилазы
Карбоксилтрансферазная субъединица E. coli ацетил-КоА-карбоксилазы



Гены
[ редактировать ]Полипептиды, составляющие многосубъединичные АСС прокариот и растений, кодируются разными генами. В Escherichia coli AccA кодирует альфа-субъединицу ацетил-КоА-карбоксилазы. [6] и accD кодирует свою бета-субъединицу. [7]
Механизм
[ редактировать ]Общая реакция ACAC(A,B) протекает по двухстадийному механизму. [8] Первая реакция осуществляется БЦ и включает АТФ-зависимое карбоксилирование биотина бикарбонатом , служащим источником CO 2 . Карбоксильная группа переносится от биотина к ацетил-КоА с образованием малонил-КоА во второй реакции, катализируемой ХТ.
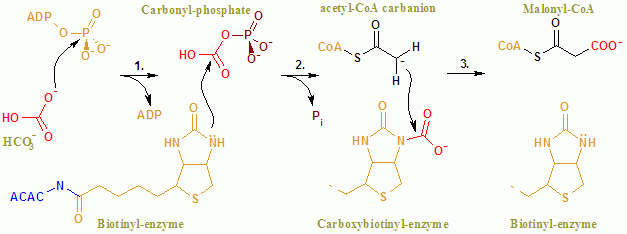
В активном центре реакция протекает при активном взаимодействии остатков Glu296 и положительно заряженных Arg338 и Arg292 с субстратами. [9] Два мг 2+ координируются фосфатными группами АТФ и необходимы для связывания АТФ с ферментом. Бикарбонат депротонируется Glu296, хотя в растворе этот перенос протона маловероятен, поскольку pKa бикарбоната составляет 10,3. Фермент, по-видимому, манипулирует рКа, облегчая депротонирование бикарбоната. рКа бикарбоната снижается за счет его взаимодействия с положительно заряженными боковыми цепями Arg338 и Arg292. Кроме того, Glu296 взаимодействует с боковой цепью Glu211, и было показано, что это взаимодействие вызывает увеличение кажущейся pKa. После депротонирования бикарбоната кислород бикарбоната действует как нуклеофил и атакует гамма-фосфат на АТФ. Промежуточный карбоксифосфат быстро разлагается на CO 2 и PO 4. 3− . ПО 4 3− депротонирует биотин, создавая енолят, стабилизированный Arg338, который впоследствии атакует CO2, что приводит к образованию карбоксибиотина. [9] Карбоксибиотин транслоцируется в активный центр карбоксилтрансферазы (CT), где карбоксильная группа переносится на ацетил-КоА. В отличие от домена BC, о механизме реакции CT мало что известно. Предполагаемый механизм — высвобождение CO 2 из биотина, который впоследствии отрывает протон от метильной группы ацетил-КоА-карбоксилазы. Образующийся енолят атакует CO 2 с образованием малонил-КоА. В конкурирующем механизме отрыв протона сочетается с атакой ацетил-КоА.
Функция
[ редактировать ]Функция АСС заключается в регулировании метаболизма жирных кислот. Когда фермент активен, вырабатывается продукт малонил-КоА, который является строительным блоком для новых жирных кислот и может ингибировать перенос жирной ацильной группы от ацил-КоА к карнитину с помощью карнитинацилтрансферазы , которая ингибирует бета-окисление. жирных кислот в митохондриях .
У млекопитающих две основные изоформы экспрессируются АСС, АСС1 и АСС2, которые различаются как по распределению в тканях, так и по функциям. АСС1 обнаруживается в цитоплазме всех клеток, но он обогащен липогенной тканью, такой как жировая ткань и лактирующие молочные железы , где важен синтез жирных кислот. [10] В окислительных тканях, таких как скелетные мышцы и сердце , соотношение экспрессируемого АСС2 выше. ACC1 и ACC2 высоко экспрессируются в печени , где важны как окисление, так и синтез жирных кислот. [11] Различия в распределении тканей указывают на то, что АСС1 поддерживает регуляцию синтеза жирных кислот , тогда как АСС2 главным образом регулирует окисление жирных кислот (бета-окисление).
Митохондриальная изоформа АСС1 (mACC1) играет частично дублирующую роль в синтезе липоевой кислоты и, следовательно, в липоилировании белков, обеспечивая малонил-КоА для синтеза митохондриальных жирных кислот (mtFASII) в тандеме с ACSF3 . [12] [13]
Регулирование
[ редактировать ]
Регуляция АСС у млекопитающих является сложной: она контролирует два различных пула малонил-КоА, которые направляют либо ингибирование бета-окисления, либо активацию биосинтеза липидов. [14]
ACC1 и ACC2 млекопитающих регулируются транскрипционно с помощью множества промоторов , которые опосредуют численность ACC в ответ на статус питания клеток. Активация экспрессии генов посредством различных промоторов приводит к альтернативному сплайсингу ; однако физиологическое значение специфических изоферментов АСС остается неясным. [11] Чувствительность к статусу питания является результатом контроля этих промоторов с помощью факторов транскрипции, таких как белок, связывающий регуляторные элементы стерола 1 , контролируемый инсулином на уровне транскрипции, и ChREBP , экспрессия которого увеличивается при диете с высоким содержанием углеводов . [15] [16]
Через петлю прямой связи цитрат аллостерически активирует АСС. [17] Цитрат может усиливать полимеризацию АСС , повышая ферментативную активность; однако неясно, является ли полимеризация основным механизмом цитрата повышения активности АСС или полимеризация является артефактом экспериментов in vitro. Другие аллостерические активаторы включают глутамат и другие дикарбоновые кислоты . [18] Длинно- и короткоцепочечные жирные ацил-КоА являются с отрицательной обратной связью . ингибиторами АСС [19] Одним из таких отрицательных аллостерических модуляторов является пальмитоил-КоА. [20]
Фосфорилирование может произойти, когда гормон глюкагон [21] или адреналин [22] связываются с рецепторами клеточной поверхности , но основная причина фосфорилирования связана с повышением уровня AMP при низком энергетическом статусе клетки, что приводит к активации AMP-активируемой протеинкиназы (AMPK). AMPK является основным регулятором киназы АСС, способным фосфорилировать ряд остатков серина на обеих изоформах АСС. [23] На ACC1 AMPK фосфорилирует Ser79, Ser1200 и Ser1215. Протеинкиназа А также обладает способностью фосфорилировать АСС, причем способность фосфорилировать АСС2 гораздо выше, чем АСС1. Ser80 и Ser1263 на ACC1 также могут служить местом фосфорилирования в качестве регуляторного механизма. [24] Однако физиологическое значение протеинкиназы А в регуляции АСС в настоящее время неизвестно. Исследователи предполагают, что существуют и другие киназы АСС, важные для его регуляции, поскольку существует множество других возможных сайтов фосфорилирования АСС. [25]
Когда инсулин связывается со своими рецепторами на клеточной мембране , он активирует фермент фосфатазу, называемый протеинфосфатазой 2А (PP2A), для дефосфорилирования фермента; тем самым устраняя ингибирующий эффект. Кроме того, инсулин индуцирует фосфодиэстеразу, которая снижает уровень цАМФ в клетке, ингибируя тем самым PKA, а также напрямую ингибирует AMPK. [ нужна ссылка ]
Этот белок может использовать морфеиновую модель аллостерической регуляции . [26]
Клинические последствия
[ редактировать ]На стыке путей синтеза и окисления липидов АКК открывает множество клинических возможностей для производства новых антибиотиков и разработки новых методов лечения диабета , ожирения и других проявлений метаболического синдрома . [27] Исследователи стремятся воспользоваться структурными различиями между бактериальными и человеческими АКК для создания антибиотиков, специфичных для бактериальных АКК, чтобы минимизировать побочные эффекты для пациентов. Многообещающие результаты о полезности ингибитора АСС включают в себя обнаружение того, что мыши без экспрессии АСС2 имеют постоянное окисление жирных кислот, снижение массы тела и массы тела, несмотря на увеличение потребления пищи. Эти мыши также защищены от диабета. [14] Отсутствие ACC1 у мутантных мышей смертельно опасно уже на эмбриональной стадии. Однако неизвестно, должны ли препараты, нацеленные на АСС у человека, быть специфичными для АСС2. [28]
Фирсокостат (ранее GS-976, ND-630, NDI-010976) является мощным аллостерическим ингибитором АСС, действующим в домене BC АСС. [29] Фирсокостат находится в стадии разработки в 2019 году (Фаза II). [30] фармацевтической компанией Gilead в рамках комбинированного лечения неалкогольного стеатогепатита (НАСГ), который считается растущей причиной печеночной недостаточности. [31]
Кроме того, селективные ингибиторы АСС широко используются в качестве гербицидов . [32] что предполагает клиническое применение против паразитов Apicomplexa , которые основаны на изоформе ACC растительного происхождения, [33] включая малярию .
Считается, что гетерогенные клинические фенотипы метаболического заболевания, сочетающего малоновую и метилмалоновую ацидурию (CMAMMA), обусловленные дефицитом ACSF3, являются результатом частичной компенсации митохондриальной изоформы ACC1 (mACC1) дефицита ACSF3 в синтезе митохондриальных жирных кислот (mtFASII). [34]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Тонг Л. (август 2005 г.). «Ацетил-коэнзим А-карбоксилаза: важнейший метаболический фермент и привлекательная мишень для открытия лекарств» . Клеточные и молекулярные науки о жизни . 62 (16): 1784–1803. дои : 10.1007/s00018-005-5121-4 . ПМЦ 11139103 . ПМИД 15968460 . S2CID 1131957 .
- ^ Браунси Р.В., Жанде Р., Бун А.Н. (ноябрь 1997 г.). «Изоформы ацетил-КоА-карбоксилазы: структура, регуляторные свойства и метаболические функции». Труды Биохимического общества . 25 (4): 1232–1238. дои : 10.1042/bst0251232 . ПМИД 9449982 .
- ^ Абу-Эльхейга Л., Джаякумар А., Бальдини А., Чирала С.С., Вакил С.Дж. (апрель 1995 г.). «Человеческая ацетил-КоА-карбоксилаза: характеристика, молекулярное клонирование и доказательства существования двух изоформ» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (9): 4011–4015. Бибкод : 1995PNAS...92.4011A . дои : 10.1073/pnas.92.9.4011 . ПМК 42092 . ПМИД 7732023 .
- ^ Видмер Дж., Фассихи К.С., Шлихтер С.С., Уиллер К.С., Крут Б.Е., Кинг Н. и др. (июнь 1996 г.). «Идентификация второго гена ацетил-КоА-карбоксилазы человека» . Биохимический журнал . 316 (Часть 3): 915–922. дои : 10.1042/bj3160915 . ПМЦ 1217437 . ПМИД 8670171 .
- ^ Сасаки Ю, Нагано Ю (июнь 2004 г.). «Растительная ацетил-КоА-карбоксилаза: структура, биосинтез, регуляция и манипуляции с генами для селекции растений» . Бионауки, биотехнологии и биохимия . 68 (6): 1175–1184. дои : 10.1271/bbb.68.1175 . ПМИД 15215578 . S2CID 41506311 .
- ^ «accA, альфа-субъединица ацетил-КоА-карбоксилазы ( субстр. MG1655 Escherichia coli str. K-12)» . Ген NCBI . Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ «accD, бета-субъединица ацетил-КоА-карбоксилазы ( Escherichia coli субстр. K-12, MG1655)» . Ген NCBI . Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ Ли С.К., Чеонг Х.К., Рю К.С., Ли Джи, Ли В., Чон Ю.Х. и др. (август 2008 г.). «Биотиноильный домен ацетил-КоА-карбоксилазы человека: структурное понимание механизма переноса карбоксила». Белки . 72 (2): 613–624. дои : 10.1002/прот.21952 . ПМИД 18247344 . S2CID 24548083 .
- ^ Jump up to: а б Чоу С.И., Ю Л.П., Тонг Л. (апрель 2009 г.). «Кристаллическая структура биотинкарбоксилазы в комплексе с субстратами и значение ее каталитического механизма» . Журнал биологической химии . 284 (17): 11690–11697. дои : 10.1074/jbc.M805783200 . ПМК 2670172 . ПМИД 19213731 .
- ^ Ким Т.С., Лихи П., Фрик ХК (август 1996 г.). «Использование промотора определяет тканеспецифическую чувствительность гена ацетил-КоА-карбоксилазы крысы». Связь с биохимическими и биофизическими исследованиями . 225 (2): 647–653. дои : 10.1006/bbrc.1996.1224 . ПМИД 8753813 .
- ^ Jump up to: а б Барбер MC, Прайс NT, Трэверс MT (март 2005 г.). «Структура и регуляция генов ацетил-КоА-карбоксилазы многоклеточных животных». Biochimica et Biophysical Acta (BBA) - Молекулярная и клеточная биология липидов . 1733 (1): 1–28. дои : 10.1016/j.bbalip.2004.12.001 . ПМИД 15749055 .
- ^ Монтеууис Г., Суоми Ф., Кератар Дж.М., Масуд А.Дж., Кастаниотис А.Дж. (ноябрь 2017 г.). «Консервативная митохондриальная изоформа ацетил-КоА-карбоксилазы ACC1 млекопитающих обеспечивает малонил-КоА, необходимый для биогенеза митохондрий, в тандеме с ACSF3» . Биохимический журнал . 474 (22): 3783–3797. дои : 10.1042/BCJ20170416 . ПМИД 28986507 .
- ^ Кастаниотис А.Дж., Аутио К.Дж., Р.Наир Р. (апрель 2021 г.). «Митохондриальные жирные кислоты и нейродегенеративные заболевания» . Нейробиолог . 27 (2): 143–158. дои : 10.1177/1073858420936162 . ISSN 1073-8584 . ПМИД 32644907 . S2CID 220472402 .
- ^ Jump up to: а б Абу-Эльхейга Л., Мацук М.М., Або-Хашема К.А., Вакил С.Дж. (март 2001 г.). «Непрерывное окисление жирных кислот и снижение накопления жира у мышей, лишенных ацетил-КоА-карбоксилазы 2». Наука . 291 (5513): 2613–2616. Бибкод : 2001Sci...291.2613A . дои : 10.1126/science.1056843 . ПМИД 11283375 . S2CID 748630 .
- ^ Филд Ф.Дж., Борн Э., Мурти С., Матур С.Н. (декабрь 2002 г.). «Полиненасыщенные жирные кислоты снижают экспрессию белка-1, связывающего регуляторный элемент стерола, в клетках CaCo-2: влияние на синтез жирных кислот и транспорт триацилглицеринов» . Биохимический журнал . 368 (Часть 3): 855–864. дои : 10.1042/BJ20020731 . ПМК 1223029 . ПМИД 12213084 .
- ^ Исии С., Иизука К., Миллер Б.С., Уеда К. (ноябрь 2004 г.). «Белок, связывающий элемент ответа на углеводы, непосредственно способствует транскрипции гена липогенного фермента» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (44): 15597–15602. Бибкод : 2004PNAS..10115597I . дои : 10.1073/pnas.0405238101 . ПМК 524841 . ПМИД 15496471 .
- ^ Мартин Д.Б., Vagelos PR (июнь 1962 г.). «Механизм регуляции цикла трикарбоновых кислот синтеза жирных кислот» . Журнал биологической химии . 237 (6): 1787–1792. дои : 10.1016/S0021-9258(19)73938-6 . ПМИД 14470343 .
- ^ Бун А.Н., Чан А., Кулпа Дж.Э., Браунси Р.В. (апрель 2000 г.). «Бимодальная активация ацетил-КоА-карбоксилазы глутаматом» . Журнал биологической химии . 275 (15): 10819–10825. дои : 10.1074/jbc.275.15.10819 . ПМИД 10753875 .
- ^ Фагерман, штат Нью-Джерси, Кнудсен Дж. (апрель 1997 г.). «Роль длинноцепочечных жирных эфиров ацил-КоА в регуляции метаболизма и передаче сигналов в клетках» . Биохимический журнал . 323 (Часть 1): 1–12. дои : 10.1042/bj3230001 . ПМЦ 1218279 . ПМИД 9173866 .
- ^ Фам Т., Уолден Э., Хуард С., Пезаки Дж., Фуллертон М.Д., Баец К. (июль 2022 г.). «Точная настройка активности ацетил-КоА-карбоксилазы 1 посредством локализации: функциональная геномика раскрывает роль лизин-ацетилтрансферазы NuA4 и метаболизма сфинголипидов в регулировании активности и локализации Acc1» . Генетика . 221 (4). doi : 10.1093/genetics/iyac086 . ПМЦ 9339284 . ПМИД 35608294 .
- ^ Пэн И.С., Чен З., Сюй П.Х., Су Ми, Цай М.Д., Шай Цзюй (апрель 2010 г.). «Глюкагон активирует AMP-активируемый путь протеинкиназы/ацетил-КоА-карбоксилазы в адипоцитах» . Журнал ФАСЭБ . 24 (С1). дои : 10.1096/fasebj.24.1_supplement.995.4 . ISSN 0892-6638 . S2CID 81688050 .
- ^ Ко Х.Дж., Хиршман М.Ф., Хе Х., Ли Ю., Манабе Ю., Бальски Дж.А. и др. (май 2007 г.). «Адреналин является важнейшим медиатором острой активации АМФ-активируемой протеинкиназы в адипоцитах, вызванной физической нагрузкой» . Биохимический журнал . 403 (3): 473–481. дои : 10.1042/BJ20061479 . ПМЦ 1876380 . ПМИД 17253964 .
- ^ Пак С.Х., Гаммон С.Р., Книпперс Дж.Д., Полсен С.Р., Рубинк Д.С., Уиндер В.В. (июнь 2002 г.). «Взаимосвязь активности фосфорилирования AMPK и ацетил-КоА-карбоксилазы в мышцах». Журнал прикладной физиологии . 92 (6): 2475–2482. doi : 10.1152/japplphysicalol.00071.2002 . ПМИД 12015362 .
- ^ Вэй Дж., Тонг Л. (сентябрь 2018 г.). «Как полимеризация регулирует ацетил-КоА-карбоксилазу человека 1?» . Биохимия . 57 (38): 5495–5496. doi : 10.1021/acs.biochem.8b00881 . ПМИД 30211541 . S2CID 52193976 .
- ^ Браунси Р.В., Бун А.Н., Эллиотт Дж.Э., Кулпа Дж.Э., Ли В.М. (апрель 2006 г.). «Регуляция ацетил-КоА-карбоксилазы». Труды Биохимического общества . 34 (Часть 2): 223–227. дои : 10.1042/BST20060223 . ПМИД 16545081 .
- ^ Селвуд Т., Яффе ЭК (март 2012 г.). «Динамическая диссоциация гомоолигомеров и контроль функции белка» . Архив биохимии и биофизики . 519 (2): 131–143. дои : 10.1016/j.abb.2011.11.020 . ПМЦ 3298769 . ПМИД 22182754 .
- ^ Корбетт Дж.В., Харвуд Дж.Х. (ноябрь 2007 г.). «Ингибиторы ацетил-КоА-карбоксилазы млекопитающих». Недавние патенты на открытие сердечно-сосудистых препаратов . 2 (3): 162–180. дои : 10.2174/157489007782418928 . ПМИД 18221116 .
- ^ Абу-Эльхейга Л., Мацук М.М., Кордари П., Ох В., Шайкенов Т., Гу З. и др. (август 2005 г.). «Мутантные мыши, у которых отсутствует ацетил-КоА-карбоксилаза 1, эмбрионально летальны» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (34): 12011–12016. Бибкод : 2005PNAS..10212011A . дои : 10.1073/pnas.0505714102 . ПМК 1189351 . ПМИД 16103361 .
- ^ Гарриман Дж., Гринвуд Дж., Бхат С., Хуанг Х., Ван Р., Пол Д. и др. (март 2016 г.). «Ингибирование ацетил-КоА-карбоксилазы с помощью ND-630 снижает стеатоз печени, улучшает чувствительность к инсулину и модулирует дислипидемию у крыс» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (13): E1796–E1805. Бибкод : 2016PNAS..113E1796H . дои : 10.1073/pnas.1520686113 . ПМЦ 4822632 . ПМИД 26976583 .
- ^ Тонг А (11 апреля 2019 г.). «Gilead укрепляет надежду на коктейль NASH, увидев положительные данные, подтверждающие концепцию» . Новости конечных точек .
- ^ Лукас С., Лукас Г., Лукас Н., Кржовска-Фирич Дж., Томасевич К. (сентябрь 2018 г.). «Систематический обзор настоящего и будущего неалкогольной жировой болезни печени» . Клиническая и экспериментальная гепатология . 4 (3): 165–174. дои : 10.5114/ceh.2018.78120 . ПМК 6185929 . ПМИД 30324141 .
- ^ Аль-Хатиб К. «Ингибиторы ацетил-КоА-карбоксилазы (АССазы)» . Симптомы гербицидов . Отдел сельского хозяйства и природных ресурсов Калифорнийского университета в Дэвисе.
- ^ Зутер Э., Джонсон Дж. Дж., Хазелькорн Р., Маклеод Р., Горницки П. (ноябрь 1999 г.). «Рост Toxoplasma gondii ингибируется арилоксифеноксипропионатными гербицидами, нацеленными на ацетил-КоА-карбоксилазу» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (23): 13387–13392. Бибкод : 1999PNAS...9613387Z . дои : 10.1073/pnas.96.23.13387 . ПМК 23957 . ПМИД 10557330 .
- ^ Туччи С (январь 2020 г.). «Мозговой метаболизм и неврологические симптомы при комбинированной малоновой и метилмалоновой ацидурии» . Сиротский журнал редких заболеваний . 15 (1): 27. дои : 10.1186/s13023-020-1299-7 . ПМК 6977288 . ПМИД 31969167 .
Дальнейшее чтение
[ редактировать ]- Воет Д., Воет Дж.Г. (2004). Биохимия (3-е изд.). Уайли. ISBN 978-0-471-19350-0 .
- Бьюкенен Б.Б., Груиссем В., Джонс Р.Л., ред. (2000). Биохимия и молекулярная биология растений . Американское общество физиологов растений. ISBN 978-0-943088-37-2 .
- Леверт К.Л., Уолдроп Г.Л., Стивенс Дж.М. (май 2002 г.). «Аналог биотина ингибирует активность ацетил-КоА-карбоксилазы и адипогенез» . Журнал биологической химии . 277 (19): 16347–16350. дои : 10.1074/jbc.C200113200 . ПМИД 11907024 .