AMP-активируемая протеинкиназа
| AMP-активируемая протеинкиназа (AMPK) | |||
|---|---|---|---|
 AMP-активируемая протеинкиназа | |||
| Идентификаторы | |||
| Номер ЕС. | 2.7.11.1 | ||
| Номер CAS. | 172522-01-9 | ||
| Альт. имена | AMP-активируемая протеинкиназа | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
5'-АМФ-активируемая протеинкиназа или AMPK или 5'-аденозинмонофосфат-активируемая протеинкиназа представляет собой фермент (EC 2.7.11.31), который играет роль в гомеостазе клеточной энергии, в основном активируя поглощение и окисление глюкозы и жирных кислот, когда клеточная энергия расходуется. низкий. Он принадлежит к высококонсервативному семейству эукариотических белков, его ортологами являются SNF1 у дрожжей и SnRK1 у растений. Он состоит из трех белков ( субъединиц ), которые вместе образуют функциональный фермент, сохраняющийся от дрожжей до человека. Он экспрессируется в ряде тканей, включая печень , мозг и скелетные мышцы . В ответ на связывание AMP и ADP , [1] Конечным эффектом активации AMPK является стимуляция в печени окисления жирных кислот , кетогенеза , стимуляция окисления жирных кислот скелетных мышц и поглощения глюкозы, ингибирование синтеза холестерина , липогенеза и синтеза триглицеридов , ингибирование липогенеза адипоцитов, ингибирование липолиза адипоцитов и модуляция. секреции инсулина поджелудочной железы β-клетками . [2]
Его не следует путать с циклической АМФ -активируемой протеинкиназой ( протеинкиназа А ). [3]
Структура
[ редактировать ]AMPK представляет собой гетеротримерный белковый комплекс , образованный субъединицами α, β и γ. Каждая из этих трех субъединиц играет определенную роль как в стабильности, так и в активности AMPK. [4] [5] В частности, субъединица γ включает четыре конкретных домена цистатионин-β-синтазы (CBS) , что придает AMPK способность чувствительно обнаруживать изменения в соотношении AMP / ATP . AMPK деактивируется при замещении AMP АТФ в сайте CBS 3, что позволяет предположить, что CBS3 является первичным аллостерическим регуляторным сайтом. [6] [7] [8] Четыре домена CBS создают два сайта связывания для AMP, обычно называемые доменами Бейтмана. Связывание одного AMP с доменом Бейтмана кооперативно увеличивает аффинность связывания второго AMP с другим доменом Бейтмана. [9] [ не удалось пройти проверку ] Поскольку AMP связывает оба домена Бейтмана, субъединица γ претерпевает конформационные изменения, в результате которых обнажается каталитический домен, обнаруженный в субъединице α. Именно в этом каталитическом домене AMPK активируется, когда фосфорилирование происходит по треонину -172 (на изоформе α1) или Thr-174 (на изоформе α2) с помощью вышестоящей AMPK киназы ( AMPKK ). [10] [6] Субъединицы α, β и γ также могут находиться в разных изоформах: субъединица γ может существовать как изоформа γ1, γ2 или γ3 ; субъединица β может существовать как изоформа β1 или β2; и субъединица α может существовать как изоформа α1 или α2. Хотя наиболее распространенными изоформами, экспрессируемыми в большинстве клеток, являются изоформы α1, β1 и γ1, было продемонстрировано, что изоформы α2, β2, γ2 и γ3 также экспрессируются в сердечных и скелетных мышцах . [4] [11] [12]
Следующие гены человека кодируют субъединицы AMPK:
Кристаллическая структура регуляторного основного домена AMPK млекопитающих (α C-конец, β C-конец, γ) была решена в комплексе с AMP. [13] АДП [14] или АТФ. [15]
Регулирование
[ редактировать ]Благодаря наличию изоформ ее компонентов у млекопитающих существует 12 вариантов AMPK, каждый из которых может иметь разную тканевую локализацию и разные функции в разных условиях. [16] AMPK регулируется аллостерически и посттрансляционной модификацией, которые работают вместе. [16]
Если остаток Thr-172 α1-субъединицы AMPK (или Thr-174 α2-субъединицы AMPK) фосфорилируется, AMPK активируется примерно в 100 раз; [6] доступ фосфатаз к этому остатку блокируется, если AMP или ADP могут блокировать доступ, а АТФ может вытеснять AMP и ADP. [16] Этот остаток фосфорилируется как минимум тремя киназами ( киназой печени B1 (LKB1), [17] который работает в комплексе с STRAD и MO25 , кальций/кальмодулин-зависимой протеинкиназной киназой II-( CAMKK2 ) и TGFβ-активируемой киназой 1 (TAK1)) и дефосфорилируется тремя фосфатазами ( протеинфосфатазой 2А (PP2A); протеинфосфатазой 2C (PP2C) и Mg 2+ -/Мн 2+ -зависимая протеинфосфатаза 1Е ( PPM1E )). [16]
Регуляция AMPK с помощью CaMKK2 требует прямого взаимодействия этих двух белков через их киназные домены. Во взаимодействии CaMKK2 с AMPK участвуют только субъединицы α и β AMPK (AMPKγ отсутствует в комплексе CaMKK2), что приводит к регуляции AMPK в этом контексте за счет изменений уровней кальция, но не за счет AMP или ADP.
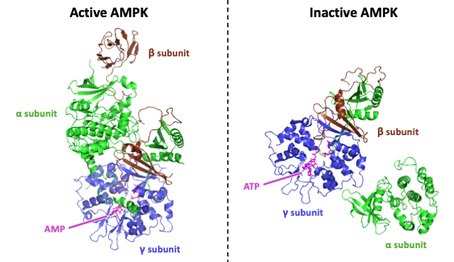
AMPK регулируется аллостерически, главным образом, путем конкурентного связывания с сайтами CBS на его γ-субъединице между АТФ (который обеспечивает доступ фосфатазы к Thr-172) и AMP или ADP (каждый из которых блокирует доступ фосфатазы). [1] Таким образом, оказывается, что AMPK является датчиком соотношения AMP/ATP или ADP/ATP и, следовательно, уровня энергии клетки. [16] AMPK претерпевает большие конформационные изменения при связывании АТФ. Область α-субъединицы, известная как киназный домен (KD), отделяется от своей конформации активного состояния и слабо ассоциируется с γ-субъединицей на расстоянии ~ 100 Å. KD также поворачивается на ~ 180 ° при конформационном изменении. При диссоциации KD активная петля (AL) α-субъединицы, которая содержит критический фосфорилированный остаток Thr, полностью подвергается воздействию вышестоящих фосфатаз. Это конформационное изменение представляет собой вероятный механизм модуляции AMPK. Когда клеточное энергетическое состояние низкое (высокие уровни AMP/ATP или ADP/ATP), AMPK принимает конформацию, связанную с KD, и AMPK защищена от дефосфорилирования и остается активированной. Когда клеточное энергетическое состояние высокое, AMPK принимает конформацию со смещением KD, AL подвергается воздействию вышестоящих фосфатаз, и AMPK деактивируется. [6]
Фармакологические соединения Merck Compound 991 и Abbott A769662 связываются с аллостерическим сайтом лекарственного средства и метаболизма (ADaM) на β-субъединице и, как было показано, активируют AMPK до 10 раз. [6] [18] Связывание сайта AdaM может играть роль в активации AMPK, а также в защите от дефосфорилирования. [19]
Существуют и другие механизмы, с помощью которых AMPK ингибируется или активируется инсулином, лептином и диацилглицерином , индуцируя различные другие фосфорилирования. [16] [а]
AMPK может ингибироваться или активироваться различными тканеспецифичными убиквитинациями . [16]
Он также регулируется несколькими белок-белковыми взаимодействиями и может либо активироваться, либо ингибироваться окислительными факторами; По состоянию на 2016 год роль окисления в регуляции AMPK была спорной. [16]
Функция
[ редактировать ]Когда AMPK фосфорилирует ацетил-КоА-карбоксилазу 1 (ACC1) или белок, связывающий регуляторные элементы стерола 1c (SREBP1c), он ингибирует синтез жирных кислот, холестерина и триглицеридов, а также активирует поглощение жирных кислот и β-окисление. [16]
AMPK стимулирует поглощение глюкозы в скелетных мышцах путем фосфорилирования Rab-GTPase-активирующего белка TBC1D1 , что в конечном итоге индуцирует слияние везикул GLUT1 с плазматической мембраной. [16] AMPK стимулирует гликолиз путем активации фосфорилирования 6-фосфофрукто-2-киназы/фруктозо-2,6-бисфосфатазы 2/3 и активации фосфорилирования гликогенфосфорилазы, а также ингибирует синтез гликогена за счет ингибирующего фосфорилирования гликогенсинтазы. [16] В печени AMPK ингибирует глюконеогенез путем ингибирования факторов транскрипции, включая ядерный фактор гепатоцитов 4 (HNF4) и коактиватор транскрипции 2, регулируемый CREB (CRTC2). [16]
AMPK ингибирует энергоемкий процесс биосинтеза белка , а также может вызвать переключение с кэп-зависимой трансляции на кэп-независимую трансляцию, которая требует меньше энергии, путем фосфорилирования TSC2 , RPTOR , фактора инициации транскрипции 1A.66 и eEF2K . [16] Когда TSC2 активирован, он ингибирует mTORC1. В результате ингибирования mTORC1 AMPK синтез белка останавливается. Активация AMPK означает низкий уровень энергии внутри клетки, поэтому все пути потребления энергии, такие как синтез белка, ингибируются, а пути, генерирующие энергию, активируются для восстановления соответствующих уровней энергии в клетке. [20]
AMPK активирует аутофагию , прямо и косвенно активируя ULK1 . [16] AMPK также, по-видимому, стимулирует биогенез митохондрий, регулируя PGC-1α , что, в свою очередь, способствует транскрипции генов в митохондриях. [16] AMPK также активирует антиоксидантную защиту. [16]
Клиническое значение
[ редактировать ]Упражнения/тренировки
[ редактировать ]Многие биохимические адаптации скелетных мышц, которые происходят во время одного упражнения или продолжительной тренировки , такие как увеличение митохондрий , биогенеза и емкости [21] [22] увеличение мышечного гликогена , [23] и увеличение количества ферментов , которые специализируются на поглощении глюкозы клетками, таких как GLUT4 и гексокиназа II. [24] [25] Считается, что они частично опосредуются AMPK, когда он активируется. [26] [27] Кроме того, недавние открытия предположительно могут указывать на прямую роль AMPK в увеличении кровоснабжения тренируемых/тренированных мышечных клеток путем стимуляции и стабилизации как васкулогенеза , так и ангиогенеза . [28] В совокупности эти адаптации , скорее всего, происходят в результате как временного, так и постоянного повышения активности AMPK, вызванного увеличением соотношения AMP:ATP во время отдельных тренировок и длительных тренировок.
Во время одной тренировки AMPK позволяет сокращающимся мышечным клеткам адаптироваться к энергетическим нагрузкам за счет увеличения экспрессии гексокиназы II. [23] транслокация GLUT4 на плазматическую мембрану , [29] [30] [31] [32] для поглощения глюкозы и путем стимуляции гликолиза. [33] Если тренировки продолжаются в рамках длительного режима тренировок , AMPK и другие сигналы будут способствовать адаптации мышечных сокращений, сопровождая активность мышечных клеток к метаболическому переходу, что приводит к окислению жирных кислот для выработки АТФ, а не к гликолитическому подходу. AMPK осуществляет этот переход к окислительному режиму метаболизма путем повышения регуляции и активации окислительных ферментов, таких как гексокиназа II , PPAR-α , PPAR-δ , PGC-1 , UCP-3 , цитохром C и TFAM . [26] [23] [25] [34] [35] [36] [37]
Мутации в канале высвобождения кальция скелетных мышц ( RYR1 ) лежат в основе опасной для жизни реакции на тепло у пациентов с предрасположенностью к злокачественной гипертермии (MHS). При остром воздействии тепла эти мутации вызывают неконтролируемый Ca 2+ высвобождение из саркоплазматической сети , что приводит к устойчивым мышечным контрактурам, тяжелой гипертермии и внезапной смерти. [38] В базальных условиях температурозависимая Ca 2+ утечка также приводит к увеличению потребности в энергии и активации киназы AMP, чувствительной к энергии (AMPK) в скелетных мышцах. [38] Активированная AMPK увеличивает метаболическую активность мышц, включая гликолиз, что приводит к заметному повышению уровня циркулирующего лактата . [38]
LKB1/MO25/STRAD Активность AMPK увеличивается при физической нагрузке, и комплекс считается основным вышестоящим AMPKK 5'-AMP-активируемой протеинкиназы, фосфорилирующей α-субъединицу AMPK по Thr-172. [10] [39] [40] [17] Этот факт озадачивает, учитывая, что, хотя белка было показано, что содержание AMPK в скелетной ткани увеличивается при тренировках на выносливость , было показано, что уровень его активности снижается при тренировках на выносливость как в тренированных, так и в нетренированных тканях. [41] [42] [43] [44] В настоящее время активность AMPK сразу после двухчасовой тренировки у крыс, тренированных на выносливость, неясна. Вполне возможно, что существует прямая связь между наблюдаемым снижением активности AMPK в скелетных мышцах, тренируемых на выносливость, и очевидным снижением реакции AMPK на тренировку при тренировке на выносливость.
Хотя считается, что активация AMPKα2 важна для адаптации митохондрий к тренировкам, недавнее исследование, изучающее реакцию на тренировку у мышей с нокаутом AMPKα2, противоречит этой идее. [45] В их исследовании сравнивалась реакция на тренировку нескольких белков и ферментов у мышей дикого типа и мышей с нокаутом AMPKα2. И хотя у нокаутных мышей были более низкие базальные маркеры плотности митохондрий (COX-1, CS и HAD), после тренировки эти маркеры увеличивались так же, как и у мышей дикого типа. Эти результаты подтверждаются другим исследованием, также показывающим отсутствие различий в адаптации митохондрий к тренировкам между мышами дикого типа и нокаутными мышами. [46]
Максимальная продолжительность жизни
[ редактировать ]необходим для и его коллеги показали, что гомолог AMPK C. elegans, aak-2, Майкл Ристоу продления продолжительности жизни в состояниях ограничения глюкозы, опосредующих процесс, называемый митогормезисом . [47]
Липидный обмен
[ редактировать ]Одним из эффектов физических упражнений является увеличение метаболизма жирных кислот , что обеспечивает больше энергии клетке . Одним из ключевых путей регуляции окисления жирных кислот AMPK является фосфорилирование и инактивация ацетил-КоА-карбоксилазы . [28] Ацетил-КоА-карбоксилаза (АСС) превращает ацетил-КоА в малонил-КоА , ингибитор карнитин-пальмитоилтрансферазы 1 ( CPT-1 ). CPT-1 транспортирует жирные кислоты в митохондрии для окисления . Таким образом, инактивация АСС приводит к усилению транспорта жирных кислот и последующему окислению. Также считается, что снижение малонил-КоА происходит в результате действия малонил-КоА-декарбоксилазы (MCD), которая может регулироваться AMPK. [21] MCD является антагонистом АСС, декарбоксилируя малонил-КоА в ацетил-КоА, что приводит к снижению малонил-КоА и увеличению окисления CPT-1 и жирных кислот.AMPK также играет важную роль в метаболизме липидов в печени . Давно известно, что печеночная АСС регулируется в печени путем фосфорилирования . [22] AMPK также фосфорилирует и инактивирует 3-гидрокси-3-метилглутарил-КоА-редуктазу (HMGCR), ключевой фермент в синтезе холестерина . [29] HMGR преобразует 3-гидрокси-3-метилглутарил-КоА, который производится из ацетил-КоА, в мевалоновую кислоту , которая затем проходит еще несколько этапов метаболизма и превращается в холестерин . Таким образом, AMPK помогает регулировать окисление жирных кислот и синтез холестерина.
Транспорт глюкозы
[ редактировать ]Инсулин – это гормон , который помогает регулировать уровень глюкозы в организме. Когда уровень глюкозы в крови высокий, инсулин высвобождается из островков Лангерганса . Инсулин, среди прочего, будет способствовать поглощению глюкозы клетками посредством увеличения экспрессии и транслокации переносчика глюкозы GLUT-4 . [27] Однако в условиях физических упражнений уровень сахара в крови не обязательно высок, а инсулин не обязательно активируется, однако мышцы все равно способны доставлять глюкозу. AMPK, по-видимому, частично ответственен за поглощение глюкозы, вызванное физическими упражнениями . Гудиер и др. [24] наблюдали, что при физической нагрузке концентрация GLUT-4 увеличивается в плазматической мембране , но снижается в микросомальных мембранах, что позволяет предположить, что физические упражнения облегчают транслокацию везикулярного GLUT-4 на плазматическую мембрану . В то время как интенсивные физические упражнения увеличивают транслокацию GLUT-4, тренировки на выносливость увеличивают общее количество доступного белка GLUT-4. [25] Было показано, что как электрическое сокращение, так и обработка рибонуклеотидом AICA (AICAR) увеличивают активацию AMPK, поглощение глюкозы и транслокацию GLUT-4 в перфузируемых мышцах задних конечностей крыс , связывая поглощение глюкозы, вызванное физической нагрузкой, с AMPK. [48] [23] [34] Хронические инъекции AICAR, имитирующие некоторые эффекты тренировок на выносливость , также увеличивают общее количество белка GLUT-4 в мышечных клетках . [35]
Два белка необходимы для регуляции экспрессии GLUT-4 на уровне транскрипции — фактор 2 энхансера миоцитов ( MEF2 ) и фактор энхансера GLUT4 (GEF). Мутации в участках связывания ДНК любого из этих белков приводят к исчезновению GLUT экспрессии трансгена -4. [30] [31] Эти результаты побудили провести исследование в 2005 году, которое показало, что AMPK непосредственно фосфорилирует GEF, но, похоже, не активирует напрямую MEF2. [32] Однако было показано, что обработка AICAR увеличивает транспорт обоих белков в ядро , а также увеличивает связывание обоих с областью промотора GLUT-4 . [32]
еще один белок, участвующий в углеводном обмене Наряду с GLUT-4 стоит упомянуть . Фермент гексокиназа фосфорилирует шестиуглеродный сахар, особенно глюкозу , что является первой стадией гликолиза . Когда глюкоза транспортируется в клетку, она фосфорилируется гексокиназой. Это фосфорилирование удерживает глюкозу от выхода из клетки и, изменяя структуру глюкозы посредством фосфорилирования, снижает концентрацию молекул глюкозы, поддерживая градиент для транспортировки большего количества глюкозы в клетку. гексокиназы II Транскрипция увеличивается как в красных, так и в белых скелетных мышцах при лечении AICAR. [36] При хронических инъекциях AICAR общее содержание белка гексокиназы II в крыс увеличивается. скелетных мышцах [49]
Митохондрии
[ редактировать ]Митохондриальные ферменты, такие как цитохром С , сукцинатдегидрогеназа , малатдегидрогеназа , α-кетоглутаратдегидрогеназа и цитратсинтаза , увеличивают экспрессию и активность в ответ на физическую нагрузку. [43] Стимуляция AICAR AMPK увеличивает цитохром c и δ-аминолевулинатсинтазу ( ALAS ), фермент, ограничивающий скорость, участвующий в производстве гема . Малатдегидрогеназа и сукцинатдегидрогеназа также увеличиваются, а также активность цитратсинтазы у крыс, получавших инъекции AICAR. [44] И наоборот, у мышей с нокаутом LKB1 наблюдается снижение активности цитохрома с и цитратсинтазы, даже если мыши «тренируются» произвольными упражнениями. [50]
AMPK необходим для увеличения экспрессии рецептора γ-коактиватора-1α ( PGC-1α ), активируемого пролифератором пероксисом, в скелетных мышцах в ответ на истощение запасов креатина . [51] PGC-1α является регулятором транскрипции генов, участвующих в окислении жирных кислот , глюконеогенезе , и считается главным регулятором митохондриального биогенеза . [52]
Для этого он усиливает активность факторов транскрипции, таких как ядерный респираторный фактор 1 ( NRF-1 ), миоцитов фактор 2 усилителя (MEF2), фактор клетки-хозяина (HCF) и других. [9] [33] Он также имеет петлю положительной обратной связи , улучшающую его собственное выражение. [53] И MEF2, и элемент ответа цАМФ ( CRE ) необходимы для индуцируемой сокращением активности промотора PGC-1α . [33] У мышей с нокаутом LKB1 наблюдается снижение PGC-1α, а также митохондриальных белков. [50] [54]
Гормон щитовидной железы
[ редактировать ]AMPK и щитовидной железы гормон регулируют некоторые схожие процессы. Зная об этих сходствах, Winder и Hardie et al. разработали эксперимент, чтобы выяснить, влияет ли на AMPK гормон щитовидной железы . [55] [56] [57] Они обнаружили, что все субъединицы AMPK были увеличены в скелетных мышцах , особенно в камбаловидной и красной четырехглавой мышце, при лечении гормонами щитовидной железы. Также наблюдалось увеличение фосфо-АСС, маркера активности AMPK. [55]
Системы измерения глюкозы
[ редактировать ]Сообщалось, что потеря AMPK изменяет чувствительность клеток, чувствительных к глюкозе, посредством плохо определенных механизмов. Потеря субъединицы AMPKα2 в β-клетках поджелудочной железы и нейронах гипоталамуса снижает чувствительность этих клеток к изменениям внеклеточной концентрации глюкозы. [58] [59] [60] [61] Более того, воздействие на крыс повторяющихся приступов инсулин-индуцированной гипогликемии /глюкопении снижает активацию AMPK в гипоталамусе , одновременно подавляя контррегуляторную реакцию на гипогликемию. [62] [63] Фармакологическая активация AMPK путем доставки активирующего AMPK препарата AICAR непосредственно в гипоталамус может усилить контррегуляторный ответ на гипогликемию. [64]
Лизосомальное повреждение, воспалительные заболевания и метформин
[ редактировать ]AMPK рекрутируется в лизосомы и регулируется в лизосомах через несколько систем, имеющих клиническое значение. Сюда входит комплекс AXIN - LKB1 , действующий в ответ на ограничение глюкозы, функционирующий независимо от чувствительности АМФ, который определяет низкий уровень глюкозы как отсутствие фруктозо-1,6-бисфосфата посредством динамического набора взаимодействий между лизосомально локализованной V-АТФазой и альдолазой при контакте. в эндоплазматическом ретикулуме локализуется ТРПВ . [65] Вторая система управления AMPK [66] локализация в лизосомах зависит от системы галектин-9 - ТАК1 и реакций убиквитинирования, контролируемых деубиквитинирующими ферментами, такими как USP9X, что приводит к активации AMPK в ответ на лизосомальное повреждение, [66] состояние, которое может возникать биохимически, физически через белковые агрегаты, такие как протеопатический тау при болезни Альцгеймера , [67] [68] кристаллический кремнезем, вызывающий силикоз , [68] кристаллы холестерина, вызывающие воспаление через NLRP3 инфламмасому и разрыв атеросклеротических поражений, [69] кристаллы уратов , связанные с подагрой или во время микробной инвазии, такой как Mycobacterium Tuberculosis [68] [70] или коронавирусы, вызывающие атипичную пневмонию . [71] Обе вышеперечисленные лизосомально локализованные системы, контролирующие AMPK, активируют ее в ответ на метформин . [66] [72] широко назначаемый противодиабетический препарат.
Подавление и продвижение опухоли
[ редактировать ]Некоторые данные указывают на то, что AMPK может играть роль в подавлении опухоли. Исследования показали, что AMPK может проявлять большую часть или даже все противоопухолевые свойства киназы печени B1 (LKB1). [17] Кроме того, исследования, в которых активатор AMPK метформин использовался для лечения диабета, обнаружили корреляцию со снижением риска развития рака по сравнению с другими лекарствами. Исследования нокаута и нокдауна генов на мышах показали, что мыши без гена, экспрессирующего AMPK, имели больший риск развития лимфом, хотя, поскольку ген был нокаутирован глобально, а не только в В-клетках , было невозможно заключить, что нокаут AMP имел клеточно-автономный характер. Эффекты внутри опухолевых клеток-предшественников. [73]
Напротив, некоторые исследования связали AMPK с ролью промотора опухоли, защищая раковые клетки от стресса. Таким образом, как только в организме образуются раковые клетки, AMPK может переключиться с защиты от рака на защиту самого рака. Исследования показали, что опухолевые клетки с нокаутом AMPK более подвержены гибели из-за голодания глюкозы или отслойки внеклеточного матрикса , что может указывать на то, что AMPK играет роль в предотвращении этих двух последствий. [73] Недавнее исследование рака поджелудочной железы предполагает, что AMPKα может играть роль в метастатическом каскаде и фенотипе раковых клеток. С механистической точки зрения авторы предполагают, что в отсутствие AMPKα клетки рака поджелудочной железы более уязвимы к окислительному стрессу, что поддерживает функцию AMPKα, способствующую развитию опухолей. [74]
Споры по поводу роли в адаптации к упражнениям/тренировкам
[ редактировать ]Этот раздел написан как личное размышление, личное эссе или аргументативное эссе , в котором излагаются личные чувства редактора Википедии или представлены оригинальные аргументы по определенной теме. ( январь 2021 г. ) |
Кажущаяся парадоксальной роль AMPK проявляется, когда мы более внимательно рассмотрим фермент, чувствительный к энергии, в отношении физических упражнений и длительных тренировок. Подобно шкале краткосрочных интенсивных тренировок, долгосрочные исследования тренировок на выносливость также показывают увеличение окислительных метаболических ферментов, GLUT-4, размера и количества митохондрий, а также повышенную зависимость от окисления жирных кислот; однако Winder et al. сообщили в 2002 году, что, несмотря на наблюдение этих повышенных окислительных биохимических адаптаций к длительным тренировкам на выносливость (аналогично упомянутым выше), реакция AMPK (активация AMPK с началом тренировки) на острые приступы упражнений снизилась в красных квадрицепсах (RQ). с обучением (3 – см. рис.1). И наоборот, в исследовании не было обнаружено таких же результатов в белых четырехглавых мышцах (WQ) и камбаловидной мышце (SOL), как в RQ. Обученные крысы , использованные в этом исследовании на выносливость, бегали на беговых дорожках 5 дней в неделю в течение двух часовых занятий, утром и днем . Крысы также бегали со скоростью до 31 м/мин (степень 15%). Наконец, после тренировки крыс умерщвляли либо в состоянии покоя, либо после 10-минутной тренировки.
Поскольку реакция AMPK на физические упражнения снижается с увеличением продолжительности тренировки, возникает множество вопросов, которые ставят под сомнение роль AMPK в отношении биохимической адаптации к физическим нагрузкам и тренировкам на выносливость. Частично это связано с заметным усилением митохондриального биогенеза , активацией GLUT-4 , UCP-3 , гексокиназы II, а также других метаболических и митохондриальных ферментов, несмотря на снижение активности AMPK при тренировке. Вопросы также возникают потому, что скелетных мышц клетки , которые демонстрируют снижение активности AMPK в ответ на тренировки на выносливость, также, по-видимому, поддерживают окислительно-зависимый подход к метаболизму, который, как полагают, также в некоторой степени регулируется активностью AMPK. [34] [35]
Если реакция AMPK на физические упражнения частично ответственна за биохимическую адаптацию к тренировкам, как тогда можно поддерживать эту адаптацию к тренировкам, если реакция AMPK на физические упражнения ослабляется в процессе тренировки? Предполагается, что эта адаптивная роль к тренировке поддерживается активностью AMPK и что увеличение активности AMPK в ответ на упражнения в тренированных скелетных мышцах еще не наблюдалось из-за биохимических адаптаций, которые сама тренировка стимулировала в мышечной ткани , чтобы уменьшить метаболическая потребность в активации AMPK. Другими словами, из-за предыдущей адаптации к тренировке AMPK не будет активирована, и дальнейшая адаптация не произойдет до тех пор, пока внутриклеточные уровни АТФ не истощатся из-за энергетического воздействия еще более высокой интенсивности, чем до этих предыдущих адаптаций.
См. также
[ редактировать ]Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Сутер М., Риек У., Тюрк Р., Шлаттнер У., Валлиманн Т., Нойманн Д. (октябрь 2006 г.). «Анализ роли 5'-АМФ в аллостерической стимуляции, активации и деактивации АМФ-активируемой протеинкиназы» . Журнал биологической химии . 281 (43): 32207–16. дои : 10.1074/jbc.M606357200 . ПМИД 16943194 .
- ^ Уиндер В.В., Харди Д.Г. (июль 1999 г.). «АМФ-активируемая протеинкиназа, главный метаболический переключатель: возможная роль при диабете 2 типа». Американский журнал физиологии . 277 (1): Е1-10. дои : 10.1152/ajpendo.1999.277.1.E1 . ПМИД 10409121 .
- ^ Хэллоус К.Р., Альзамора Р., Ли Х., Гонг Ф., Смолак С., Нойманн Д., Пастор-Солер Н.М. (апрель 2009 г.). «АМФ-активируемая протеинкиназа ингибирует щелочное рН и РКА-индуцированное накопление апикальной вакуолярной Н+-АТФазы в прозрачных клетках придатка яичка» . Американский журнал физиологии. Клеточная физиология . 296 (4): C672-81. doi : 10.1152/ajpcell.00004.2009 . ПМК 2670645 . ПМИД 19211918 .
- ^ Перейти обратно: а б Стэплтон Д., Митчелхилл К.И., Гао Г., Видмер Дж., Мичелл Б.Дж., Тех Т. и др. (январь 1996 г.). «Подсемейство AMP-активируемых протеинкиназ млекопитающих» . Журнал биологической химии . 271 (2): 611–4. дои : 10.1074/jbc.271.2.611 . ПМИД 8557660 .
- ^ Риек У., Шольц Р., Конарев П., Руфер А., Сутер М., Назабал А. и др. (июнь 2008 г.). «Структурные свойства AMP-активируемой протеинкиназы: димеризация, молекулярная форма и изменения при связывании лиганда» . Журнал биологической химии . 283 (26): 18331–18343. дои : 10.1074/jbc.M708379200 . ПМИД 18372250 .
- ^ Перейти обратно: а б с д и Ян Ю, Мукерджи С., Харикумар К.Г., Струценберг Т.С., Чжоу К.Э., Суино-Пауэлл К. и др. (июль 2021 г.). «Структура комплекса AMPK в неактивном, связанном с АТФ состоянии» . Наука . 373 (6553): 413–419. Бибкод : 2021Sci...373..413Y . дои : 10.1126/science.abe7565 . ПМЦ 8428800 . ПМИД 34437114 .
- ^ Окхилл Дж.С., Чен З.П., Скотт Дж.В., Стил Р., Кастелли Л.А., Линг Н. и др. (ноябрь 2010 г.). «Миристоилирование β-субъединицы является привратником для инициации восприятия метаболического стресса с помощью AMP-активируемой протеинкиназы (AMPK)» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (45): 19237–19241. Бибкод : 2010PNAS..10719237O . дои : 10.1073/pnas.1009705107 . ПМЦ 2984171 . ПМИД 20974912 .
- ^ Чен Л., Ван Дж., Чжан Ю.Ю., Ян С.Ф., Нейман Д., Шлаттнер У. и др. (июнь 2012 г.). «АМФ-активируемая протеинкиназа претерпевает нуклеотид-зависимые конформационные изменения». Структурная и молекулярная биология природы . 19 (7): 716–718. дои : 10.1038/nsmb.2319 . ПМИД 22659875 . S2CID 13591617 .
- ^ Перейти обратно: а б Адамс Дж., Чен З.П., Ван Дендерен Б.Дж., Мортон С.Дж., Паркер М.В., Уиттерс Л.А. и др. (январь 2004 г.). «Внутристерический контроль AMPK через аллостерический регуляторный сайт субъединицы гамма1 AMP» . Белковая наука . 13 (1): 155–65. дои : 10.1110/ps.03340004 . ПМК 2286513 . ПМИД 14691231 .
- ^ Перейти обратно: а б Хоули С.А., Дэвисон М., Вудс А., Дэвис С.П., Бери Р.К., Карлинг Д., Харди Д.Г. (ноябрь 1996 г.). «Характеристика AMP-активируемой протеинкиназы из печени крыс и идентификация треонина 172 как основного сайта, в котором он фосфорилирует AMP-активируемую протеинкиназы» . Журнал биологической химии . 271 (44): 27879–87. дои : 10.1074/jbc.271.44.27879 . ПМИД 8910387 .
- ^ Торнтон С., Сноуден М.А., Карлинг Д. (май 1998 г.). «Идентификация новой изоформы бета-субъединицы АМФ-активируемой протеинкиназы, которая высоко экспрессируется в скелетных мышцах» . Журнал биологической химии . 273 (20): 12443–50. дои : 10.1074/jbc.273.20.12443 . ПМИД 9575201 .
- ^ Чунг П.С., Солт И.П., Дэвис С.П., Харди Д.Г., Карлинг Д. (март 2000 г.). «Характеристика изоформ гамма-субъединицы АМФ-активируемой протеинкиназы и их роли в связывании АМФ» . Биохимический журнал . 346 (3): 659–69. дои : 10.1042/0264-6021:3460659 . ПМЦ 1220898 . ПМИД 10698692 .
- ^ Сяо Б., Хит Р., Сайу П., Лейпер Ф.К., Леоне П., Цзин С. и др. (сентябрь 2007 г.). «Структурная основа связывания AMP с AMP-активируемой протеинкиназой млекопитающих». Природа . 449 (7161): 496–500. Бибкод : 2007Natur.449..496X . дои : 10.1038/nature06161 . ПМИД 17851531 . S2CID 4345919 .
- ^ Сяо Б., Сандерс М.Дж., Андервуд Е., Хит Р., Майер Ф.В., Кармена Д. и др. (апрель 2011 г.). «Структура АМПК млекопитающих и ее регуляция с помощью АДФ» . Природа . 472 (7342): 230–3. Бибкод : 2011Natur.472..230X . дои : 10.1038/nature09932 . ПМК 3078618 . ПМИД 21399626 .
- ^ Чен Л., Ван Дж., Чжан Ю.Ю., Ян С.Ф., Нейман Д., Шлаттнер У. и др. (июнь 2012 г.). «АМФ-активируемая протеинкиназа претерпевает нуклеотид-зависимые конформационные изменения». Структурная и молекулярная биология природы . 19 (7): 716–8. дои : 10.1038/nsmb.2319 . ПМИД 22659875 . S2CID 13591617 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д Чон СМ (июль 2016 г.). «Регуляция и функция AMPK в физиологии и заболеваниях» . Экспериментальная и молекулярная медицина . 48 (7): е245. дои : 10.1038/эмм.2016.81 . ПМЦ 4973318 . ПМИД 27416781 .
- ^ Перейти обратно: а б с Вудс А., Джонстон С.Р., Дикерсон К., Лейпер Ф.К., Фрайер Л.Г., Нейман Д. и др. (ноябрь 2003 г.). «LKB1 представляет собой вышестоящую киназу в AMP-активируемом протеинкиназном каскаде» . Современная биология . 13 (22): 2004–8. дои : 10.1016/j.cub.2003.10.031 . ПМИД 14614828 .
- ^ Калабрезе М.Ф., Раджамохан Ф., Харрис М.С., Касперс Н.Л., Мадьяр Р., Витка Дж.М. и др. (август 2014 г.). «Структурная основа активации AMPK: природные и синтетические лиганды регулируют активность киназы с противоположных полюсов с помощью разных молекулярных механизмов» . Структура . 22 (8): 1161–1172. дои : 10.1016/j.str.2014.06.009 . ПМИД 25066137 .
- ^ Сяо Б., Сандерс М.Дж., Кармена Д., Брайт Н.Дж., Хайр Л.Ф., Андервуд Е. и др. (декабрь 2013 г.). «Структурные основы регуляции AMPK низкомолекулярными активаторами» . Природные коммуникации . 4 (1): 3017. Бибкод : 2013NatCo...4.3017X . дои : 10.1038/ncomms4017 . ПМЦ 3905731 . ПМИД 24352254 .
- ^ Харди Д.Г., Росс Ф.А., Хоули С.А. (март 2012 г.). «AMPK: датчик питательных веществ и энергии, поддерживающий энергетический гомеостаз» . Обзоры природы. Молекулярно-клеточная биология . 13 (4): 251–62. дои : 10.1038/nrm3311 . ПМК 5726489 . ПМИД 22436748 .
- ^ Перейти обратно: а б Бержерон Р., Рен Дж.М., Кэдман К.С., Мур И.К., Перрет П., Пайпарт М. и др. (декабрь 2001 г.). «Хроническая активация киназы AMP приводит к активации NRF-1 и митохондриальному биогенезу». Американский журнал физиологии. Эндокринология и обмен веществ . 281 (6): E1340-6. дои : 10.1152/ajpendo.2001.281.6.e1340 . ПМИД 11701451 . S2CID 21577702 .
- ^ Перейти обратно: а б Зонг Х., Рен Дж.М., Янг Л.Х., Пайпарт М., Му Дж., Бирнбаум М.Дж., Шульман Г.И. (декабрь 2002 г.). «АМФ-киназа необходима для митохондриального биогенеза в скелетных мышцах в ответ на хроническое лишение энергии» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (25): 15983–7. Бибкод : 2002PNAS...9915983Z . дои : 10.1073/pnas.252625599 . ПМЦ 138551 . ПМИД 12444247 .
- ^ Перейти обратно: а б с д Холмс Б.Ф., Курт-Крачек Э.Дж., Уиндер В.В. (ноябрь 1999 г.). «Хроническая активация 5'-АМФ-активируемой протеинкиназы увеличивает GLUT-4, гексокиназу и гликоген в мышцах». Журнал прикладной физиологии . 87 (5): 1990–5. doi : 10.1152/яп.1999.87.5.1990 . ПМИД 10562646 . S2CID 9622267 .
- ^ Перейти обратно: а б Оджука Э.О., Джонс Т.Э., Нолте Л.А., Чен М., Вамхофф Б.Р., Стурек М., Холлоши Д.О. (май 2002 г.). «Регуляция биогенеза GLUT4 в мышцах: доказательства участия AMPK и Ca (2+)». Американский журнал физиологии. Эндокринология и обмен веществ . 282 (5): E1008-13. дои : 10.1152/ajpendo.00512.2001 . ПМИД 11934664 . S2CID 7891060 .
- ^ Перейти обратно: а б с Стоппани Дж., Хильдебрандт А.Л., Сакамото К., Кэмерон-Смит Д., Гудиер Л.Дж., Нойфер П.Д. (декабрь 2002 г.). «АМФ-активируемая протеинкиназа активирует транскрипцию генов UCP3 и HKII в скелетных мышцах крыс». Американский журнал физиологии. Эндокринология и обмен веществ . 283 (6): E1239-48. дои : 10.1152/ajpendo.00278.2002 . ПМИД 12388122 . S2CID 27188844 .
- ^ Перейти обратно: а б Оджука Э.О. (май 2004 г.). «Роль киназы кальция и AMP в регуляции биогенеза митохондрий и уровня GLUT4 в мышцах» . Труды Общества питания . 63 (2): 275–8. дои : 10.1079/PNS2004339 . hdl : 11427/35025 . ПМИД 15294043 .
- ^ Перейти обратно: а б Уиндер В.В., Холмс Б.Ф., Рубинк Д.С., Дженсен Э.Б., Чен М., Холлоши Д.О. (июнь 2000 г.). «Активация АМФ-активируемой протеинкиназы увеличивает количество митохондриальных ферментов в скелетных мышцах». Журнал прикладной физиологии . 88 (6): 2219–26. дои : 10.1152/яп.2000.88.6.2219 . ПМИД 10846039 . S2CID 5627995 .
- ^ Перейти обратно: а б Оучи Н., Шибата Р., Уолш К. (апрель 2005 г.). «AMP-активируемая передача сигналов протеинкиназы стимулирует экспрессию VEGF и ангиогенез в скелетных мышцах» . Исследование кровообращения . 96 (8): 838–46. дои : 10.1161/01.RES.0000163633.10240.3b . ПМИД 15790954 .
- ^ Перейти обратно: а б Хаяши Т., Хиршман М.Ф., Курт Э.Дж., Уиндер В.В., Goodyear LJ (август 1998 г.). «Доказательства опосредования 5'-АМФ-активируемой протеинкиназой влияния мышечного сокращения на транспорт глюкозы». Диабет . 47 (8): 1369–73. дои : 10.2337/диабет.47.8.1369 . ПМИД 9703344 .
- ^ Перейти обратно: а б Хаяши Т., Хиршман М.Ф., Фуджи Н., Хабиновски С.А., Уиттерс Л.А., Goodyear LJ (апрель 2000 г.). «Метаболический стресс и измененный транспорт глюкозы: активация AMP-активируемой протеинкиназы как объединяющий механизм сцепления» . Диабет . 49 (4): 527–31. дои : 10.2337/диабет.49.4.527 . ПМИД 10871188 .
- ^ Перейти обратно: а б Курт-Крачек Э.Дж., Хиршман М.Ф., Гудиер Л.Дж., Уиндер В.В. (август 1999 г.). «Активация 5'-АМФ-активируемой протеинкиназы вызывает транслокацию GLUT4 в скелетных мышцах». Диабет . 48 (8): 1667–71. дои : 10.2337/диабет.48.8.1667 . ПМИД 10426389 . S2CID 29891424 .
- ^ Перейти обратно: а б с Меррилл Г.Ф., Курт Э.Дж., Харди Д.Г., Уиндер В.В. (декабрь 1997 г.). «Рибозид AICA увеличивает AMP-активируемую протеинкиназу, окисление жирных кислот и поглощение глюкозы в мышцах крысы». Американский журнал физиологии . 273 (6): E1107-12. дои : 10.1152/ajpendo.1997.273.6.E1107 . ПМИД 9435525 .
- ^ Перейти обратно: а б с Марсин А.С., Бертран Л., Райдер М.Х., Депре Дж., Болуайе С., Винсент М.Ф. и др. (октябрь 2000 г.). «Фосфорилирование и активация сердечной PFK-2 с помощью AMPK играет роль в стимуляции гликолиза во время ишемии» . Современная биология . 10 (20): 1247–55. дои : 10.1016/S0960-9822(00)00742-9 . ПМИД 11069105 .
- ^ Перейти обратно: а б с Ли У.Дж., Ким М., Пак Х.С., Ким Х.С., Чон М.Дж., О К.С. и др. (февраль 2006 г.). «Активация AMPK увеличивает окисление жирных кислот в скелетных мышцах за счет активации PPARalpha и PGC-1». Связь с биохимическими и биофизическими исследованиями . 340 (1): 291–5. дои : 10.1016/j.bbrc.2005.12.011 . ПМИД 16364253 .
- ^ Перейти обратно: а б с Сува М., Эгашира Т., Накано Х., Сасаки Х., Кумагай С. (декабрь 2006 г.). «Метформин увеличивает активность белка PGC-1альфа и окислительных ферментов, возможно, за счет фосфорилирования AMPK в скелетных мышцах in vivo». Журнал прикладной физиологии . 101 (6): 1685–92. doi : 10.1152/japplphysicalol.00255.2006 . ПМИД 16902066 . S2CID 17877895 .
- ^ Перейти обратно: а б Оджука Э.О., Нолте Л.А., Холлоши Д.О. (март 2000 г.). «Повышение экспрессии GLUT-4 и гексокиназы в эпитрохлеарной мышце крысы, подвергшейся воздействию AICAR in vitro». Журнал прикладной физиологии . 88 (3): 1072–5. дои : 10.1152/яп.2000.88.3.1072 . ПМИД 10710405 . S2CID 22504819 .
- ^ Наркар В.А., Даунс М., Ю РТ, Эмблер Е., Ван YX, Банайо Е. и др. (август 2008 г.). «Агонисты AMPK и PPARdelta являются миметиками упражнений» . Клетка . 134 (3): 405–15. дои : 10.1016/j.cell.2008.06.051 . ПМК 2706130 . ПМИД 18674809 .
- ^ Перейти обратно: а б с Ван Х.Дж., Ли К.С., Йи Р.С., Грум Л., Фридман И., Бэбкок Л. и др. (октябрь 2020 г.). «Адаптивный термогенез усиливает опасную для жизни реакцию на тепло у мышей с мутацией Ryr1» . Природные коммуникации . 11 (1): 5099. Бибкод : 2020NatCo..11.5099W . дои : 10.1038/s41467-020-18865-z . ПМК 7547078 . ПМИД 33037202 .
- ^ Стейн С.К., Вудс А., Джонс Н.А., Дэвисон М.Д., Карлинг Д. (февраль 2000 г.). «Регуляция AMP-активируемой протеинкиназы путем фосфорилирования» . Биохимический журнал . 345 (3): 437–43. дои : 10.1042/0264-6021:3450437 . ПМК 1220775 . ПМИД 10642499 .
- ^ Хоули С.А., Будо Дж., Рид Дж.Л., Мастард К.Дж., Удд Л., Мякеля Т.П. и др. (2003). «Комплексы между опухолевым супрессором LKB1, STRAD альфа/бета и MO25 альфа/бета являются вышестоящими киназами в AMP-активируемом протеинкиназном каскаде» . Журнал биологии . 2 (4): 28. дои : 10.1186/1475-4924-2-28 . ПМК 333410 . ПМИД 14511394 .
- ^ Херст Д., Тейлор Э.Б., Клайн Т.Д., Гринвуд Л.Дж., Комптон К.Л., Лэмб Дж.Д., Уиндер В.В. (октябрь 2005 г.). «Активность АМФ-активируемой протеинкиназы и фосфорилирование АМФ-активируемой протеинкиназы при сокращении мышц малоподвижных и тренированных на выносливость крыс» . Американский журнал физиологии. Эндокринология и обмен веществ . 289 (4): Е710-5. дои : 10.1152/ajpendo.00155.2005 . ПМИД 15928023 . S2CID 25296834 .
- ^ Хатбер Калифорния, Харди Д.Г., Уиндер В.В. (февраль 1997 г.). «Электрическая стимуляция инактивирует мышечную ацетил-КоА-карбоксилазу и увеличивает AMP-активируемую протеинкиназу». Американский журнал физиологии . 272 (2 ч. 1): E262-6. дои : 10.1152/ajpendo.1997.272.2.E262 . ПМИД 9124333 .
- ^ Перейти обратно: а б Тейлор Э.Б., Херст Д., Гринвуд Л.Дж., Лэмб Дж.Д., Клайн Т.Д., Судвикс С.Н., Уиндер В.В. (декабрь 2004 г.). «Тренировка на выносливость увеличивает белок LKB1 и MO25, но не активность AMP-активируемой протеинкиназы в скелетных мышцах». Американский журнал физиологии. Эндокринология и обмен веществ . 287 (6): E1082-9. дои : 10.1152/ajpendo.00179.2004 . ПМИД 15292028 .
- ^ Перейти обратно: а б Тейлор Э.Б., Лэмб Дж.Д., Херст Р.В., Чессер Д.Г., Эллингсон В.Дж., Гринвуд Л.Дж. и др. (декабрь 2005 г.). «Тренировки на выносливость увеличивают количество белков LKB1 и PGC-1альфа в скелетных мышцах: влияние времени и интенсивности». Американский журнал физиологии. Эндокринология и обмен веществ . 289 (6): Е960-8. дои : 10.1152/ajpendo.00237.2005 . ПМИД 16014350 . S2CID 14454264 .
- ^ Йоргенсен С.Б., Трибак Дж.Т., Виоллет Б., Шерлинг П., Волонт С., Войташевски Дж.Ф., Рихтер Э.А. (январь 2007 г.). «Роль AMPKalpha2 в базальной, тренировочной и AICAR-индуцированной экспрессии GLUT4, гексокиназы II и митохондриальных белков в мышцах мыши». Американский журнал физиологии. Эндокринология и обмен веществ . 292 (1): Е331-9. дои : 10.1152/ajpendo.00243.2006 . ПМИД 16954334 .
- ^ Рёкль К.С., Хиршман М.Ф., Брандауэр Дж., Фуджи Н., Виттерс Л.А., Goodyear LJ (август 2007 г.). «Адаптация скелетных мышц к физическим упражнениям: AMP-активируемая протеинкиназа опосредует смену типа мышечных волокон» . Диабет . 56 (8): 2062–9. дои : 10.2337/db07-0255 . ПМИД 17513699 .
- ^ Шульц Т.Дж., Зарс К., Фойгт А., Урбан Н., Бирринджер М., Ристоу М. (октябрь 2007 г.). «Ограничение глюкозы продлевает продолжительность жизни Caenorhabditis elegans, индуцируя митохондриальное дыхание и усиливая окислительный стресс» . Клеточный метаболизм . 6 (4): 280–93. дои : 10.1016/j.cmet.2007.08.011 . ПМИД 17908557 .
- ^ Виндер В.В. (сентябрь 2001 г.). «Ощущение энергии и передача сигналов с помощью AMP-активируемой протеинкиназы в скелетных мышцах». Журнал прикладной физиологии . 91 (3): 1017–28. дои : 10.1152/яп.2001.91.3.1017 . ПМИД 11509493 . S2CID 18290292 .
- ^ Минокоши Ю., Ким Ю.Б., Перони О.Д., Фрайер Л.Г., Мюллер С., Карлинг Д., Кан Б.Б. (январь 2002 г.). «Лептин стимулирует окисление жирных кислот путем активации АМФ-активируемой протеинкиназы». Природа . 415 (6869): 339–43. Бибкод : 2002Natur.415..339M . дои : 10.1038/415339а . ПМИД 11797013 . S2CID 4409274 .
- ^ Перейти обратно: а б Сакамото К., Йоранссон О., Харди Д.Г., Алесси Д.Р. (август 2004 г.). «Активность киназ, связанных с LKB1 и AMPK, в скелетных мышцах: эффекты сокращения, фенформина и AICAR». Американский журнал физиологии. Эндокринология и обмен веществ . 287 (2): Е310-7. дои : 10.1152/ajpendo.00074.2004 . ПМИД 15068958 .
- ^ Минокоши Ю., Алкьер Т., Фурукава Н., Ким Ю.Б., Ли А., Сюэ Б. и др. (апрель 2004 г.). «АМФ-киназа регулирует потребление пищи, реагируя на гормональные и питательные сигналы в гипоталамусе». Природа . 428 (6982): 569–74. Бибкод : 2004Natur.428..569M . дои : 10.1038/nature02440 . ПМИД 15058305 . S2CID 4302317 .
- ^ Тейлор Э.Б., Эллингсон В.Дж., Лэмб Дж.Д., Чессер Д.Г., Уиндер В.В. (июнь 2005 г.). «Длинноцепочечные эфиры ацил-КоА ингибируют фосфорилирование AMP-активируемой протеинкиназы по треонину-172 с помощью LKB1/STRAD/MO25». Американский журнал физиологии. Эндокринология и обмен веществ . 288 (6): E1055-61. дои : 10.1152/ajpendo.00516.2004 . ПМИД 15644453 .
- ^ Харди Д.Г., Хоули С.А. (декабрь 2001 г.). «AMP-активируемая протеинкиназа: новый взгляд на гипотезу энергетического заряда». Биоэссе . 23 (12): 1112–9. дои : 10.1002/bies.10009 . ПМИД 11746230 . S2CID 39876683 .
- ^ Томсон Д.М., Портер Б.Б., Талл Дж.Х., Ким Х.Дж., Барроу-младший, Уиндер В.В. (январь 2007 г.). «Дефицит LKB1 в скелетных мышцах и сердце вызывает снижение произвольного бега и снижение экспрессии мышечных митохондриальных маркерных ферментов у мышей». Американский журнал физиологии. Эндокринология и обмен веществ . 292 (1): Е196-202. дои : 10.1152/ajpendo.00366.2006 . ПМИД 16926377 .
- ^ Перейти обратно: а б Парк, Ш.; Полсен, СР; Гаммон, СР; Горчица, К.Дж.; Харди, генеральный директор; Уиндер, WW (декабрь 2002 г.). «Влияние состояния щитовидной железы на экспрессию АМФ-активируемой протеинкиназы и ацетил-КоА-карбоксилазы в мышцах» . Журнал прикладной физиологии . 93 (6): 2081–2088. doi : 10.1152/japplphysicalol.00504.2002 . ISSN 8750-7587 . ПМИД 12433937 .
- ^ Бранволд, диджей; Оллред, ДР; Бекстед, диджей; Ким, HJ; Филлмор, Н.; Кондон, Б.М.; Браун, доктор юридических наук; Судвикс, С.Н.; Томсон, DM; Уиндер, WW (октябрь 2008 г.). «Влияние гормонов щитовидной железы на LKB1, MO25, фосфо-AMPK, фосфо-CREB и PGC-1альфа в мышцах крыс» . Журнал прикладной физиологии . 105 (4): 1218–1227. doi : 10.1152/japplphysicalol.00997.2007 . ISSN 8750-7587 . ПМИД 18669938 . S2CID 2019764 .
- ^ Виндер, WW; Харди, генеральный директор; Горчица, К.Дж.; Гринвуд, LJ; Пакстон, Бельгия; Парк, Ш.; Рубинк, Д.С.; Тейлор, Э.Б. (февраль 2003 г.). «Долгосрочная регуляция АМФ-активируемой протеинкиназы и ацетил-КоА-карбоксилазы в скелетных мышцах» . Труды Биохимического общества . 31 (Часть 1): 182–185. дои : 10.1042/bst0310182 . ISSN 0300-5127 . ПМИД 12546681 .
- ^ Сан Г., Тарасов А.И., МакГинти Дж., Макдональд А., да Силва Ксавье Г., Горман Т. и др. (май 2010 г.). «Аблация AMP-активируемых протеинкиназ альфа1 и альфа2 из бета-клеток поджелудочной железы мыши и нейронов RIP2.Cre подавляет высвобождение инсулина in vivo» . Диабетология . 53 (5): 924–36. дои : 10.1007/s00125-010-1692-1 . ПМК 4306708 . ПМИД 20221584 .
- ^ Билл С., Пийпари К., Аль-Кассаб Х., Смит М.А., Паркер Н., Карлинг Д. и др. (июль 2010 г.). «Потеря субъединицы АМФ-активируемой протеинкиназы альфа2 в бета-клетках мыши ухудшает стимулируемую глюкозой секрецию инсулина и подавляет их чувствительность к гипогликемии» . Биохимический журнал . 429 (2): 323–33. дои : 10.1042/BJ20100231 . ПМЦ 2895783 . ПМИД 20465544 .
- ^ Кларет М., Смит М.А., Баттерхэм Р.Л., Селман С., Чоудхури А.И., Фрайер Л.Г. и др. (август 2007 г.). «AMPK необходим для регуляции энергетического гомеостаза и восприятия глюкозы нейронами POMC и AgRP» . Журнал клинических исследований . 117 (8): 2325–36. дои : 10.1172/JCI31516 . ЧВК 1934578 . ПМИД 17671657 .
- ^ Билл С., Гамильтон Д.Л., Галлахер Дж., Логи Л., Райт К., Саутар М.П. и др. (сентябрь 2012 г.). «Клетки GT1-7 гипоталамуса мыши демонстрируют AMPK-зависимое внутреннее поведение по чувствительности к глюкозе» . Диабетология . 55 (9): 2432–44. дои : 10.1007/s00125-012-2617-y . ПМК 3411292 . ПМИД 22760787 .
- ^ Алкьер Т., Кавасима Дж., Цудзи Ю., Кан Б.Б. (март 2007 г.). «Роль гипоталамической аденозин-5'-монофосфат-активируемой протеинкиназы в нарушенной контррегуляторной реакции, вызванной повторяющейся нейроглюкопенией» . Эндокринология . 148 (3): 1367–75. дои : 10.1210/en.2006-1039 . ПМИД 17185376 .
- ^ МакКриммон Р.Дж., Шоу М., Фан Икс, Ченг Х., Дин Ю., Велла М.К. и др. (февраль 2008 г.). «Ключевая роль AMP-активируемой протеинкиназы в вентромедиальном гипоталамусе в регуляции контррегуляторных гормональных реакций на острую гипогликемию» . Диабет . 57 (2): 444–50. дои : 10.2337/db07-0837 . ПМИД 17977955 .
- ^ Фан Х, Дин Ю, Браун С., Чжоу Л., Шоу М., Велла М.К. и др. (июнь 2009 г.). «Активация гипоталамической АМФ-активируемой протеинкиназы с помощью AICAR усиливает контррегуляторные реакции на гипогликемию на модели диабета 1 типа на грызунах» . Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 296 (6): Р1702-8. дои : 10.1152/ajpregu.90600.2008 . ПМЦ 2692788 . ПМИД 19357294 .
- ^ Ли М., Чжан К.С., Цзун Ю., Фэн Дж.В., Ма Т., Ху М. и др. (сентябрь 2019 г.). «Переходные потенциальные V-каналы рецепторов необходимы для восприятия глюкозы с помощью альдолазы и AMPK» . Клеточный метаболизм . 30 (3): 508–524.e12. doi : 10.1016/j.cmet.2019.05.018 . ПМК 6720459 . ПМИД 31204282 .
- ^ Перейти обратно: а б с Цзя Дж., Бисса Б., Брехт Л., Аллерс Л., Чой С.В., Гу Ю. и др. (март 2020 г.). «AMPK, регулятор метаболизма и аутофагии, активируется лизосомальным повреждением посредством новой галектин-направленной системы передачи сигнала убиквитина» . Молекулярная клетка . 77 (5): 951–969.е9. дои : 10.1016/j.molcel.2019.12.028 . ПМЦ 7785494 . ПМИД 31995728 .
- ^ Пападопулос С., Киршнер П., Баг М., Грум Д., Кервер Л., Шульце Н. и др. (январь 2017 г.). «VCP/p97 взаимодействует с YOD1, UBXD1 и PLAA, обеспечивая очистку разорвавшихся лизосом посредством аутофагии» . Журнал ЭМБО . 36 (2): 135–150. дои : 10.15252/embj.201695148 . ПМК 5242375 . ПМИД 27753622 .
- ^ Перейти обратно: а б с Цзя Дж., Клод-Топен А., Гу Ю., Чой С.В., Питерс Р., Бисса Б. и др. (январь 2020 г.). «Галектин-3 координирует клеточную систему лизосомального восстановления и удаления» . Развивающая клетка . 52 (1): 69–87.e8. дои : 10.1016/j.devcel.2019.10.025 . ПМК 6997950 . ПМИД 31813797 .
- ^ Гребе А., Латц Э. (март 2013 г.). «Кристаллы холестерина и воспаление» . Текущие отчеты по ревматологии . 15 (3): 313. doi : 10.1007/s11926-012-0313-z . ПМЦ 3623938 . ПМИД 23412688 .
- ^ Чаухан С., Кумар С., Джайн А., Понпуак М., Мадд М.Х., Кимура Т. и др. (октябрь 2016 г.). «TRIMs и галектины сотрудничают во всем мире, а TRIM16 и галектин-3 совместно направляют аутофагию в гомеостазе повреждения эндомембраны» . Развивающая клетка . 39 (1): 13–27. дои : 10.1016/j.devcel.2016.08.003 . ПМК 5104201 . ПМИД 27693506 .
- ^ Юэ Ю, Набар Н.Р., Ши К.С., Каменева О., Сяо X, Хван И.Ю. и др. (сентябрь 2018 г.). «Открытая рамка считывания-3a SARS-коронавируса вызывает мультимодальную некротическую гибель клеток» . Смерть клеток и болезни . 9 (9): 904. doi : 10.1038/s41419-018-0917-y . ПМК 6125346 . ПМИД 30185776 .
- ^ Чжан К.С., Ли М., Ма Т., Цзун Ю., Цуй Дж., Фэн Дж.В. и др. (октябрь 2016 г.). «Метформин активирует AMPK через лизосомальный путь» . Клеточный метаболизм . 24 (4): 521–522. дои : 10.1016/j.cmet.2016.09.003 . ПМИД 27732831 .
- ^ Перейти обратно: а б Генеральный менеджер Меррилл, Курт Э., Харди Д.Г., Уиндер В.В. (2020). «АМФ-активируемая протеинкиназа: друг или враг при раке?» . Ежегодный обзор биологии рака . 4 : 1–16. doi : 10.1146/annurev-cancerbio-030419-033619 .
- ^ Шнайдер, Кэролин; Гильберт, Джорина; Женево, Франциска; Хёфер, Стефани; Краусс, Люк; Шиктанц, Феликс; Контрерас, Констанца Тапиа; Джансари, Шаишави; Папаргириу, Аристейдис; Рихтер, Торстен; Алфайоми, Абдалла М.; Фалькомата, Кьяра; Шневайс, Кристиан; Орбен, Феликс; Оллингер, Руперт (17 июня 2024 г.). «Новый ингибитор AMPK повышает чувствительность клеток рака поджелудочной железы к индукции ферроптоза» . Передовая наука . дои : 10.1002/advs.202307695 . ISSN 2198-3844 .
Внешние ссылки
[ редактировать ]- AMP-активированная+протеин+киназа в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- ЭК 2.7.11.31