МАП2К7
| МАП2К7 | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | MAP2K7 , JNKK2, MAPKK7, MEK, MEK 7, MKK7, PRKMK7, SAPKK-4, SAPKK4, митоген-активируемая протеинкиназа киназа 7 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 603014 ; МГИ : 1346871 ; Гомологен : 56548 ; Генные карты : MAP2K7 ; ОМА : MAP2K7 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||




Киназа протеинкиназы 7, активируемая митогеном, двойной специфичности , также известная как киназа 7 MAP киназы или MKK7 , представляет собой фермент , который у людей кодируется MAP2K7 геном . [ 5 ] Этот белок является членом семейства митоген-активируемых протеинкиназ . Белок MKK7 существует в виде шести различных изоформ с тремя возможными N-концами (изоформы α, β и γ) и двумя возможными C-концами (изоформы 1 и 2). [ 6 ]
МКК7 участвует в передаче сигналов, опосредующих клеточные ответы на провоспалительные цитокины и стрессы окружающей среды . Эта киназа специфически активирует MAPK8 /JNK1 и MAPK9 /JNK2, а сама эта киназа фосфорилируется и активируется киназными киназами MAP-киназы, включая MAP3K1 /MEKK1, MAP3K2 /MEKK2, MAP3K3 /MEKK5 и MAP4K2 /GCK. [ нужна ссылка ]
МКК7 повсеместно экспрессируется во всех тканях. Однако он демонстрирует более высокий уровень экспрессии в скелетных мышцах. [ 7 ] Было обнаружено множество альтернативно сплайсированных вариантов транскриптов, кодирующих различные изоформы. [ 5 ]
Номенклатура
[ редактировать ]MAP2K7 также известен как:
- МКК7
- JNK-активируемая киназа 2
- Киназа 7 MAPK/ERK (MEK7)
- Стресс-активируемая протеинкиназа-киназа 4 (киназа SAPK 4, SAPKK4)
- c-Jun N-концевая киназа 2 (киназа JNK 2, JNKK2)
- Активируемая стрессом/регулируемая внеклеточными сигналами протеинкиназа 2 (SEK2)
Изоформы
[ редактировать ]Мышиный белок MKK7 кодируется 14 экзонами , которые можно альтернативно сплайсировать с образованием группы протеинкиназ . В результате образуются шесть изоформ с тремя возможными N-концами (изоформы α, β и γ) и двумя возможными C-концами (изоформы 1 и 2). Молекулярная масса изоформ составляет от 38 до 52 кДа и содержит от 345 до 467 аминокислот. [ 6 ]
Физиологическая значимость различных изоформ MKK7 до сих пор неясна. Данные показывают, что MKK7α, у которой отсутствует NH 2 -концевое удлинение, демонстрирует более низкую базальную активность в связывании JNK по сравнению с изоформами MKKβ и γ. Повышенная базальная активность β- и γ-изоформ может быть обусловлена тремя D-мотивами, присутствующими на N-конце этих изоформ. [ 8 ]
Структура и функции
[ редактировать ]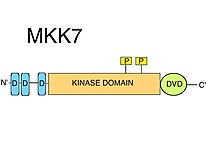
D-паттерны
[ редактировать ]MKK7 имеет три консервативных D-мотива (MAPK-рекрутирующие короткие линейные мотивы ) на своем внутренне неупорядоченном N-конце . D-мотивы обычно состоят из группы положительно заряженных аминокислот, за которыми следуют чередующиеся гидрофобные аминокислоты. [ 8 ] D-мотивы строго необходимы для рекрутирования субстратов MAPKK, таких как JNK. [ 10 ] Киназные домены MAPK содержат определенные поверхностные особенности, такие как так называемая область общего стыковки (CD), наряду с бороздкой стыковки (D), которые специфически распознают родственные им D-мотивы. [ 8 ] D-мотивы, обнаруженные в MKK7, высокоспецифичны для JNK, но имеют относительно низкую аффинность связывания. Было высказано предположение, что мотивы MKK7 могут синергизировать друг с другом, обеспечивая эффективное фосфорилирование субстрата. [ 11 ] Показано, что все три D-мотива необходимы для правильного образования комплекса JNK1:MKK7, а также для фосфорилирования и активации JNK1 с помощью МКК7. [ 12 ]
DVD-регион
[ редактировать ]Особое расширение ядра C-концевого киназного домена, так называемый «Домен универсального стыковки» (DVD), представляет собой область, обнаруженную в MKK7, как и в большинстве известных MAP2K. [ 10 ] Область DVD представляет собой стабильную, преимущественно спиральную складку примерно из 20 аминокислот, которая присоединяется к обратной стороне каталитического ядра киназных доменов MAP2K. [ 13 ] Это расширение домена требуется как для специфической привязки, так и для активации MKK7 соответствующими вышестоящими MAPKKK. Другим митоген-активируемым протеинкиназным киназам также требуется область DVD (в дополнение к различным другим неканоническим элементам их киназных доменов, таким как «петля MKK1/2»), чтобы иметь возможность различать различные вышестоящие MAPKKK . [ 14 ] Эти особые взаимодействия MAPKK:MAPKKK-киназа-домен/киназа-домен облегчают фосфорилирование MKK7. [ 8 ] Помимо активации MKK7, связывание с областью DVD может также влиять на петлю активации MKK7 таким образом, что Ser и Thr мотива S -KAK- T становятся доступными для фосфорилирования . [ 8 ]
Киназный домен
[ редактировать ]MKK7 содержит один киназный домен. Прямое взаимодействие MKK7:MAPKKK (с использованием области DVD) облегчает фосфорилирование MKK7 с помощью MAPKKK на серине и треонине в мотиве S -KAK- T в каталитическом домене (киназном домене). [ 9 ]
Сигнализация и регулирование
[ редактировать ]MKK7 играют важную роль в сигнальном пути активируемой стрессом протеинкиназы/c-Jun N-концевой киназы (SAP/JNK). [ 15 ] В сотрудничестве с другой митоген-активируемой протеинкиназной киназой MKK4 , MKK7 действуют как важные преобразователи выше передачи сигналов JNK. [ 16 ] Благодаря совместным усилиям два МКК фосфорилируют разные изоформы JNK . В результате МКК7 оказывает большое влияние на многочисленные физиологические процессы, такие как пролиферация и дифференцировка , а также на патологические процессы, такие как апоптоз и онкогенез . [ 9 ] МКК7 активируются в результате клеточных стрессов. [ 16 ] Они активируются рядом MKKK посредством фосфорилирования по мотиву S -KAK- T , расположенному в киназном домене MKK7s. МККК связаны с МКК7 через его сайт DVD на С-конце и фосфорилируют МКК7 по остаткам серина и треонина . [ 9 ] После активации MKK4 и MKK7 непосредственно фосфорилируют специфические остатки тирозина и треонина, расположенные в консервативном мотиве TPY петли активации белка JNK. [ 9 ] Хотя MKK7 действует посредством двойной специфичности, он имеет тенденцию фосфорилировать треонин в белке JNK, оставляя MKK4 фосфорилировать тирозин. [ 16 ] Фосфорилированные и активированные JNK активируют такие субстраты, как факторы транскрипции или проапоптотический белок. [ 9 ] MKK7 и MKK4, по-видимому, регулируют экспрессию друг друга, тем самым влияя на передачу сигналов JNK. Монофосфорилирование JNK по остатку треонина является адекватным для увеличения активности JNK, что свидетельствует о том, что МКК7 является важным компонентом активности JNK, тогда как дополнительное фосфорилирование остатка тирозина с помощью МКК4 обеспечивает более благоприятную активацию. [ 9 ] В целом MAP2K7 содержит несколько аминокислотных участков, которые фосфорилированы и убиквитинированы. [ 17 ]
Каркасные белки
[ редактировать ]
Помимо прямых взаимодействий между JNK, MKK7 и другими вышестоящими протеинкиназами, различные каркасные белки функционируют, обеспечивая специфичность между компонентами сигнального каскада MAPK. [ 8 ] [ 16 ] Различные изоформы JNK, MAPK и MAPKK (например, MKK7 или MKK4) специфически связываются с каркасными белками. Идентифицировано несколько каркасных белков млекопитающих. К ним относятся JNK-взаимодействующий белок (JIP) 1 и его более близкий гомолог JIP2 или (совершенно неродственные) белки JIP3 и JIP4. Тем не менее, было показано, что JIP1/2 и JIP3/4 способны напрямую взаимодействовать друг с другом. [ 19 ] Также было показано, что множество Src-гомологии-3 (POSH) является партнером JIP1/2. [ 16 ]
Все эти регуляторы пути JNK собирают транспортные комплексы, связанные с кинезин-зависимым везикулярным транспортом. В этом контексте JIP1/2 действуют как адаптеры груза, одновременно связываясь с моторным белком и белком-грузом. Помимо своих «обычных» грузов (С-концы трансмембранных белков) они также транспортируют ферменты MAP2K и MAP3K, а именно MKK7, DLK и MLK3. Киназы, связанные с каркасом JIP1/2, обычно секвестрируются и считаются неактивными. [ 18 ] Поскольку механизм связывания карго-связей этого комплекса считается зависимым от фосфорилирования, фосфорилирование с помощью киназы JNK может высвобождать свои собственные вышестоящие активаторы из каркаса, тем самым создавая сильную локальную петлю положительной обратной связи. [ 18 ] [ 20 ]
Взаимодействия
[ редактировать ]Было показано, что MAP2K7 взаимодействует с:
Биологическая значимость
[ редактировать ]МКК7 участвует в развитии эпителиальных тканей, таких как кожа и легкие, а также развивающихся зубов во время раннего эмбриогенеза у мышей. [ 8 ] Эксперименты также показывают, что MKK7 в дополнение к MKK4 необходимы для организации плана тела млекопитающих во время эмбриогенеза . [ 16 ] Также было высказано предположение, что MKK7 действует как предполагаемый ген-супрессор метастазов (MSG), возможно, способствуя состоянию покоя опухоли в месте метастазирования . [ 33 ] У мелких млекопитающих стресс, такой как перегрузка давлением, может вызвать сердечную гипертрофию и недостаточность, если MKK7 нокаутирован. [ 34 ] Условное удаление Map2k7 в нервных стволовых клетках и постмитотических нейронах выявило роль MKK7 в удлинении аксонов. [ 35 ] Нейрон-специфическая делеция Map2k7 показала роль MKK7 в возрастной двигательной дисфункции. [ 36 ] Генетические вариации MAP2K7 связаны с шизофренией у людей. [ 37 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000076984 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000002948 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Перейти обратно: а б «Ген Энтреза: митоген-активируемая протеинкиназная киназа 7 MAP2K7» .
- ^ Перейти обратно: а б Турнье С., Уитмарш А.Дж., Кавана Дж. и др. (февраль 1999 г.). «Ген MKK7 кодирует группу киназ c-Jun NH2-концевой киназы» . Молекулярная и клеточная биология . 19 (2): 1569–81. дои : 10.1128/mcb.19.2.1569 . ПМК 116085 . ПМИД 9891090 .
- ^ Фольц И.Н., Герл Р.Э., Вилер Дж.С. и др. (апрель 1998 г.). «Человеческая митоген-активируемая протеинкиназа-киназа 7 (MKK7) представляет собой высококонсервативную c-Jun N-концевую киназу/активируемую стрессом протеинкиназу (JNK/SAPK), активируемую стрессами окружающей среды и физиологическими стимулами» . Журнал биологической химии . 273 (15): 9344–51. дои : 10.1074/jbc.273.15.9344 . ПМИД 9535930 .
- ^ Перейти обратно: а б с д и ж г Ван X, Destrument A, Tournier C (август 2007 г.). «Физиологические роли MKK4 и MKK7: данные на животных моделях» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1773 (8): 1349–57. дои : 10.1016/j.bbamcr.2006.10.016 . ПМИД 17157936 .
- ^ Перейти обратно: а б с д и ж г Хаусген В., Хердеген Т., Ветциг В. (2011). «Узкое место передачи сигналов JNK: молекулярные и функциональные характеристики MKK4 и MKK7». Европейский журнал клеточной биологии . 90 (6–7): 536–44. дои : 10.1016/j.ejcb.2010.11.008 . ПМИД 21333379 .
- ^ Перейти обратно: а б Гантерт С., Хонеркамп Дж., Тиммер Дж. (1992). «Анализ динамики временных рядов тремора рук». Биологическая кибернетика . 66 (6): 479–84. дои : 10.1007/bf00204112 . ПМИД 1586672 . S2CID 22250412 .
- ^ Хо Д.Т., Бардуэлл А.Дж., Абдоллахи М. и др. (август 2003 г.). «Сайт стыковки в MKK4 обеспечивает высокое сродство связывания с JNK MAPK и конкурирует с аналогичными сайтами стыковки в субстратах JNK» . Журнал биологической химии . 278 (35): 32662–72. дои : 10.1074/jbc.M304229200 . ПМК 3017503 . ПМИД 12788955 .
- ^ Хо Д.Т., Бардуэлл А.Дж., Гревал С. и др. (май 2006 г.). «Взаимодействующие сайты стыковки JNK в MKK7 способствуют связыванию и активации митоген-активируемых протеинкиназ JNK» . Журнал биологической химии . 281 (19): 13169–79. дои : 10.1074/jbc.M601010200 . ПМК 3017509 . ПМИД 16533805 .
- ^ Раман М., Чен В., Кобб М.Х. (май 2007 г.). «Дифференциальное регулирование и свойства МАПК» . Онкоген . 26 (22): 3100–12. дои : 10.1038/sj.onc.1210392 . ПМИД 17496909 .
- ^ Ременьи А., Good MC, Лим В.А. (декабрь 2006 г.). «Стыковочные взаимодействия в сетях протеинкиназ и фосфатаз». Современное мнение в области структурной биологии . 16 (6): 676–85. дои : 10.1016/j.sbi.2006.10.008 . ПМИД 17079133 .
- ^ Яо З., Динер К., Ван К.С. и др. (декабрь 1997 г.). «Активация стресс-активируемых протеинкиназ/N-концевых протеинкиназ c-Jun (SAPK/JNK) новой митоген-активируемой протеинкиназной киназой» . Журнал биологической химии . 272 (51): 32378–83. дои : 10.1074/jbc.272.51.32378 . ПМИД 9405446 .
- ^ Перейти обратно: а б с д и ж г Асаока Ю., Нишина Х (октябрь 2010 г.). «Разнообразные физиологические функции МКК4 и МКК7 во время раннего эмбриогенеза» . Журнал биохимии . 148 (4): 393–401. дои : 10.1093/jb/mvq098 . ПМИД 20801953 .
- ^ «МКК7 (человек)» . www.фосфосайт.орг . Проверено 28 октября 2020 г.
- ^ Перейти обратно: а б с Нихалани Д., Вонг Х.Н., Хольцман Л.Б. (август 2003 г.). «Привлечение JNK к JIP1 и JNK-зависимое фосфорилирование JIP1 регулирует динамику и активацию модуля JNK» . Журнал биологической химии . 278 (31): 28694–702. дои : 10.1074/jbc.M304212200 . ПМИД 12756254 .
- ^ Хаммонд Дж.В., Гриффин К., Джих Г.Т. и др. (май 2008 г.). «Кооперативная и независимая транспортировка различных грузов Кинесином-1». Трафик . 9 (5): 725–41. дои : 10.1111/j.1600-0854.2008.00722.x . hdl : 2027.42/72137 . ПМИД 18266909 . S2CID 21901129 .
- ^ Нихалани Д., Вонг Х., Верма Р. и др. (апрель 2007 г.). «Киназы семейства Src напрямую регулируют динамику и активацию модуля JIP1» . Молекулярная и клеточная биология . 27 (7): 2431–41. дои : 10.1128/MCB.01479-06 . ПМК 1899903 . ПМИД 17242197 .
- ^ Яффе А.Б., зал А, Шмидт А. (март 2005 г.). «Ассоциация CNK1 с факторами обмена гуаниновых нуклеотидов Rho контролирует специфичность передачи сигналов ниже Rho» . Современная биология . 15 (5): 405–12. Бибкод : 2005CBio...15..405J . дои : 10.1016/j.cub.2004.12.082 . ПМИД 15753034 . S2CID 16479940 .
- ^ Карандикар М., Сюй С., Кобб М.Х. (декабрь 2000 г.). «MEKK1 связывает raf-1 и компоненты каскада ERK2» . Журнал биологической химии . 275 (51): 40120–7. дои : 10.1074/jbc.M005926200 . ПМИД 10969079 .
- ^ Турнье С., Уитмарш А.Дж., Кавана Дж. и др. (июль 1997 г.). «Митоген-активируемая протеинкиназа-киназа 7 является активатором NH2-концевой киназы c-Jun» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (14): 7337–42. Бибкод : 1997PNAS...94.7337T . дои : 10.1073/pnas.94.14.7337 . ПМК 23822 . ПМИД 9207092 .
- ^ Перейти обратно: а б Ченг Дж., Ян Дж., Ся Ю. и др. (апрель 2000 г.). «Синергическое взаимодействие киназы 2 MEK, N-концевой киназы 2 c-Jun (JNK) и JNK1 приводит к эффективной и специфической активации JNK1» . Молекулярная и клеточная биология . 20 (7): 2334–42. дои : 10.1128/MCB.20.7.2334-2342.2000 . ПМЦ 85399 . ПМИД 10713157 .
- ^ Келкар Н., Гупта С., Диккенс М. и др. (февраль 2000 г.). «Взаимодействие митоген-активируемого сигнального модуля протеинкиназы с нейрональным белком JIP3» . Молекулярная и клеточная биология . 20 (3): 1030–43. дои : 10.1128/MCB.20.3.1030-1043.2000 . ПМЦ 85220 . ПМИД 10629060 .
- ^ Мацуура Х., Нисито Х., Такеда К. и др. (октябрь 2002 г.). «Зависимая от фосфорилирования поддерживающая роль JSAP1/JIP3 в сигнальном пути ASK1-JNK. Новый способ регуляции киназного каскада MAP» . Журнал биологической химии . 277 (43): 40703–9. дои : 10.1074/jbc.M202004200 . hdl : 2297/2692 . ПМИД 12189133 .
- ^ Перейти обратно: а б Ясуда Дж., Уитмарш А.Дж., Кавана Дж. и др. (октябрь 1999 г.). «Группа JIP митоген-активируемых каркасных белков протеинкиназ» . Молекулярная и клеточная биология . 19 (10): 7245–54. дои : 10.1128/mcb.19.10.7245 . ПМЦ 84717 . ПМИД 10490659 .
- ^ Папа С., Заззерони Ф., Бубичи С. и др. (февраль 2004 г.). «Gadd45 бета опосредует подавление передачи сигналов JNK NF-каппа B путем нацеливания на MKK7/JNKK2». Природная клеточная биология . 6 (2): 146–53. дои : 10.1038/ncb1093 . ПМИД 14743220 . S2CID 5250125 .
- ^ Мерритт С.Е., Мата М., Нихалани Д. и др. (апрель 1999 г.). «Киназа DLK смешанного происхождения использует в качестве субстрата MKK7, а не MKK4» . Журнал биологической химии . 274 (15): 10195–202. дои : 10.1074/jbc.274.15.10195 . ПМИД 10187804 .
- ^ Негри С., Оберсон А., Штайнманн М. и др. (март 2000 г.). «Клонирование кДНК и картирование нового белка, взаимодействующего с островком мозга / JNK». Геномика . 64 (3): 324–30. дои : 10.1006/geno.2000.6129 . ПМИД 10756100 .
- ^ Зама Т., Аоки Р., Камимото Т. и др. (июнь 2002 г.). «Роль каркаса митоген-активируемой протеинкиназы-фосфатазы, SKRP1, для сигнального пути JNK» (PDF) . Журнал биологической химии . 277 (26): 23919–26. дои : 10.1074/jbc.M200838200 . ПМИД 11959862 . S2CID 12430487 .
- ^ Зама Т., Аоки Р., Камимото Т. и др. (июнь 2002 г.). «Новая фосфатаза двойной специфичности SKRP1 взаимодействует с киназой MAPK MKK7 и инактивирует путь JNK MAPK. Значение для точной регуляции конкретного пути MAPK» . Журнал биологической химии . 277 (26): 23909–18. дои : 10.1074/jbc.M200837200 . ПМИД 11959861 .
- ^ Наумов Г.Н., Фолкман Дж., Страуме О. и др. (2008). «Опухолево-сосудистые взаимодействия и покой опухоли» . АПМИС . 116 (7–8): 569–85. дои : 10.1111/j.1600-0463.2008.01213.x . ПМЦ 3508681 . ПМИД 18834403 .
- ^ Лю В., Цзы М., Чи Х. и др. (апрель 2011 г.). «Лишение МКК7 в кардиомиоцитах провоцирует сердечную недостаточность у мышей при воздействии перегрузки давлением». Журнал молекулярной и клеточной кардиологии . 50 (4): 702–11. дои : 10.1016/j.yjmcc.2011.01.013 . ПМИД 21284947 .
- ^ Ямасаки Т., Кавасаки Х., Аракава С. и др. (ноябрь 2011 г.). «Стресс-активируемая протеинкиназа MKK7 регулирует удлинение аксонов в развивающейся коре головного мозга» . Журнал неврологии . 31 (46): 16872–83. doi : 10.1523/JNEUROSCI.1111-11.2011 . ПМК 6633308 . ПМИД 22090513 .
- ^ Ямасаки Т., Деки-Арима Н., Канеко А. и др. (август 2017 г.). «Возрастная двигательная дисфункция из-за нейронспецифического нарушения стресс-активируемой протеинкиназы MKK7» . Научные отчеты . 7 (1): 7348. Бибкод : 2017НатСР...7.7348Y . дои : 10.1038/s41598-017-07845-x . ПМК 5544763 . ПМИД 28779160 .
- ^ Винчестер К.Л., Озеки Х., Вуюуклис Д.А. и др. (ноябрь 2012 г.). «Сходящиеся доказательства того, что вариации последовательности нового гена-кандидата MAP2K7 (MKK7) функционально связаны с шизофренией» . Молекулярная генетика человека . 21 (22): 4910–21. дои : 10.1093/hmg/dds331 . ПМИД 22899651 .
Дальнейшее чтение
[ редактировать ]- Лин А (2006). «Сигнальный путь JNK (отдел молекулярной биологии)». Ландес Бионауки . 1 :1–97. ISBN 978-1-58706-120-2 .