Ви1
| Ви1 | |||
|---|---|---|---|
 Кристаллическая структура человеческого Wee1 | |||
| Идентификаторы | |||
| Символ | Ингибитор митоза протеинкиназа Wee1 | ||
| Альт. символы | протеинкиназа двойной специфичности wee1 Wee1 | ||
| ген NCBI | 2539123 | ||
| ЮниПрот | P07527 | ||
| Другие данные | |||
| Номер ЕС | 2.7.11.1 | ||
| |||
Wee1 представляет собой ядерную киназу, принадлежащую к семейству протеинкиназ Ser/Thr делящихся дрожжей Schizosaccharomyces pombe ( S. pombe ). Wee1 имеет молекулярную массу 96 кДа и является ключевым регулятором клеточного цикла . Он влияет на размер клеток, ингибируя вступление в митоз посредством ингибирования Cdk1 . Wee1 имеет гомологи у многих других организмов, включая млекопитающих.
Введение
[ редактировать ]Регулирование размера клеток имеет решающее значение для обеспечения функциональности клетки. Помимо факторов окружающей среды, таких как питательные вещества, факторы роста и функциональная нагрузка, размер клеток также контролируется контрольной точкой размера клеток.
Wee1 является компонентом этой контрольной точки. Это киназа, определяющая момент вступления в митоз, тем самым влияя на размер дочерних клеток. Потеря функции Wee1 приведет к образованию дочерних клеток меньшего размера, чем нормальные, поскольку деление клеток происходит преждевременно.
Его название происходит от шотландского диалектного слова «wee», что означает «маленький» — его первооткрыватель Пол Нерс во время открытия работал в Эдинбургском университете в Шотландии. [1] [2]
Функция
[ редактировать ]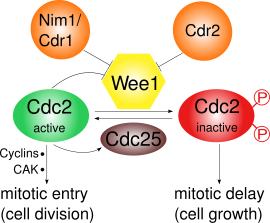
Wee1 ингибирует Cdk1 , фосфорилируя его в двух разных сайтах: Tyr15 и Thr14. [3] Cdk1 имеет решающее значение для циклин-зависимого прохождения различных контрольных точек клеточного цикла.Существует как минимум три контрольных точки, для которых важно ингибирование Cdk1 с помощью Wee1:
- Контрольная точка G2/M : Wee1 фосфорилирует аминокислоты Tyr15 и Thr14 Cdk1, что поддерживает низкую киназную активность Cdk1 и предотвращает вступление в митоз ; у S. pombe может происходить дальнейший рост клеток. Было показано, что опосредованная Wee1 инактивация Cdk1 является сверхчувствительной в результате конкуренции субстратов. [4] Во время вступления в митоз активность Wee1 снижается несколькими регуляторами и, таким образом, активность Cdk1 увеличивается. У S. pombe протеинкиназа . Pom1 локализуется на полюсах клетки Это активирует путь, по которому Cdr2 ингибирует Wee1 через Cdr1. Сам Cdk1 отрицательно регулирует Wee1 посредством фосфорилирования, что приводит к возникновению петли положительной обратной связи. Снижение активности Wee1 само по себе недостаточно для входа в митоз: также необходимы синтез циклинов и активирующее фосфорилирование с помощью киназы, активирующей Cdk (CAK). [5]
- Контрольная точка размера клеток: существуют доказательства существования контрольной точки размера клеток, которая предотвращает вступление мелких клеток в митоз. Wee1 играет роль в этой контрольной точке, координируя размер клеток и развитие клеточного цикла. [6]
- Контрольная точка повреждения ДНК: эта контрольная точка также контролирует переход G2/M. У S. pombe эта контрольная точка задерживает вступление в митоз клеток с повреждением ДНК (например, вызванным гамма-излучением ). Удлинение фазы G2 зависит от Wee1; Мутанты wee1 не имеют продленной фазы G2 после гамма-облучения. [7]
Также сообщалось об эпигенетической функции киназы Wee1. Было показано, что Wee1 фосфорилирует гистон H2B по остатку тирозина 37, который регулирует глобальную экспрессию гистонов. [8] [9]
Гомологи
[ редактировать ]| человеческий гомолог WEE1 ( S. pombe ) | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | WEE1 | ||
| ген NCBI | 7465 | ||
| HGNC | 12761 | ||
| МОЙ БОГ | 193525 | ||
| RefSeq | НМ_003390 | ||
| ЮниПрот | P30291 | ||
| Другие данные | |||
| Локус | Хр. 11 п15.3-15.1 | ||
| |||
| гомолог 2 человеческого WEE1 ( S. pombe ) | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ОЭЭ2 | ||
| ген NCBI | 494551 | ||
| HGNC | 19684 | ||
| RefSeq | НМ_001105558 | ||
| ЮниПрот | P0C1S8 | ||
| Другие данные | |||
| Локус | Хр. 7 q32-q32 | ||
| |||
WEE1 Ген имеет два известных гомолога у человека: WEE1 (также известный как WEE1A) и WEE2 (WEE1B). Соответствующими белками являются Wee1-подобная протеинкиназа и Wee1-подобная протеинкиназа 2, которые действуют на гомолог Cdk1 человека Cdk1 .
Гомолог Wee1 у почкующихся дрожжей Saccharomyces cerevisiae называется Swe1.
Регулирование
[ редактировать ]У S. pombe Wee1 фосфорилирован.
Cdk1 и циклин B составляют фактор, способствующий созреванию (MPF), который способствует вступлению в митоз. Он инактивируется фосфорилированием через Wee1 и активируется фосфатазой Cdc25C . Cdc25C, в свою очередь, активируется киназой Polo и инактивируется Chk1 . [6] Таким образом, у S. pombe регуляция Wee1 в основном находится под контролем фосфорилирования через путь киназы полярности Pom1 , включающий Cdr2 и Cdr1. [10] [11] [12] [13]
При переходе G2/M Cdk1 активируется Cdc25 посредством дефосфорилирования Tyr15. В то же время Wee1 инактивируется посредством фосфорилирования его С-концевого каталитического домена с помощью Nim1/Cdr1. [12] Кроме того, активный MPF будет способствовать своей собственной активности, активируя Cdc25 и инактивируя Wee1, создавая петлю положительной обратной связи , хотя это еще не изучено подробно. [6]
Высшие эукариоты регулируют Wee1 посредством фосфорилирования и деградации.
У высших эукариот инактивация Wee1 происходит как за счет фосфорилирования, так и за счет деградации . [14] Белковый комплекс [номер 1] СКФ β-TrCP1/2 E3 представляет собой убиквитинлигазу , которая участвует в убиквитинировании Wee1A . М -фазы Киназы Polo-подобная киназа (Plk1) и Cdc2 фосфорилируют два сериновых остатка в Wee1A, которые распознаются SCF. β-TrCP1/2 . [15]
Гомолог S. cerevisiae Swe1
У S. cerevisiae циклинзависимая киназа Cdc28 (гомолог Cdk1) фосфорилируется Swe1 (гомолог Wee1) и дефосфорилируется Mih1 (гомолог Cdc25). Гомолог Nim1/Cdr1 в S. cerevisiae , Hsl1, вместе с родственными киназами Gin4 и Kcc4 локализуют Swe1 на шейке почки . Киназы Cla4 и Cdc5, ассоциированные с шейкой почки (гомолог полокиназы), фосфорилируют Swe1 на разных стадиях клеточного цикла. Swe1 также фосфорилируется с помощью Clb2-Cdc28, что служит признанием дальнейшего фосфорилирования с помощью Cdc5.
Белок S. cerevisiae Swe1 также регулируется путем деградации. Swe1 гиперфосфорилируется с помощью Clb2-Cdc28 и Cdc5, что может быть сигналом для убиквитинирования и деградации с помощью SCF E3, как у высших эукариот. убиквитинлигазного комплекса [16]
Роль в раке
[ редактировать ]Фактор, способствующий митозу MPF, также регулирует повреждением ДНК , вызванный апоптоз . Негативная регуляция MPF с помощью WEE1 вызывает аберрантный митоз и, следовательно, устойчивость к апоптозу, вызванному повреждением ДНК. Круппель-подобный фактор 2 (KLF2) отрицательно регулирует человеческий WEE1, тем самым повышая чувствительность к апоптозу, вызванному повреждением ДНК, в раковых клетках. [17]
Мутантный фенотип
[ редактировать ]Wee1 действует как дозозависимый ингибитор митоза. [18] Таким образом, количество белка Wee1 коррелирует с размером клеток:
делящихся дрожжей Мутант wee1 , также называемый wee1. − , делится при значительно меньшем размере клеток, чем клетки дикого типа. Поскольку Wee1 ингибирует вступление в митоз, его отсутствие приведет к преждевременному делению и субнормальному размеру клеток. Наоборот,когда экспрессия Wee1 увеличивается, митоз задерживается, и клетки вырастают до больших размеров перед делением.
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Белок 1/2, содержащий повторы β-трансдуцина (β-TrCP1/2) Белковый комплекс F-box, содержащий SKP1/Cul1/F-box
Ссылки
[ редактировать ]- ^ Медсестра П. (декабрь 2004 г.). «Маленькие зверюшки» . Природа . 432 (7017): 557. Бибкод : 2004Natur.432..557N . дои : 10.1038/432557a . ПМИД 15577889 . S2CID 29840746 .
- ^ Медсестра П., Тюрио П. (ноябрь 1980 г.). «Регуляторные гены, контролирующие митоз у делящихся дрожжей Schizosaccharomyces pombe» . Генетика . 96 (3): 627–37. дои : 10.1093/генетика/96.3.627 . ПМЦ 1214365 . ПМИД 7262540 .
- ^ Ден Хаезе Г.Дж., Уолворт Н., Карр А.М., Гулд К.Л. (1995). «Протеинкиназа Wee1 регулирует фосфорилирование T14 делящихся дрожжей Cdc2» . Мол Биол Клетка . 6 (4): 371–85. дои : 10.1091/mbc.6.4.371 . ПМК 301198 . ПМИД 7626804 .
- ^ Ким С.Ю., Феррелл Дж.Э.Дж. (23 марта 2007 г.). «Субстратная конкуренция как источник сверхчувствительности при инактивации Wee1» . Клетка . 128 (6): 1133–45. дои : 10.1016/j.cell.2007.01.039 . ПМИД 17382882 . S2CID 14138576 .
- ^ Коулман Т.Р., Данфи В.Г. (1994). «Факторы регуляции Cdc2». Современное мнение в области клеточной биологии . 6 (6): 877–82. дои : 10.1016/0955-0674(94)90060-4 . ПМИД 7880537 .
- ^ Jump up to: а б с Келлог Д.Р. (2003). «Wee1-зависимые механизмы, необходимые для координации роста и деления клеток». J Cell Sci . 116 (24): 4883–90. дои : 10.1242/jcs.00908 . ПМИД 14625382 . S2CID 14743149 .
- ^ Роули Р., Хадсон Дж., Янг П.Г. (1992). «Протеинкиназа wee1 необходима для радиационно-индуцированной задержки митоза». Природа . 356 (6367): 353–5. Бибкод : 1992Natur.356..353R . дои : 10.1038/356353a0 . ПМИД 1549179 . S2CID 4280074 .
- ^ Махаджан К., Фанг Б., Кумен Дж. М., Махаджан Н. П. (2012). «Фосфорилирование H2B Tyr37 подавляет экспрессию зависимых от репликации основных генов гистонов» . Структурная и молекулярная биология природы . 19 (9): 930–7. дои : 10.1038/nsmb.2356 . ПМЦ 4533924 . ПМИД 22885324 .
- ^ Махаджан К., Махаджан Н.П. (2013). «Тирозинкиназа WEE1, новый эпигенетический модификатор» . Тенденции Жене . 29 (7): 394–402. дои : 10.1016/j.tig.2013.02.003 . ПМК 3700603 . ПМИД 23537585 .
- ^ Бодди М.Н., Фурнари Б., Мондесерт О., Рассел П. (май 1998 г.). «Контрольная точка репликации, обеспечиваемая киназами Cds1 и Chk1». Наука . 280 (5365): 909–12. Бибкод : 1998Sci...280..909B . дои : 10.1126/science.280.5365.909 . ПМИД 9572736 .
- ^ Ву Л, Рассел П. (июнь 1993 г.). «Киназа Nim1 способствует митозу, инактивируя тирозинкиназу Wee1». Природа . 363 (6431): 738–41. Бибкод : 1993Natur.363..738W . дои : 10.1038/363738a0 . ПМИД 8515818 . S2CID 4320080 .
- ^ Jump up to: а б Коулман Т.Р., Тан З., Данфи В.Г. (март 1993 г.). «Негативная регуляция протеинкиназы wee1 путем прямого действия митотического индуктора nim1/cdr1». Клетка . 72 (6): 919–29. дои : 10.1016/0092-8674(93)90580-J . ПМИД 7681363 . S2CID 42256641 .
- ^ Тан З., Коулман Т.Р., Данфи В.Г. (сентябрь 1993 г.). «Два различных механизма негативной регуляции протеинкиназы Wee1» . ЭМБО Дж . 12 (9): 3427–36. дои : 10.1002/j.1460-2075.1993.tb06017.x . ПМК 413619 . ПМИД 7504624 .
- ^ Ватанабэ Н., Брум М., Хантер Т. (май 1995 г.). «Регуляция тирозин-15-киназы WEE1Hu CDK человека во время клеточного цикла» . ЭМБО Дж . 14 (9): 1878–91. дои : 10.1002/j.1460-2075.1995.tb07180.x . ПМЦ 398287 . ПМИД 7743995 .
- ^ Ватанабэ Н., Арай Х., Нисихара Ю. и др. (март 2004 г.). «Киназы М-фазы индуцируют фосфозависимое убиквитинирование соматического Wee1 с помощью SCFbeta-TrCP» . Учеб. Натл. акад. наук. США . 101 (13): 4419–24. Бибкод : 2004PNAS..101.4419W . дои : 10.1073/pnas.0307700101 . ПМЦ 384762 . ПМИД 15070733 .
- ^ Ли К.С., Асано С., Пак Дж.Э., Сакчайсри К., Эриксон Р.Л. (октябрь 2005 г.). «Мониторинг клеточного цикла посредством мультикиназно-зависимой регуляции Swe1/Wee1 у почкующихся дрожжей» . Клеточный цикл . 4 (10): 1346–9. дои : 10.4161/cc.4.10.2049 . ПМИД 16123596 .
- ^ Ван Ф., Чжу Ю, Хуан Ю и др. (июнь 2005 г.). «Транкрипционная репрессия WEE1 с помощью Круппелеподобного фактора 2 участвует в апоптозе, вызванном повреждением ДНК» . Онкоген . 24 (24): 3875–85. дои : 10.1038/sj.onc.1208546 . ПМИД 15735666 .
- ^ Рассел П., медсестра П. (май 1987 г.). «Негативная регуляция митоза с помощью wee1+, гена, кодирующего гомолог протеинкиназы». Клетка . 49 (4): 559–67. дои : 10.1016/0092-8674(87)90458-2 . ПМИД 3032459 . S2CID 42801276 .