ГСК-3
| Киназа 3 гликогенсинтазы, каталитический домен | |
|---|---|
| Идентификаторы | |
| Символ | STKc_GSK3 |
| ИнтерПро | ИПР039192 |
| CDD | cd14137 |
| гликогенсинтаза киназа 3 альфа | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ГСК3А | ||
| ген NCBI | 2931 | ||
| HGNC | 4616 | ||
| МОЙ БОГ | 606784 | ||
| RefSeq | НМ_019884 | ||
| ЮниПрот | P49840 | ||
| Другие данные | |||
| Номер ЕС | 2.7.11.26 | ||
| Локус | Хр. 19 q13.2 | ||
| |||
| гликогенсинтаза киназа 3 бета | |||
|---|---|---|---|
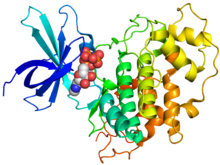 Кристаллографическая структура человеческого GSK-3β (радужного цвета, N-конец = синий, C-конец = красный), связанного с аденилатным эфиром фосфоаминофосфоновой кислоты (сферы). [1] | |||
| Идентификаторы | |||
| Символ | ГСК3Б | ||
| ген NCBI | 2932 | ||
| HGNC | 4617 | ||
| МОЙ БОГ | 605004 | ||
| ПДБ | 1Q3W Больше структур | ||
| RefSeq | НМ_002093 | ||
| ЮниПрот | P49841 | ||
| Другие данные | |||
| Номер ЕС | 2.7.11.26 | ||
| Локус | 3 q13.33 | ||
| |||
Киназа гликогенсинтазы 3 ( GSK-3 ) представляет собой серин/треониновую протеинкиназу , которая опосредует добавление молекул фосфата к серина и треонина аминокислотным остаткам . Впервые обнаружена в 1980 году как регуляторная киназа своей тезки, гликогенсинтазы (GS). [2] С тех пор GSK-3 была идентифицирована как протеинкиназа для более чем 100 различных белков в различных путях. [3] [4] У млекопитающих, включая человека, GSK-3 существует в виде двух изоферментов , кодируемых двумя гомологичными генами GSK-3α ( GSK3A ) и GSK-3β ( GSK3B ). GSK-3 был предметом многочисленных исследований, поскольку он участвует в ряде заболеваний, включая диабет 2 типа , болезнь Альцгеймера , воспаления , рак , зависимость. [5] и биполярное расстройство .
GSK-3 представляет собой серин/треониновую протеинкиназу, которая фосфорилирует либо треонин , либо серин , и это фосфорилирование контролирует различные биологические активности, такие как метаболизм гликогена , передача сигналов в клетках , клеточный транспорт и другие. [6] Ингибирование GS с помощью GSK-3β приводит к снижению синтеза гликогена в печени и мышцах наряду с повышением уровня глюкозы в крови или гипергликемией. [7] Вот почему GSK-3β связан с патогенезом и прогрессированием многих заболеваний, таких как диабет , ожирение , рак , [8] и болезнь Альцгеймера. [9] Он активен в покоящихся клетках и ингибируется несколькими гормонами, такими как инсулин , фактор роста эндотелия и фактор роста тромбоцитов . Инсулин косвенно инактивирует GSK3 посредством нижележащего фосфорилирования специфических остатков серина Ser21 и Ser9 в изоформах GSK-3 α и β соответственно через путь PI3K/Akt . [10] [11]
По состоянию на 2019 год [update]GSK-3 является единственным названным и признанным типом киназы гликогенсинтазы . Символы генов GSK1 и GSK2 были отозваны Комитетом по номенклатуре генов HUGO (HGNC), и не было указано ни новых названий этих «генов», ни их местонахождения. [12] [13]
Механизм
[ редактировать ]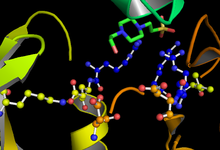
GSK-3 функционирует путем фосфорилирования остатка серина или треонина на целевом субстрате. Положительно заряженный карман, прилегающий к активному сайту, связывает «праймирующую» фосфатную группу, прикрепленную к четырем остаткам серина или треонина, С-концевым целевого сайта фосфорилирования. Активный центр в остатках 181, 200, 97 и 85 связывает концевой фосфат АТФ и переносит его в целевое место на субстрате (см. рисунок 1). [14]
Гликогенсинтаза
[ редактировать ]Гликогенсинтаза — фермент , отвечающий за синтез гликогена . Он активируется глюкозо-6-фосфатом (G6P) и ингибируется киназой гликогенсинтазы ( GSK3 ). Эти два механизма играют важную роль в метаболизме гликогена. [15]
Функция
[ редактировать ]Фосфорилирование белка с помощью GSK-3 обычно ингибирует активность его нижестоящей мишени. [16] [17] [18] GSK-3 активен в ряде центральных внутриклеточных сигнальных путей, включая клеточную пролиферацию, миграцию, регуляцию глюкозы и апоптоз.
GSK-3 был первоначально обнаружен в связи с его участием в регуляции гликогенсинтазы . [2] После праймирования казеинкиназой 2 (СК2) гликогенсинтаза фосфорилируется по кластеру из трех С-концевых остатков серина, снижая ее активность. [19] Помимо своей роли в регуляции гликогенсинтазы, GSK-3 участвует в других аспектах гомеостаза глюкозы, включая фосфорилирование инсулинового рецептора IRS1. [20] и глюконеогенных ферментов фосфоенолпируваткарбоксикиназы и глюкозо-6-фосфатазы . [21] Однако эти взаимодействия не были подтверждены, поскольку эти пути могут быть ингибированы без усиления регуляции GSK-3. [19]
Также было показано, что GSK-3 регулирует иммунные и миграционные процессы. GSK-3 участвует в ряде сигнальных путей врожденного иммунного ответа, включая выработку провоспалительных цитокинов и интерлейкинов. [22] [23] Инактивация GSK3B различными протеинкиназами также влияет на адаптивный иммунный ответ, индуцируя выработку и пролиферацию цитокинов в наивных CD4+ Т-клетках и клетках памяти. [23] Сообщалось, что в клеточной миграции, неотъемлемом аспекте воспалительных реакций, ингибирование GSK-3 играет противоречивую роль, поскольку было показано, что локальное ингибирование в конусах роста способствует подвижности, в то время как было показано, что глобальное ингибирование клеточного GSK-3 подавляет Распространение и миграция клеток. [22]
GSK-3 также неразрывно связан с путями клеточной пролиферации и апоптоза. Было показано, что GSK-3 фосфорилирует бета-катенин , тем самым направляя его на деградацию. [24] Таким образом, GSK-3 является частью канонического пути бета-катенин / Wnt , который сигнализирует клетке о необходимости деления и пролиферации. GSK-3 фосфорилирует циклины D и E, важные для перехода из фазы G1 в S-фазу, и вызывает их деградацию. Факторы транскрипции c-myc и c-fos (также промоторы S-фазы), которые в первую очередь фосфорилируются киназой, регулируемой фосфорилированием тирозина двойной специфичности, также фосфорилируются с помощью GSK3, вызывая их деградацию. [25] GSK-3 также участвует в ряде апоптотических сигнальных путей путем фосфорилирования факторов транскрипции, которые регулируют апоптоз . [4] GSK-3 может стимулировать апоптоз, активируя проапоптотические факторы, такие как р53. [26] и инактивация факторов, способствующих выживанию, посредством фосфорилирования. [27] Однако роль GSK-3 в регуляции апоптоза противоречива, поскольку некоторые исследования показали, что мыши с нокаутом GSK-3β чрезмерно чувствительны к апоптозу и умирают на эмбриональной стадии, в то время как другие показали, что сверхэкспрессия GSK-3 может индуцировать апоптоз. . [28] В целом, GSK-3, по-видимому, одновременно способствует и ингибирует апоптоз, и эта регуляция варьируется в зависимости от конкретного молекулярного и клеточного контекста. [29]
GSK-3 также участвует в сигнальном пути ядерного активатора транскрипции каппа B (NFκB), сигнальном пути Hedgehog, сигнальном пути Notch и эпителиально-мезенхимальном переходе. [25]
Регулирование
[ редактировать ]Из-за своей важности для многочисленных клеточных функций активность GSK-3 подлежит жесткому регулированию.
Скорость и эффективность фосфорилирования GSK-3 регулируется несколькими факторами. Фосфорилирование определенных остатков GSK-3 может увеличивать или уменьшать его способность связывать субстрат. Фосфорилирование тирозина-216 в GSK-3β или тирозина-279 в GSK-3α усиливает ферментативную активность GSK-3, тогда как фосфорилирование аутоингибирующего серина-9 в GSK-3β или серина-21 в GSK-3α значительно снижает доступность активного центра. (см. рисунок). [22] Кроме того, GSK-3 необычен среди киназ тем, что для первого фосфорилирования субстрата обычно требуется «праймирующая киназа». Фосфорилированный остаток серина или треонина, расположенный на С-конце четырех аминокислот по отношению к целевому сайту фосфорилирования, позволяет субстрату связывать карман положительного заряда, образованный остатками аргинина и лизина. [19] [30]
В зависимости от пути использования GSK-3 может дополнительно регулироваться за счет клеточной локализации или образования белковых комплексов. Активность GSK-3 гораздо выше в ядре и митохондриях, чем в цитозоле корковых нейронов. [31] в то время как фосфорилирование бета-катенина с помощью GSK-3 опосредовано связыванием обоих белков с аксином , каркасным белком, что позволяет бета-катенину получить доступ к активному сайту GSK-3. [22]
Инсулин косвенно инактивирует GSK3 посредством нижележащего фосфорилирования специфических остатков серина Ser21 и Ser9 в изоформах GSK-3 α и β соответственно через путь PI3K/Akt (протеинкиназа B). [10] [11]
Актуальность заболевания
[ редактировать ]Из-за своего участия в большом количестве сигнальных путей GSK-3 связан с множеством громких заболеваний. Ингибиторы GSK-3 в настоящее время тестируются на предмет терапевтического действия при болезни Альцгеймера , сахарном диабете 2 типа (СД2), некоторых формах рака и биполярном расстройстве . [32]
Есть доказательства того, что литий , который используется для лечения биполярного расстройства , действует как стабилизатор настроения, избирательно ингибируя GSK-3. Механизм, посредством которого ингибирование GSK-3 может стабилизировать настроение, неизвестен, хотя есть подозрения, что ингибирование способности GSK-3 способствовать воспалению способствует терапевтическому эффекту. [22] Ингибирование GSK-3 также дестабилизирует альфа-репрессор транскрипции Rev-ErbA, который играет важную роль в циркадных часах. [33] Элементы циркадных часов могут быть связаны с предрасположенностью к биполярному расстройству настроения. [34]
Активность GSK-3 связана как с патологическими особенностями болезни Альцгеймера, а именно с накоплением отложений амилоида-β (Aβ), так и с образованием нейрофибриллярных клубков . Считается, что GSK-3 напрямую способствует производству Aβ и связан с процессом гиперфосфорилирования тау -белков , что приводит к образованию клубков. [4] [22] Благодаря роли GSK-3 в развитии болезни Альцгеймера, ингибиторы GSK-3 могут оказывать положительное терапевтическое воздействие на пациентов с болезнью Альцгеймера и в настоящее время находятся на ранних стадиях испытаний. [35]
Аналогичным образом целенаправленное ингибирование GSK-3 может иметь терапевтический эффект при некоторых видах рака. Хотя было показано, что GSK-3 в некоторых случаях способствует апоптозу , также сообщается, что он является ключевым фактором онкогенеза при некоторых видах рака. [36] В подтверждение этого утверждения было показано, что ингибиторы GSK-3 индуцируют апоптоз в клетках глиомы и рака поджелудочной железы. [28] [37] GSK-3 также, по-видимому, ответственен за аберрантную активность NFκB при остром лимфобластном лейкозе у детей и клетках рака поджелудочной железы. В клетках рака почки ингибиторы GSK-3 вызывают остановку клеточного цикла, дифференцировку злокачественных клеток и аутофагию. В отличие от вышеперечисленных новообразований, высокая экспрессия неактивного pGSK3β-S9 обнаруживается при раке кожи, полости рта и легких, что позволяет предположить опухолесупрессивное действие фермента при этих видах рака. При меланоме микроРНК миР-769 ингибирует активность GSK-3 в процессе развития опухоли, что также указывает на опухолесупрессивное действие GSK3. [25]
Ингибиторы GSK-3 также оказались многообещающими в лечении СД2. [19] Хотя активность GSK-3 при диабетических состояниях может радикально различаться в разных типах тканей, исследования показали, что введение конкурентных ингибиторов GSK-3 может повысить толерантность к глюкозе у мышей с диабетом. [22] Ингибиторы GSK-3 также могут оказывать терапевтическое воздействие на геморрагическую трансформацию после острого ишемического инсульта. [38] GSK-3 может отрицательно регулировать сигнальный путь инсулина, ингибируя IRS1 посредством фосфорилирования серина-332. [20] делая рецептор инсулина неспособным активировать IRS1 и дополнительно инициировать канонический путь PI3K/Akt. Роль, которую ингибирование GSK-3 может играть в других его сигнальных функциях, еще не полностью понята.
Ингибирование GSK-3 также опосредует увеличение транскрипции транскрипционного фактора Tbet (Tbx21) и ингибирование транскрипции ингибирующего корецептора программируемой гибели клеток-1 (PD-1) на Т-клетках. [39] Ингибиторы GSK-3 повышали in vivo функцию CD8(+) OT-I CTL и клиренс вирусных инфекций, вызванных мышиным гамма-герпесвирусом 68 и клоном 13 лимфоцитарного хориоменингита, а также анти-PD-1 при иммунотерапии.
Ингибиторы
[ редактировать ]Ингибиторы киназы гликогенсинтазы относятся к различным хемотипам и имеют разные механизмы действия; это могут быть катионы из природных источников, синтетические АТФ- и не-АТФ-конкурентные ингибиторы и субстрат-конкурентные ингибиторы. GSK3 представляет собой билобарную архитектуру с N-концом и C-концом , N-конец отвечает за связывание АТФ, а C-конец, который называется петлей активации , опосредует киназную активность, тирозин, расположенный на C-конце, необходим для полная активность GSK3. [40]
Преимущества ингибиторов GSK-3β
[ редактировать ]При диабете ингибиторы GSK-3β повышают чувствительность к инсулину, синтез гликогена и метаболизм глюкозы в скелетных мышцах, уменьшают ожирение, влияя на процесс адипогенеза . [41] GSK-3β также чрезмерно экспрессируется при некоторых типах рака, таких как колоректальный рак , яичников и рак простаты . [40] Ингибиторы GSK-3β также помогают в лечении болезни Альцгеймера . [ нужна ссылка ] гладить , [38] и расстройства настроения , включая биполярное расстройство . [42]
Конкретные агенты
[ редактировать ]К ингибиторам ГСК-3 относятся: [43] [44] [45] [46]
Катионы металлов
[ редактировать ]АТФ-конкурентный
[ редактировать ]Полученные из морских организмов
[ редактировать ]- 6-БИО (IC 50 = 1,5 мкм)
- Дибромкантареллин (IC 50 = 3 мкм)
- Гимениалдезин (IC 50 = 10 нМ)
- Индирубин (IC 50 =5-50 нМ)
- Меридианин
Аминопиримидины
[ редактировать ]- CHIR99021 (IC 50 = 6,9–10 нМ)
- CHIR98014 (IC 50 = 0,58-0,65 нМ)
- CT98014
- CT98023
- CT99021
- TWS119 (IC 50 =30 нМ)
Арилиндолмалеимид
[ редактировать ]Тиазолы
[ редактировать ]- AR-A014418 (IC 50 =104 нМ)
- АЗД-1080 (ИК 50 =6,9нМ-31нМ)
Пауллонес
[ редактировать ]IC 50 = 4-80 нМ:
Алоизины
[ редактировать ]IC 50 =0,5-1,5 мкм:
Не-ATP конкурентный
[ редактировать ]Полученные из морских организмов
[ редактировать ]- Манзамин А (IC 50 = 1,5 мкм)
- Палинурин (IC 50 = 4,5 мкм)
- Трикантин (IC 50 =7,5 мкм)
Тиазолидиндионы
[ редактировать ]- ТДЗД-8 (ИК 50 =2мкМ)
- NP00111 (IC 50 = 2 мкм)
- NP031115 (IC 50 = 4 мкм)
- Тидеглузиб (IC 50 =60 нМ)
Галометилкетоны
[ редактировать ]- ХМК-32 (ИК 50 =1,5мкМ)
Пептиды
[ редактировать ]Неизвестный механизм (низкомолекулярные ингибиторы)
[ редактировать ]Литий
[ редактировать ]Литий, который используется при лечении биполярного расстройства, был первым обнаруженным природным ингибитором GSK-3. Он ингибирует GSK-3 напрямую за счет конкуренции с ионами магния и косвенно за счет фосфорилирования и ауторегуляции серина.Было обнаружено, что литий оказывает инсулиноподобное воздействие на метаболизм глюкозы, включая стимуляцию синтеза гликогена в жировых клетках, коже и мышцах, увеличение поглощения глюкозы и активацию активности GS. Помимо ингибирования GSK-3, он также ингибирует другие ферменты, участвующие в регуляции метаболизма глюкозы, такие как мио-инозитол-1-монофосфатаза и 1,6-бисфосфатаза. Кроме того, он показал терапевтическую пользу при болезни Альцгеймера и других нейродегенеративных заболеваниях, таких как эпилептическая нейродегенерация. [45]
Напроксен и Кромолин
[ редактировать ]Напроксен — нестероидный противовоспалительный препарат , а кромолин — противоаллергическое средство , действующее как стабилизатор тучных клеток . Оба препарата продемонстрировали противораковый эффект помимо гипогликемического эффекта за счет ингибирования киназы гликогенсинтазы-3β (GSK-3β).
Чтобы подтвердить гипотезу анти-GSK-3β напроксена и кромолина, в дополнение к измерению уровня глюкозы в сыворотке, инсулина в сыворотке было проведено стыковка двух структур с карманом связывания GSK-3β и сравнение их соответствия с известным ингибитором GSK-3β ARA014418. С-пептид сыворотки, изменение веса и уровни гликогена в печени для моделей нормальных и диабетических животных натощак для оценки их гипогликемических эффектов in vitro. [ нужна ссылка ]
Напроксен и кромолин были успешно пристыкованы к сайту связывания GSK-3β (оба были помещены в его связывающий карман). Они демонстрировали электростатические, гидрофобные и водородные взаимодействия с ключевыми аминокислотами внутри связывающего кармана с профилями взаимодействия связывания, аналогичными AR-A014418 (известному ингибитору). Отрицательные заряды групп карбоновой кислоты в обоих препаратах электростатически взаимодействуют с положительно заряженной гуанидиновой группой Arg141. Кроме того, наблюдаются взаимодействия водородных связей между карбоксильными фрагментами кромолина и аммониевыми группами Lys183 и Lys60, а также π-стэкинг нафталиновой кольцевой системы напроксена с фенольным кольцом Tyr134.
Противодиабетическое действие напроксена и кромолина. На нормальных животных моделях оба препарата показали дозозависимое снижение уровня глюкозы в крови и повышение уровня гликогена. В модели хронического диабета II типа уровни глюкозы также снижались, а уровень гликогена и уровень инсулина повышались дозозависимым образом со снижением уровня глюкозы в плазме. [ нужна ссылка ]
Эффекты напроксена и кромолина против ожирения: оба препарата продемонстрировали значительный эффект против ожирения, поскольку они снижают массу тела, уровень резистина и глюкозы в зависимости от дозы. Также было обнаружено, что они повышают уровни адипонектина , инсулина и С-пептида дозозависимым образом. [41]
Фамотидин
[ редактировать ]длительного действия Фамотидин — специфический антагонист H2 , снижающий секрецию желудочной кислоты. Он используется при лечении язвенной болезни, ГЭРБ и патологических гиперсекреторных состояний, таких как синдром Золлингера-Эллисона. (14,15) Антагонисты H2-рецепторов влияют на метаболизм гормонов, но их влияние на метаболизм глюкозы недостаточно изучено. (16) Исследование выявило сахароснижающий эффект фамотидина. [ нужна ссылка ]
Исследование связывания фамотидина с ферментом показало, что фамотидин может стыковаться внутри связывающего кармана GSK-3β, обеспечивая значительные взаимодействия с ключевыми точками внутри связывающего кармана GSK-3β. Взаимодействия сильных водородных связей с ключевыми аминокислотами PRO-136 и VAL-135 и потенциальное гидрофобное взаимодействие с LEU-188 были аналогичны тем, которые были обнаружены при связывании лиганда с ферментом (AR-A014418). [ нужна ссылка ]
Кроме того, фамотидин продемонстрировал высокую аффинность связывания GSK-3β и ингибирующую активность благодаря взаимодействиям, которые стабилизируют комплекс, а именно водородным связыванию гуанидиновой группы в фамотидине с сульфагидрильным фрагментом в CYS-199; электростатические взаимодействия той же гуанидиновой группы с карбоксильной группой в ASP-200, водородная связь между концевой группой NH2, OH TYR-143 и гидрофобное взаимодействие атома серы в тиоэфире с ILE-62. Исследования in vitro показали, что фамотидин ингибирует активность GSK-3β и увеличивает запасы гликогена в печени дозозависимым образом. Наблюдалось четырехкратное увеличение уровня гликогена в печени при применении максимальной дозы фамотидина (4,4 мг/кг). Кроме того, было показано, что фамотидин снижает уровень глюкозы в сыворотке через 30 и 60 минут после пероральной нагрузки глюкозой у здоровых людей. [47]
Куркумин
[ редактировать ]Куркумин, входящий в состав специи куркума , обладает вкусовыми и красящими свойствами. [48] Он имеет две симметричные формы: енол (наиболее распространенные формы) и кетон. [49] [50]
Куркумин обладает широкой фармакологической активностью: противовоспалительной, [51] противомикробный, [52] гипогликемическое, антиоксидантное и ранозаживляющее действие. [53] На животных моделях с болезнью Альцгеймера он оказывает антидеструктивное действие на бета-амилоид в головном мозге. [54] и в последнее время он проявляет противомалярийную активность. [55]
Куркумин также обладает химиопрофилактическим и противораковым действием. [ цитата не найдена ] и было показано, что он ослабляет окислительный стресс и почечную дисфункцию у животных с диабетом при хроническом применении. [56]
Механизм действия куркумина противовоспалительный; он ингибирует ядерный активатор транскрипции каппа B ( NF-KB ), который активируется всякий раз, когда возникает воспалительная реакция. [ нужна ссылка ]
NF-kB имеет два регуляторных фактора: IkB и GSK-3. [57] что предполагает, что куркумин напрямую связывает и ингибирует GSK-3B. Исследование in vitro подтвердило ингибирование GSK-3B путем моделирования молекулярного докинга с использованием метода докинга кремния. [58] Концентрация, при которой куркумин ингибирует 50% ГК-3В, составляет 66,3 нМ. [58]
Среди двух его форм экспериментальные и теоретические исследования показывают, что енольная форма является предпочтительной формой из-за ее внутримолекулярных водородных связей, а эксперимент ЯМР показывает, что енольная форма существует в различных растворителях. [ нужна ссылка ]
Оланзапин
[ редактировать ]Антипсихотические препараты все чаще используются при шизофрении , биполярном расстройстве, тревоге и других психических расстройствах. [59] Атипичные нейролептики используются чаще, чем антипсихотики первого поколения, поскольку они снижают риск экстрапирамидных симптомов, таких как поздняя дискинезия , и обладают большей эффективностью. [60]
Оланзапин и атипичные нейролептики вызывают увеличение веса за счет увеличения жировых отложений. [61] Он также влияет на метаболизм глюкозы, и некоторые исследования показывают, что он может ухудшить течение диабета. [62]
Недавнее исследование показывает, что оланзапин ингибирует активность GSK3, что позволяет предположить, что оланзапин способствует синтезу гликогена. Исследование влияния оланзапина на уровень глюкозы и гликогена в крови мышей показало значительное снижение уровня глюкозы в крови и повышение уровня гликогена у мышей, при этом IC50% оланзапина составляла 91,0 нм, что считается мощным ингибитором. Исследование также показывает, что субхроническое применение оланзапина приводит к мощному ингибированию GSK3. [42]
Производные пиримидина
[ редактировать ]Аналоги пиримидина являются антиметаболитами, препятствующими синтезу нуклеиновых кислот. [63] Было показано, что некоторые из них соответствуют АТФ-связывающему карману GSK-3β, снижая уровень глюкозы в крови и улучшая некоторые нейрональные заболевания. [64]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ PDB : 1J1B ; Аоки М., Ёкота Т., Сугиура И., Сасаки С., Хасегава Т., Окумура С. и др. (март 2004 г.). «Структурное понимание распознавания нуклеотидов в тау-протеинкиназе I/гликогенсинтазы киназы 3 бета». Акта Кристаллографика. Раздел D. Биологическая кристаллография . 60 (Часть 3): 439–446. Бибкод : 2004AcCrD..60..439A . дои : 10.1107/S090744490302938X . ПМИД 14993667 .
- ^ Jump up to: а б Эмби Н., Рилатт Д.Б., Коэн П. (июнь 1980 г.). «Киназа гликогенсинтазы-3 из скелетных мышц кролика. Отделение от циклической АМФ-зависимой протеинкиназы и киназы фосфорилазы» . Европейский журнал биохимии . 107 (2): 519–527. дои : 10.1111/j.1432-1033.1980.tb06059.x . ПМИД 6249596 .
- ^ Бойрел Э., Грико С.Ф., Джоуп Р.С. (апрель 2015 г.). «Киназа гликогенсинтазы-3 (GSK3): регуляция, действие и заболевания» . Фармакология и терапия . 148 : 114–131. doi : 10.1016/j.pharmthera.2014.11.016 . ПМК 4340754 . ПМИД 25435019 .
- ^ Jump up to: а б с Джоуп Р.С., Джонсон Г.В. (февраль 2004 г.). «Гламур и мрак киназы гликогенсинтазы-3». Тенденции биохимических наук . 29 (2): 95–102. дои : 10.1016/j.tibs.2003.12.004 . ПМИД 15102436 .
- ^ Турлик Ю., Васикевич Э., Домарадска А., Хростек Г., Гняджик В., Домагальский М., Дуда П. (декабрь 2021 г.). «Активность GSK3β в функционировании схемы вознаграждения и зависимости» . Нейронаука . 2 (4): 443–466. дои : 10.3390/neurosci2040033 . ISSN 2673-4087 .
- ^ Пандей МК, ДеГрадо Т.Р. (2016). «Таргетная терапия и визуализация гликогенсинтазокиназы-3 (GSK-3)» . Тераностика . 6 (4): 571–593. дои : 10.7150/thno.14334 . ПМЦ 4775866 . ПМИД 26941849 .
- ^ Али А., Хефлих К.П., Вуджетт-младший (август 2001 г.). «Киназа гликогенсинтазы-3: свойства, функции и регуляция». Химические обзоры . 101 (8): 2527–2540. дои : 10.1021/cr000110o . ПМИД 11749387 .
- ^ Эльдар-Финкельман Х (март 2002 г.). «Киназа гликогенсинтазы 3: новая терапевтическая мишень». Тенденции молекулярной медицины . 8 (3): 126–132. дои : 10.1016/S1471-4914(01)02266-3 . ПМИД 11879773 .
- ^ Хупер С., Киллик Р., Лавстон С. (март 2008 г.). «Гипотеза GSK3 болезни Альцгеймера» . Журнал нейрохимии . 104 (6): 1433–1439. дои : 10.1111/j.1471-4159.2007.05194.x . ПМЦ 3073119 . ПМИД 18088381 .
- ^ Jump up to: а б Эрмида М.А., Кумар Дж.Д., Лесли Н.Р. (август 2017 г.). «GSK3 и его взаимодействие с сигнальной сетью PI3K/AKT/mTOR» . Достижения в области биологического регулирования . 65 : 5–15. дои : 10.1016/j.jbior.2017.06.003 . ПМИД 28712664 . Проверено 15 декабря 2023 г.
- ^ Jump up to: а б Ли Ц, Чжао Ц, Чжан Дж, Линкан Л, Вэньхао В, Чуа Б, Чэнь Ю, Сюй Л, Ли П (24 сентября 2019 г.). «Комплекс протеинфосфатазы 1 является прямой мишенью АКТ, которая связывает передачу сигналов инсулина с отложением гликогена в печени» . Отчеты по ячейкам . 28 (13): 3406–3422. дои : 10.1016/j.celrep.2019.08.066 . ПМИД 31553910 .
- ^ Гликоген + синтаза + киназа Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- ^ ГСК1 , ГСК2 . NCBI Ген .
- ^ Даджани Р., Фрейзер Э., Роу С.М., Янг Н., Гуд В., Дейл Т.К., Перл Л.Х. (июнь 2001 г.). «Кристаллическая структура киназы гликогенсинтазы 3 бета: структурная основа специфичности фосфат-примированного субстрата и аутоингибирования» . Клетка . 105 (6): 721–732. дои : 10.1016/S0092-8674(01)00374-9 . ПМИД 11440715 . S2CID 17401752 .
- ^ Бускила М., Хантер Р.В., Ибрагим А.Ф., Делатр Л., Пегги М., ван Дипен Дж.А. и др. (ноябрь 2010 г.). «Аллостерическая регуляция гликогенсинтазы контролирует синтез гликогена в мышцах» . Клеточный метаболизм . 12 (5): 456–466. дои : 10.1016/j.cmet.2010.10.006 . ПМИД 21035757 .
- ^ Вуджетт-младший (август 1994 г.). «Регуляция и функции подсемейства гликогенсинтазной киназы-3». Семинары по биологии рака . 5 (4): 269–275. ПМИД 7803763 .
- ^ Вуджетт-младший (сентябрь 2001 г.). «Судить о белке не только по его названию: GSK-3». СТКЭ науки . 2001 (100): re12. дои : 10.1126/stke.2001.100.re12 . ПМИД 11579232 . S2CID 19052833 .
- ^ Али А., Хёфлих К.П., Вуджетт-младший (август 2001 г.). «Киназа гликогенсинтазы-3: свойства, функции и регуляция». Химические обзоры . 101 (8): 2527–2540. дои : 10.1021/cr000110o . ПМИД 11749387 . S2CID 12925005 .
- ^ Jump up to: а б с д Раясам Г.В., Туласи В.К., Содхи Р., Дэвис Дж.А., Рэй А. (март 2009 г.). «Киназа гликогенсинтазы 3: больше, чем тезка» . Британский журнал фармакологии . 156 (6): 885–898. дои : 10.1111/j.1476-5381.2008.00085.x . ПМК 2697722 . ПМИД 19366350 .
- ^ Jump up to: а б Либерман З., Эльдар-Финкельман Х (февраль 2005 г.). «Фосфорилирование серина 332 субстрата инсулинового рецептора-1 киназой гликогенсинтазы-3 ослабляет передачу сигналов инсулина» . Журнал биологической химии . 280 (6): 4422–4428. дои : 10.1074/jbc.M410610200 . ПМИД 15574412 .
- ^ Локхед, Пенсильвания, Коглан М., Райс С.К., Сазерленд К. (май 2001 г.). «Ингибирование GSK-3 избирательно снижает экспрессию генов глюкозо-6-фосфатазы, фосфатазы и фосфоенолипируваткарбоксикиназы» . Диабет . 50 (5): 937–946. дои : 10.2337/диабет.50.5.937 . ПМИД 11334436 .
- ^ Jump up to: а б с д и ж г Джоуп Р.С., Юскайтис С.Дж., Берел Э. (апрель – май 2007 г.). «Киназа гликогенсинтазы-3 (GSK3): воспаление, заболевания и терапия» . Нейрохимические исследования . 32 (4–5): 577–595. дои : 10.1007/s11064-006-9128-5 . ЧВК 1970866 . ПМИД 16944320 .
- ^ Jump up to: а б Ван Х., Браун Дж., Мартин М. (февраль 2011 г.). «Киназа гликогенсинтазы 3: точка конвергенции воспалительной реакции хозяина» . Цитокин . 53 (2): 130–140. дои : 10.1016/j.cyto.2010.10.009 . ПМК 3021641 . ПМИД 21095632 .
- ^ Миллс К.Н., Новшин С., Боннер Дж.А., Ян Э.С. (2011). «Новая роль киназы гликогенсинтазы 3 в лечении опухолей головного мозга» . Границы молекулярной нейронауки . 4 : 47. doi : 10.3389/fnmol.2011.00047 . ПМЦ 3223722 . ПМИД 22275880 .
- ^ Jump up to: а б с Глибо М., Серман А., Карин-Куюнджич В., Бекавац Влаткович И., Мишкович Б., Вранич С., Серман Л. (февраль 2021 г.). «Роль киназы гликогенсинтазы 3 (GSK3) при раке с акцентом на развитие и прогрессирование рака яичников: всесторонний обзор» . Боснийский журнал фундаментальных медицинских наук . 21 (1): 5–18. дои : 10.17305/bjbms.2020.5036 . ПМЦ 7861620 . ПМИД 32767962 .
- ^ Ваттарасит П., Бижур Г.Н., Змиевски Дж.В., Сонг Л., Змиевска А., Чен X и др. (июнь 2002 г.). «Прямое, активирующее взаимодействие между киназой гликогенсинтазы-3бета и р53 после повреждения ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (12): 7951–7955. Бибкод : 2002PNAS...99.7951W . дои : 10.1073/pnas.122062299 . ПМК 123001 . ПМИД 12048243 .
- ^ Граймс Калифорния, Джоуп Р.С. (сентябрь 2001 г.). «Активность связывания ДНК CREB ингибируется гликогенсинтазой-киназой-3 бета и облегчается литием» . Журнал нейрохимии . 78 (6): 1219–1232. дои : 10.1046/j.1471-4159.2001.00495.x . ПМК 1947002 . ПМИД 11579131 .
- ^ Jump up to: а б Котлиарова С., Пасторино С., Ковелл Л.С., Котляров Ю., Сонг Х., Чжан В. и др. (август 2008 г.). «Ингибирование киназы гликогенсинтазы-3 индуцирует гибель клеток глиомы посредством c-MYC, ядерного фактора-каппаВ и регуляции глюкозы» . Исследования рака . 68 (16): 6643–6651. дои : 10.1158/0008-5472.CAN-08-0850 . ПМЦ 2585745 . ПМИД 18701488 .
- ^ Джейкобс К.М., Бхаве С.Р., Ферраро DJ, Джабоин Дж.Дж., Халлахан Д.Е., Тотала Д. (май 2012 г.). «GSK-3β: бифункциональная роль в путях гибели клеток» . Международный журнал клеточной биологии . 2012 : 930710. doi : 10.1155/2012/930710 . ПМЦ 3364548 . ПМИД 22675363 .
- ^ Добл Б.В., Вуджетт-младший (апрель 2003 г.). «GSK-3: хитрости создания многозадачной киназы» . Журнал клеточной науки . 116 (Часть 7): 1175–1186. дои : 10.1242/jcs.00384 . ПМК 3006448 . ПМИД 12615961 .
- ^ Биджур Г.Н., Джоуп Р.С. (декабрь 2003 г.). «Киназа гликогенсинтазы-3 бета сильно активируется в ядрах и митохондриях». НейроОтчёт . 14 (18): 2415–2419. дои : 10.1097/00001756-200312190-00025 . ПМИД 14663202 . S2CID 43633965 .
- ^ Сарасвати А.П., Али Хуссаини С.М., Кришна Н.Х., Бабу Б.Н., Камаль А. (январь 2018 г.). «Киназа гликогенсинтазы-3 и ее ингибиторы: потенциальная мишень для различных терапевтических состояний». Европейский журнал медицинской химии . 144 : 843–858. дои : 10.1016/j.ejmech.2017.11.103 . ПМИД 29306837 .
- ^ Инь Л., Ван Дж., Кляйн П.С., Лазар М.А. (февраль 2006 г.). «Ядерный рецептор Rev-erbalpha является важным чувствительным к литию компонентом циркадных часов». Наука . 311 (5763): 1002–1005. Бибкод : 2006Sci...311.1002Y . дои : 10.1126/science.1121613 . ПМИД 16484495 . S2CID 11240826 .
- ^ Рыбаковски Ю.К., Дмитрзак-Вегларц М., Дембинска-Краевска Д., Хаузер Дж., Акискал К.К., Акискал Х.Х. (апрель 2014 г.). «Полиморфизм генов циркадных часов и темпераментных размеров TEMPS-A при биполярном расстройстве». Журнал аффективных расстройств . 159 : 80–84. дои : 10.1016/j.jad.2014.02.024 . ПМИД 24679394 .
- ^ Ху С., Бегум А.Н., Джонс М.Р., О М.С., Бич В.К., Бич Б.Х. и др. (февраль 2009 г.). «Ингибиторы GSK3 демонстрируют преимущества в модели нейродегенерации болезни Альцгеймера (БА), но побочные эффекты у контрольных животных» . Нейробиология болезней . 33 (2): 193–206. дои : 10.1016/j.nbd.2008.10.007 . ПМЦ 4313761 . ПМИД 19038340 .
- ^ Ван З., Смит К.С., Мерфи М., Пилото О., Сомервейл Т.С., Клири М.Л. (октябрь 2008 г.). «Киназа гликогенсинтазы 3 в поддержании и таргетной терапии лейкоза MLL» . Природа . 455 (7217): 1205–1209. Бибкод : 2008Natur.455.1205W . дои : 10.1038/nature07284 . ПМК 4084721 . ПМИД 18806775 .
- ^ Маршан Б., Трамбле И., Каньоль С., Буше М.Дж. (март 2012 г.). «Ингибирование активности гликогенсинтазы-киназы-3 запускает апоптотический ответ в клетках рака поджелудочной железы посредством JNK-зависимых механизмов» . Канцерогенез . 33 (3): 529–537. дои : 10.1093/carcin/bgr309 . ПМИД 22201186 .
- ^ Jump up to: а б Ван В., Ли М., Ван Ю., Ли К., Дэн Г., Ван Дж. и др. (декабрь 2016 г.). «Ингибитор GSK-3β TWS119 ослабляет геморрагическую трансформацию, индуцированную rtPA, и активирует сигнальный путь Wnt/β-катенин после острого ишемического инсульта у крыс» . Молекулярная нейробиология . 53 (10): 7028–7036. дои : 10.1007/s12035-015-9607-2 . ПМЦ 4909586 . ПМИД 26671619 .
- ^ Тейлор А., Харкер Дж.А., Чантхонг К., Стивенсон П.Г., Зунига Э.И., Радд CE (февраль 2016 г.). «Инактивация киназы гликогенсинтазы 3 приводит к опосредованному T-bet снижению регуляции корецептора PD-1 для усиления ответов цитолитических Т-клеток CD8 (+)» . Иммунитет . 44 (2): 274–286. doi : 10.1016/j.immuni.2016.01.018 . ПМК 4760122 . ПМИД 26885856 .
- ^ Jump up to: а б Саяс CL, Ариенс А, Понсиен Б, Муленаар WH (апрель 2006 г.). «GSK-3 активируется тирозинкиназой Pyk2 во время LPA1-опосредованной ретракции нейритов» . Молекулярная биология клетки . 17 (4): 1834–1844. doi : 10.1091/mbc.E05-07-0688 . ПМЦ 1415316 . ПМИД 16452634 .
- ^ Jump up to: а б Мотави Т.М., Бустанджи И., Эль-Мараги С.А., Таха М.О., Аль Гусейн М.А. (сентябрь 2013 г.). «Напроксен и кромолин как новые ингибиторы киназы гликогенсинтазы 3β для улучшения состояния при диабете и ожирении: исследование путем моделирования стыковки и последующей биохимической оценки in vitro/in vivo». Журнал биохимической и молекулярной токсикологии . 27 (9): 425–436. дои : 10.1002/jbt.21503 . ПМИД 23784744 . S2CID 46597394 .
- ^ Jump up to: а б Мохаммад М.К., Аль-Масри И.М., Таха М.О., Аль-Гусейн М.А., Алхатиб Х.С., Наджар С., Бустанджи И. (апрель 2008 г.). «Оланзапин ингибирует киназу-3бета гликогенсинтазы: исследование путем моделирования стыковки и экспериментальной проверки». Европейский журнал фармакологии . 584 (1): 185–191. дои : 10.1016/j.ejphar.2008.01.019 . ПМИД 18295757 .
- ^ Нури М.С., Бхатт П.М., Куррегес М.К., Газанфари Д., Каклер С., Орак К.М. и др. (декабрь 2019 г.). «Идентификация нового селективного и мощного ингибитора киназы гликогенсинтазы-3» . Американский журнал физиологии. Клеточная физиология . 317 (6): C1289–C1303. doi : 10.1152/ajpcell.00061.2019 . ПМК 6962522 . ПМИД 31553649 .
- ^ Лихт-Мурава А., Пас Р., Вакс Л., Авраами Л., Плоткин Б., Эйзенштейн М., Эльдар-Финкельман Х. (ноябрь 2016 г.). «Уникальный тип ингибитора GSK-3 открывает новые возможности для клиники». Научная сигнализация . 9 (454): ра110. дои : 10.1126/scisignal.aah7102 . ПМИД 27902447 . S2CID 34207388 .
- ^ Jump up to: а б Эльдар-Финкельман Х., Мартинес А. (2011). «Ингибиторы GSK-3: доклинические и клинические исследования ЦНС» . Границы молекулярной нейронауки . 4 : 32. doi : 10.3389/fnmol.2011.00032 . ПМК 3204427 . ПМИД 22065134 .
- ^ МакКубри Дж.А., Стилман Л.С., Бертран Ф.Е., Дэвис Н.М., Соколоски М., Абрамс С.Л. и др. (май 2014 г.). «GSK-3 как потенциальная мишень для терапевтического вмешательства при раке» . Онкотаргет . 5 (10): 2881–2911. дои : 10.18632/oncotarget.2037 . ПМК 4102778 . ПМИД 24931005 .
- ^ Мохаммад М., Аль-Масри И.М., Исса А., Аль-Гусейн М.А., Фарарьех М., Алхатиб Х. и др. (август 2013 г.). «Фамотидин ингибирует киназу-3β гликогенсинтазы: исследование путем моделирования стыковки и экспериментальной проверки» . Журнал ингибирования ферментов и медицинской химии . 28 (4): 690–694. дои : 10.3109/14756366.2012.672413 . ПМИД 22512725 . S2CID 11890710 .
- ^ Махешвари Р.К., Сингх А.К., Гаддипати Дж., Шримал Р.К. (март 2006 г.). «Множественная биологическая активность куркумина: краткий обзор». Науки о жизни . 78 (18): 2081–2087. дои : 10.1016/j.lfs.2005.12.007 . ПМИД 16413584 .
- ^ Баласубраманян К. (май 2006 г.). «Молекулярная орбитальная основа профилактики болезни Альцгеймера куркумином со специей желтого карри». Журнал сельскохозяйственной и пищевой химии . 54 (10): 3512–3520. дои : 10.1021/jf0603533 . ПМИД 19127718 .
- ^ Пэйтон Ф., Сандаски П., Алворт В.Л. (февраль 2007 г.). «ЯМР-исследование структуры раствора куркумина». Журнал натуральных продуктов . 70 (2): 143–146. дои : 10.1021/np060263s . ПМИД 17315954 .
- ^ Кохли К., Али Дж., Ансари М.Дж., Рахеман З. (2005). «Куркумин: природное противовоспалительное средство» . Индийский журнал фармакологии . 37 (3): 141. дои : 10.4103/0253-7613.16209 . hdl : 1807/8668 .
- ^ Неги П.С., Джаяпракаша Г.К., Джаган Мохан Рао Л., Сакария К.К. (октябрь 1999 г.). «Антибактериальная активность масла куркумы: побочный продукт производства куркумина». Журнал сельскохозяйственной и пищевой химии . 47 (10): 4297–4300. дои : 10.1021/jf990308d . ПМИД 10552805 .
- ^ Сидху Г.С., Сингх А.К., Талур Д., Банаудха К.К., Патнаик Г.К., Шримал Р.К., Махешвари Р.К. (1998). «Усиление заживления ран куркумином у животных». Заживление и регенерация ран . 6 (2): 167–177. дои : 10.1046/j.1524-475X.1998.60211.x . ПМИД 9776860 . S2CID 21440334 .
- ^ Ян Ф., Лим Г.П., Бегум А.Н., Убеда О.Дж., Симмонс М.Р., Амбегаокар С.С. и др. (февраль 2005 г.). «Куркумин ингибирует образование бета-олигомеров и фибрилл бета-амилоида, связывает бляшки и уменьшает количество амилоида in vivo» . Журнал биологической химии . 280 (7): 5892–5901. дои : 10.1074/jbc.M404751200 . ПМИД 15590663 .
- ^ Мишра С., Кармодия К., Суролия Н., Суролия А. (март 2008 г.). «Синтез и исследование новых аналогов куркумина в качестве противомалярийных средств». Биоорганическая и медицинская химия . 16 (6): 2894–2902. дои : 10.1016/j.bmc.2007.12.054 . ПМИД 18194869 .
- ^ Шарма С., Кулкарни С.К., Чопра К. (октябрь 2006 г.). «Куркумин, действующее вещество куркумы (Curcuma longa), облегчает диабетическую нефропатию у крыс». Клиническая и экспериментальная фармакология и физиология . 33 (10): 940–945. дои : 10.1111/j.1440-1681.2006.04468.x . ПМИД 17002671 . S2CID 25193929 .
- ^ Демарчи Ф., Бертоли С., Сэнди П., Шнайдер С. (октябрь 2003 г.). «Киназа гликогенсинтазы-3 бета регулирует стабильность NF-каппа B1/p105» . Журнал биологической химии . 278 (41): 39583–39590. дои : 10.1074/jbc.M305676200 . ПМИД 12871932 .
- ^ Jump up to: а б Бустанджи И., Таха М.О., Алмасри И.М., Аль-Гусейн М.А., Мохаммад М.К., Алхатиб Х.С. (июнь 2009 г.). «Ингибирование киназы гликогенсинтазы куркумином: исследование путем моделирования молекулярной стыковки и последующей оценки in vitro/in vivo». Журнал ингибирования ферментов и медицинской химии . 24 (3): 771–778. дои : 10.1080/14756360802364377 . ПМИД 18720192 . S2CID 23137441 .
- ^ «Нейролептики АЗ» . Mind.org.uk. 2018. [ ненадежный медицинский источник? ]
- ^ «Нейролептики при биполярном расстройстве» . ВебМД.
- ^ Гуди А.Дж., Смит Дж.А., Хэлфорд Дж.К. (декабрь 2002 г.). «Характеристика увеличения веса у крыс, вызванного оланзапином». Журнал психофармакологии . 16 (4): 291–296. дои : 10.1177/026988110201600402 . ПМИД 12503827 . S2CID 23589812 .
- ^ Ди Лоренцо Р., Брогли А (сентябрь 2010 г.). «Профиль инъекции оланзапина длительного действия для поддерживающего лечения взрослых пациентов с шизофренией» . Нервно-психические заболевания и лечение . 6 : 573–581. дои : 10.2147/NDT.S5463 . ПМЦ 2938306 . ПМИД 20856920 .
- ^ Мерфи Ф., Миддлтон М. (2012). «Цитостатические и цитотоксические препараты». Всемирный ежегодный обзор новых данных о побочных реакциях и взаимодействиях лекарств . Побочные эффекты лекарств. Ежегодник. Том. 34. стр. 731–747. дои : 10.1016/B978-0-444-59499-0.00045-3 . ISBN 978-0-444-59499-0 .
- ^ Крамер Т., Шмидт Б., Ло Монте Ф. (2012). «Низкомолекулярные ингибиторы GSK-3: структурные данные и их применение к моделям болезни Альцгеймера» . Международный журнал болезни Альцгеймера . 2012 : 381029. doi : 10.1155/2012/381029 . ПМЦ 3408674 . ПМИД 22888461 .
Внешние ссылки
[ редактировать ]- Киназа гликогенсинтазы 3 в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)