Активный сайт

В биологии и биохимии активный центр — это область фермента , где молекулы субстрата связываются и вступают в химическую реакцию. Активный центр состоит из аминокислотных остатков, которые образуют временные связи с субстратом, сайтом связывания , и остатков, катализирующих реакцию этого субстрата, каталитического сайта . Хотя активный центр занимает всего ~10–20% объема фермента, [ 1 ] : 19 это самая важная часть, поскольку она непосредственно катализирует химическую реакцию . Обычно он состоит из трех-четырех аминокислот, тогда как другие аминокислоты в белке необходимы для поддержания третичной структуры ферментов. [ 2 ]
Каждый активный центр оптимизирован для связывания определенного субстрата и катализа определенной реакции, что приводит к высокой специфичности . Эта специфичность определяется расположением аминокислот в активном центре и структурой субстратов. Иногда ферментам также необходимо связываться с некоторыми кофакторами для выполнения своей функции. Активный центр обычно представляет собой бороздку или карман фермента, который может располагаться в глубоком туннеле внутри фермента. [ 3 ] или между интерфейсами мультимерных ферментов . Активный центр может многократно катализировать реакцию, поскольку остатки не изменяются в конце реакции (они могут меняться во время реакции, но к концу регенерируются). [ 4 ] Этот процесс достигается за счет снижения энергии активации реакции, поэтому больше субстратов имеет достаточно энергии для реакции.
Связывающий сайт
[ редактировать ]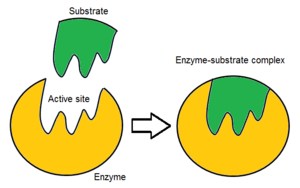
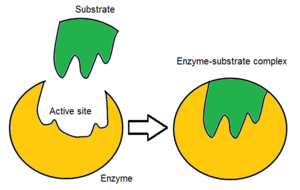
Обычно молекула фермента имеет только один активный центр, и этот активный центр соответствует одному конкретному типу субстрата. Активный центр содержит сайт связывания, который связывает субстрат и ориентирует его для катализа. Ориентация субстрата и непосредственная близость между ним и активным центром настолько важны, что в некоторых случаях фермент все еще может нормально функционировать, даже если все остальные части мутируют и теряют функцию. [ 5 ]
Первоначально взаимодействие между активным центром и субстратом является нековалентным и временным. Существует четыре важных типа взаимодействия, которые удерживают субстрат в определенной ориентации и образуют комплекс фермент-субстрат (ES-комплекс): водородные связи , взаимодействия Ван-дер-Ваальса , гидрофобные взаимодействия и электростатические силовые взаимодействия. [ 6 ] : 148 Распределение заряда на подложке и активном сайте должно быть дополняющим, что означает, что все положительные и отрицательные заряды должны быть нейтрализованы. В противном случае возникнет сила отталкивания, раздвигающая их. Активный центр обычно содержит неполярные аминокислоты, хотя иногда могут встречаться и полярные аминокислоты. [ 2 ] Связывание субстрата с сайтом связывания требует как минимум трех точек контакта для достижения стерео-, регио- и энантиоселективности. Например, алкогольдегидрогеназа , катализирующая перенос гидрид- иона от этанола к НАД. + субстрата взаимодействует с метильной группой , гидроксильной группой и про- (R) водородом, который будет отщепляться в ходе реакции. [ 6 ] : 149
Чтобы выполнять свою функцию, ферментам необходимо принять правильную складку белка ( нативную складку ) и третичную структуру . Чтобы поддерживать эту определенную трехмерную структуру, белки полагаются на различные типы взаимодействий между их аминокислотными остатками. Если этим взаимодействиям мешают, например, экстремальные значения pH, высокая температура или высокие концентрации ионов, это приведет к денатурации фермента и потере его каталитической активности. [ нужна ссылка ]
Считается, что более плотное прилегание активного центра к молекуле субстрата повышает эффективность реакции. Если плотность между активным центром ДНК-полимеразы и ее субстратом увеличивается, точность, то есть правильная скорость репликации ДНК, также увеличивается. [ 7 ] Большинство ферментов имеют глубоко спрятанные активные центры, к которым субстрат может получить доступ через каналы доступа. [ 3 ]
Существует три предложенные модели того, как ферменты соответствуют своему специфическому субстрату: модель замка и ключа , модель индуцированного соответствия и модель конформационного отбора. Последние два не исключают друг друга: за конформационным отбором может последовать изменение формы фермента. Кроме того, белок может не полностью следовать ни одной из моделей. Аминокислоты в сайте связывания убиквитина обычно следуют модели индуцированного соответствия, тогда как остальная часть белка обычно придерживается конформационного отбора. Такие факторы, как температура, вероятно, влияют на путь, выбранный во время связывания, при этом ожидается, что более высокие температуры повысят важность конформационного отбора и уменьшат важность индуцированного соответствия. [ 8 ]
Гипотеза замка и ключа
[ редактировать ]Эту концепцию предложил химик XIX века Эмиль Фишер . Он предположил, что активный центр и субстрат представляют собой две стабильные структуры, которые идеально подходят друг к другу без каких-либо дальнейших модификаций, как ключ в замок. Если один субстрат идеально связывается со своим активным центром, взаимодействия между ними будут наиболее сильными, что приведет к высокой каталитической эффективности.
Со временем начали проявляться ограничения этой модели. Например, конкурентный ингибитор ферментов метилглюкозид способен прочно связываться с активным центром 4-альфа-глюканотрансферазы и прекрасно в него вписывается. Однако 4-альфа-глюканотрансфераза не активна в отношении метилглюкозида, и перенос гликозила не происходит. Гипотеза «Замок и ключ» не может объяснить это, поскольку она предсказывает высокую эффективность переноса гликозила метилглюкозида из-за его прочного связывания. Помимо конкурентного ингибирования, эта теория не может объяснить и механизм действия неконкурентных ингибиторов , поскольку они не связываются с активным центром, но тем не менее влияют на каталитическую активность. [ 9 ]
Гипотеза индуцированной подгонки
[ редактировать ]Теория Дэниела Кошланда о связывании фермента с субстратом заключается в том, что активный центр и связывающая часть субстрата не совсем дополняют друг друга. [ 10 ] Модель индуцированной подгонки представляет собой развитие модели «замок и ключ» и предполагает, что активный центр является гибким и меняет форму до тех пор, пока подложка не будет полностью связана. Эта модель похожа на человека, носящего перчатку: перчатка меняет форму, подстраиваясь под руку. Фермент изначально имеет конформацию, притягивающую его субстрат. Поверхность фермента гибкая, и только правильный катализатор может вызвать взаимодействие, ведущее к катализу. Конформационные изменения могут произойти при связывании субстрата. После этого продукты реакции отойдут от фермента и активный центр вернется к своей первоначальной форме. Эта гипотеза подтверждается наблюдением, что весь белковый домен может перемещаться на несколько нанометров во время катализа. Это движение поверхности белка может создавать микроокружение, благоприятствующее катализу. [ 5 ]
Гипотеза конформационного отбора
[ редактировать ]Эта модель предполагает, что ферменты существуют в различных конформациях, только некоторые из которых способны связываться с субстратом. При связывании субстрата с белком равновесие в конформационном ансамбле смещается в сторону лигандов, способных связывать лиганды (поскольку ферменты со связанными субстратами выводятся из равновесия между свободными конформациями). [ 11 ]
Виды нековалентных взаимодействий
[ редактировать ]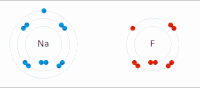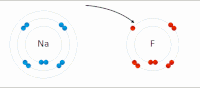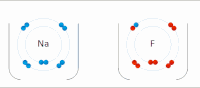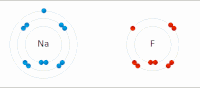


Электростатическое взаимодействие . В водной среде противоположно заряженные группы в боковых цепях аминокислот в активном центре и субстратах притягивают друг друга, что называется электростатическим взаимодействием. Например, когда карбоновая кислота (R-COOH) диссоциирует на RCOO. − и Х + ионы, COO − притягивает положительно заряженные группы, такие как протонированная гуанидиновая боковая цепь аргинина . [ нужна ссылка ]
Водородная связь : Водородная связь представляет собой особый тип диполь-дипольного взаимодействия между частично положительным атомом водорода и частично отрицательным донором электронов , которые содержат пару электронов, таких как кислород , фтор и азот . Прочность водородной связи зависит от химической природы и геометрического расположения каждой группы. [ нужна ссылка ]
Сила Ван-дер-Ваальса : Сила Ван-дер-Ваальса образуется между противоположно заряженными группами из-за временного неравномерного распределения электронов в каждой группе. Если все электроны сосредоточены на одном полюсе группы, этот конец будет отрицательным, а другой конец — положительным. Хотя индивидуальная сила слаба, поскольку общее количество взаимодействий между активным центром и подложкой огромно, сумма их будет значительной. [ нужна ссылка ]
Гидрофобное взаимодействие : Неполярные гидрофобные группы имеют тенденцию агрегироваться в водной среде и пытаться покинуть полярный растворитель. Эти гидрофобные группы обычно имеют длинную углеродную цепь и не реагируют с молекулами воды. При растворении в воде молекула белка сворачивается в шарообразную форму, оставляя гидрофильные группы снаружи, а гидрофобные группы глубоко погружены в центр. [ нужна ссылка ]
Каталитический сайт
[ редактировать ]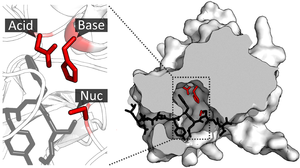
Как только субстрат связан и ориентирован в активном центре, катализ может начаться . Остатки каталитического сайта обычно расположены очень близко к сайту связывания, а некоторые остатки могут выполнять двойную роль как в связывании, так и в катализе. [ нужна ссылка ]
Каталитические остатки сайта взаимодействуют с субстратом, снижая энергию активации реакции и тем самым ускоряя ее протекание . Они делают это с помощью ряда различных механизмов, включая сближение реагентов, нуклеофильный/электрофильный катализ и кислотно-основной катализ. Эти механизмы будут объяснены ниже. [ нужна ссылка ]
Механизмы, участвующие в каталитическом процессе
[ редактировать ]Приближение реагента
[ редактировать ]Во время ферментативной каталитической реакции субстрат и активный центр располагаются в непосредственной близости. Этот подход имеет различные цели. Во-первых, когда субстраты связываются внутри активного центра, эффективная концентрация их значительно увеличивается, чем в растворе. Это означает, что количество молекул субстрата, участвующих в реакции, также увеличивается. Этот процесс также уменьшает энергию десольватации, необходимую для протекания реакции. В растворе молекулы субстрата окружены молекулами растворителя, и молекулам фермента требуется энергия для их замещения и контакта с субстратом. Поскольку объемные молекулы могут быть исключены из активного центра, выход энергии можно свести к минимуму. Затем активный центр предназначен для переориентации субстрата, чтобы уменьшить энергию активации, необходимую для протекания реакции. Выравнивание подложки после связывания фиксируется в высокоэнергетическом состоянии и может перейти к следующему этапу. Кроме того, этому связыванию благоприятствует энтропия , поскольку затраты энергии, связанные с реакцией растворения, в значительной степени устраняются, поскольку растворитель не может проникнуть в активный центр. В конце концов, активный сайт может манипулировать Молекулярную орбиталь подложки принимают в подходящую ориентацию для уменьшения энергии активации. [ 6 ] : 155–8
Электростатические состояния подложки и активного центра должны быть дополнительны друг другу. Поляризованная отрицательно заряженная боковая цепь аминокислоты отталкивает незаряженный субстрат. Но если переходное состояние включает образование ионного центра , то боковая цепь теперь будет производить благоприятное взаимодействие.
Ковалентный катализ
[ редактировать ]Многие ферменты, включая сериновую протеазу , цистеиновую протеазу , протеинкиназу и фосфатазу, эволюционировали так, чтобы образовывать временные ковалентные связи между ними и их субстратами, чтобы снизить энергию активации и позволить реакции произойти. Этот процесс можно разделить на 2 этапа: формирование и распад. Первый этап является этапом ограничения скорости, тогда как более поздний этап необходим для регенерации интактного фермента. [ 6 ] : 158
Нуклеофильный катализ фермента . Этот процесс включает в себя передачу электронов от нуклеофила субстрату для образования ковалентной связи между ними во время переходного состояния. Сила этого взаимодействия зависит от двух аспектов: способности нуклеофильной группы отдавать электроны и способности электрофила их принимать. На первый в основном влияет основность (способность отдавать электронные пары) вида, а на второй - его p K a . На обе группы также влияют их химические свойства, такие как поляризуемость , электроотрицательность и потенциал ионизации . Аминокислоты, способные образовывать нуклеофилы, включают серин , цистеин , аспартат и глутамин . [ нужна ссылка ]
Электрофильный катализ : Механизм этого процесса точно такой же, как и нуклеофильный катализ, за исключением того, что теперь аминокислоты в активном центре действуют как электрофилы , а субстраты являются нуклеофилами . Для этой реакции обычно требуются кофакторы, поскольку боковые цепи аминокислот недостаточно сильны для притяжения электронов.
Ионы металлов
[ редактировать ]Ионы металлов играют несколько ролей в ходе реакции. Во-первых, он может связываться с отрицательно заряженными группами субстрата, поэтому они не будут отталкивать электронные пары от нуклеофильных групп активного центра. Он может притягивать отрицательно заряженные электроны, увеличивая электрофильность . Он также может служить мостом между активным центром и субстратом. Наконец, они могут изменить конформационную структуру субстрата в пользу реакции. [ 6 ] : 158
Кислотно-основной катализ
[ редактировать ]В некоторых реакциях протоны и гидроксиды могут непосредственно действовать как кислота и основание с точки зрения специфического кислотного и специфического основного катализа. Но чаще группы в субстрате и активном центре действуют как кислота и основание Бренстеда-Лоури. Это называется общей теорией кислот и общих оснований. Самый простой способ отличить их — проверить, определяется ли скорость реакции концентрациями общей кислоты и основания. Если ответ положительный, то реакция общего типа. Поскольку оптимальное значение pH большинства ферментов составляет от 6 до 7, аминокислоты в боковой цепи обычно имеют p K a от 4 до 10. Кандидаты включают аспартат , глутамат , гистидин , цистеин . Эти кислоты и основания могут стабилизировать нуклеофил или электрофил, образующийся в ходе катализа, создавая положительные и отрицательные заряды. [ 6 ] : 164–70
Конформационное искажение
[ редактировать ]Количественные исследования ферментативных реакций часто обнаруживают, что ускорение скорости химической реакции не может быть полностью объяснено существующими теориями, такими как приближение, кислотно-основной катализ и электрофильно-нуклеофильный катализ. И здесь возникает очевидный парадокс: в обратимой ферментативной реакции, если активный центр идеально соответствует субстрату, обратная реакция будет замедлена, поскольку продукты не могут идеально вписаться в активный центр. Таким образом, было введено конформационное искажение, утверждающее, что и активный центр, и субстрат могут претерпевать конформационные изменения, чтобы постоянно соответствовать друг другу. [ 6 ] : 170–5
Предварительно организованная комплементарность активного сайта переходному состоянию
[ редактировать ]Эта теория немного похожа на теорию замка и ключа, но в этот раз активный центр заранее запрограммирован на идеальное связывание с субстратом в переходном состоянии, а не в основном состоянии. Формирование переходного состояния внутри раствора требует большого количества энергии для перемещения молекул растворителя, и реакция замедляется. Таким образом, активный центр может заменять молекулы растворителя и окружать подложки, чтобы минимизировать контрпродуктивный эффект, вызываемый раствором. Наличие заряженных групп в активном центре будет притягивать субстраты и обеспечивать электростатическую комплементарность. [ 6 ] : 176–8
Примеры механизмов ферментативного катализа
[ редактировать ]В действительности большинство ферментативных механизмов включают комбинацию нескольких различных типов катализа.

Роль глутатиона (GSH) заключается в удалении накопленных активных форм кислорода, которые могут повредить клетки. Во время этого процесса его тиоловая боковая цепь окисляется , и две молекулы глутатиона соединяются дисульфидной связью с образованием димера (GSSG). Для регенерации глутатиона необходимо разорвать дисульфидную связь. В клетках человека это делается с помощью глутатионредуктазы (GR). [ нужна ссылка ]
Глутатионредуктаза представляет собой димер, содержащий две идентичные субъединицы. требуется один НАДФ и один ФАД В качестве кофакторов . Активный сайт расположен в месте связи между двумя субъединицами. НАДФН участвует в образовании ФАДН-. В активном центре, помимо кофактора FAD, имеются два остатка цистеина , которые используются для разрыва дисульфидной связи во время каталитической реакции. НАДФН связан тремя положительно заряженными остатками: Arg-218, His-219 и Arg-224. [ нужна ссылка ]
Каталитический процесс начинается, когда ФАД восстанавливается НАДФН до принятия одного электрона и от ФАДН. − . Затем он атакует дисульфидную связь, образованную между двумя остатками цистеина, образуя одну связь SH и одну связь S. − группа. Это С − группа будет действовать как нуклеофил, атакуя дисульфидную связь в окисленном глутатионе (GSSG), разрывая ее и образуя комплекс цистеин-SG. Первый СГ − анион высвобождается и затем получает один протон от соседней SH-группы и от первого мономера глутатиона. Далее соседний S − группа атакует дисульфидную связь в комплексе цистеин-SG и высвобождает второй SG. − анион. Он принимает один протон в растворе и образует второй мономер глутатиона.
[ 1 ] : 137–9
Химотрипсин
[ редактировать ]
Химотрипсин представляет собой сериновую эндопептидазу которая присутствует в соке поджелудочной железы и способствует гидролизу белков , и пептидов . [ 1 ] : 84–6 Катализирует гидролиз пептидных связей в -изомерах тирозина фенилаланина , L и триптофана . В активном центре этого фермента три аминокислотных остатка работают вместе, образуя каталитическую триаду , составляющую каталитический центр. В химотрипсине такими остатками являются Ser-195, His-57 и Asp-102.
Механизм действия химотрипсина можно разделить на две фазы. Во-первых, Ser-195 нуклеофильно атакует углерод пептидной связи в субстрате с образованием тетраэдрического промежуточного соединения. Нуклеофильность Ser-195 усиливается His-57, который отрывает протон от Ser-195 и, в свою очередь, стабилизируется отрицательно заряженной карбоксилатной группой (RCOO − ) в Асп-102. Кроме того, тетраэдрический промежуточный оксианион, образующийся на этом этапе, стабилизируется водородными связями Ser-195 и Gly-193.
На втором этапе группа R'NH протонируется His-57 с образованием R'NH 2 и покидает промежуточное соединение, оставляя после себя ацилированный Ser-195. Затем His-57 снова действует как основание, отрывая один протон от молекулы воды. Образующийся гидроксид- анион нуклеофильно атакует ацил-ферментный комплекс с образованием второго тетраэдрического промежуточного оксианиона, который снова стабилизируется H-связями. В конце концов, Ser-195 покидает тетраэдрический интермедиат, разрывая связь CO, которая соединяла фермент с пептидным субстратом. Протон передается к Ser-195 через His-57, так что все три аминокислоты возвращаются в исходное состояние.
Развязывание
[ редактировать ]На развязывание субстрата влияют различные факторы. Более крупные лиганды обычно дольше остаются в активном центре. [ 12 ] как и те, у кого больше вращающихся связей (хотя это может быть побочным эффектом размера). [ 13 ] Когда растворитель исключен из активного центра, менее гибкие белки приводят к увеличению времени пребывания . Большее количество водородных связей, защищенных от растворителя, также уменьшает разъединение. [ 12 ]
Кофакторы
[ редактировать ]
Ферменты могут использовать кофакторы в качестве «молекул-помощников». Коэнзимами называют те небелковые молекулы, которые связываются с ферментами, помогая им выполнять свою работу. В основном они связаны с активным центром нековалентными связями, такими как водородная связь или гидрофобное взаимодействие . Но иногда между ними может образоваться и ковалентная связь. Например, гем в цитохроме С связан с белком посредством тиоэфирной связи . В некоторых случаях коферменты могут покидать ферменты после завершения реакции. В противном случае они навсегда связываются с ферментом. [ 6 ] : 69 Коэнзим – это широкое понятие, включающее ионы металлов, различные витамины и АТФ . Если ферменту для работы необходим кофермент, его называют апоферментом. Фактически, он сам по себе не может должным образом катализировать реакции. Только когда его кофактор входит и связывается с активным сайтом, образуя холофермент, он работает должным образом.
Одним из примеров кофермента является флавин . Он содержит отдельную конъюгированную кольцевую систему изоаллоксазина. Флавин имеет несколько окислительно-восстановительных состояний и может использоваться в процессах, включающих перенос одного или двух электронов. Он может действовать как акцептор электронов в реакции, такой как окисление НАД до НАДН, принимая два электрона и образуя 1,5-дигидрофлавин. С другой стороны, он может образовывать семихинон ( свободный радикал ), принимая один электрон, а затем превращается в полностью восстановленную форму путем добавления дополнительного электрона. Это свойство позволяет использовать его в процессе одноэлектронного окисления.
Ингибиторы
[ редактировать ]Ингибиторы нарушают взаимодействие фермента и субстрата, замедляя скорость реакции. Существуют различные типы ингибиторов, включая как обратимые, так и необратимые формы.
Конкурентные ингибиторы — это ингибиторы, которые нацелены только на свободные молекулы ферментов. Они конкурируют с субстратами за свободный акцептор фермента и могут быть преодолены путем увеличения концентрации субстрата. У них есть два механизма. Конкурентные ингибиторы обычно имеют структурное сходство с субстратами и/или комплексами ES. В результате они могут вписаться в активный сайт и вызвать благоприятные взаимодействия, заполняя пространство и блокируя проникновение субстратов. Они также могут вызывать временные конформационные изменения в активном центре, из-за чего субстраты не могут идеально совпадать с ним. Через короткий период времени конкурентные ингибиторы исчезнут, оставив фермент нетронутым.
Ингибиторы классифицируются как неконкурентные ингибиторы, если они связывают как свободный фермент, так и комплекс ES. Поскольку они не конкурируют с субстратами за активный центр, их невозможно преодолеть простым увеличением концентрации субстрата. Обычно они связываются с другим участком фермента и изменяют трехмерную структуру активного сайта, блокируя вход или выход субстратов из фермента.
Необратимые ингибиторы аналогичны конкурентным ингибиторам, поскольку оба они связываются с активным центром. Однако необратимые ингибиторы образуют необратимые ковалентные связи с аминокислотными остатками в активном центре и никогда не покидают его. Следовательно, активный центр занят и субстрат не может войти. Иногда ингибитор уходит, но форма каталитического центра постоянно меняется. Эти ингибиторы обычно содержат электрофильные группы, такие как заменители галогенов и эпоксиды . С течением времени все больше и больше ферментов связываются необратимыми ингибиторами и больше не могут функционировать.
| Пример | Привязывает активный сайт? | Снижает скорость реакции? | |
|---|---|---|---|
| Конкурентный обратимый ингибитор | Ингибиторы протеазы ВИЧ | Да | Да |
| Неконкурентный обратимый ингибитор | Тяжелые металлы, такие как свинец и ртуть | Нет | Да |
| Необратимый ингибитор | Цианид | Да | Да |
Примеры конкурентных и необратимых ингибиторов ферментов
[ редактировать ]Конкурентный ингибитор: ингибитор протеазы ВИЧ.
[ редактировать ]
Ингибиторы протеазы ВИЧ используются для лечения пациентов с вирусом СПИДа путем предотвращения репликации его ДНК . Протеаза ВИЧ используется вирусом для расщепления полипротеина Gag-Pol на 3 более мелких белка, которые отвечают за сборку, упаковку и созревание вириона. Этот фермент нацелен на специфический сайт расщепления фенилаланин - пролин в целевом белке. [ 14 ] Если протеаза ВИЧ отключена, частица вириона потеряет функцию и не сможет заразить пациентов. Поскольку он необходим для репликации вируса и отсутствует у здорового человека, он является идеальной мишенью для разработки лекарств .
Протеаза ВИЧ принадлежит к семейству аспарагиновых протеаз и имеет аналогичный механизм. Во-первых, остаток аспартата активирует молекулу воды и превращает ее в нуклеофил . Затем он атакует карбонильную группу внутри пептидной связи (NH-CO) с образованием тетраэдрического промежуточного соединения. Атом азота в составе интермедиата принимает протон, образуя амидную группу , и последующая перегруппировка приводит к разрыву связи между ним и интермедиатом и образованию двух продуктов. [ 15 ]
Ингибиторы обычно содержат негидролизуемые гидроксиэтиленовые или гидроксиэтиламиновые группы, которые имитируют тетраэдрическое промежуточное соединение. Поскольку они имеют аналогичную структуру и электростатическое расположение, что и переходное состояние субстратов, они все еще могут помещаться в активный центр, но не могут быть разрушены, поэтому гидролиз не может произойти.
Неконкурентный ингибитор: стрихнин.
[ редактировать ]Стрихнин — это нейротоксин , вызывающий смерть, поражая нервы, контролирующие мышечные сокращения , и вызывая затруднение дыхания. Импульс передается между синапсами через нейромедиатор , называемый ацетилхолином . Он высвобождается в синапс между нервными клетками и связывается с рецепторами постсинаптической клетки. Затем потенциал действия генерируется , который передается через постсинаптическую клетку, чтобы начать новый цикл.
Глицин может ингибировать активность рецепторов нейромедиаторов, поэтому для запуска потенциала действия требуется большее количество ацетилхолинэстеразы. Это гарантирует, что генерация нервных импульсов строго контролируется. Однако этот контроль нарушается при добавлении стрихнина. Он ингибирует глициновые рецепторы ( хлоридный канал ), и гораздо более низкий уровень концентрации нейромедиатора может вызвать потенциал действия. Нервы теперь постоянно передают сигналы и вызывают чрезмерное сокращение мышц, что приводит к удушью и смерти. [ 16 ]
Необратимый ингибитор: диизопропилфторфосфат.
[ редактировать ]
Диизопропилфторфосфат (ДИФП) — необратимый ингибитор, блокирующий действие сериновой протеазы . Когда он связывается с ферментом, нуклеофильного замещения происходит реакция фторида водорода , в результате которой высвобождается одна молекула . Группа ОН в активном центре действует как нуклеофил, атакуя фосфор в DIFP, образуя тетраэдрический промежуточный продукт и высвобождая протон. Затем связь PF разрывается, один электрон передается атому F, и он покидает интермедиат как F. − анион. Он соединяется с протоном в растворе, образуя одну молекулу HF. Между активным центром и DIFP образуется ковалентная связь, поэтому боковая цепь серина больше не доступна для субстрата. [ 17 ]
В открытии лекарств
[ редактировать ]Идентификация активных центров имеет решающее значение в процессе открытия лекарств . Трехмерная структура фермента анализируется для выявления остатков активного центра и разработки лекарств, которые могут в них вписаться. Протеолитические ферменты являются мишенью для некоторых лекарств, например, ингибиторов протеаз, к которым относятся препараты против СПИДа и гипертонии. [ 18 ] Эти ингибиторы протеазы связываются с активным центром фермента и блокируют взаимодействие с природными субстратами. [ 19 ] Важным фактором при разработке лекарств является сила связывания между активным центром и ингибитором фермента. [ 20 ] Если фермент, обнаруженный в бактериях, значительно отличается от человеческого фермента, то можно разработать ингибитор против этой конкретной бактерии, не нанося вреда человеческому ферменту. Если один вид фермента присутствует только в одном виде организма, его ингибитор можно использовать для целенаправленного уничтожения.
Активные сайты могут быть картированы, чтобы помочь в разработке новых лекарств, таких как ингибиторы ферментов. Это включает в себя описание размера активного сайта, а также количества и свойств подсайтов, таких как подробности взаимодействия привязки. [ 18 ] Однако современная технология баз данных, называемая CPASS (Сравнение структур активных сайтов белков), позволяет более детально сравнивать активные сайты и находить структурное сходство с помощью программного обеспечения. [ 21 ]
Применение ингибиторов ферментов
[ редактировать ]| Пример | Механизм действия | |
|---|---|---|
| Антибактериальное средство | Пенициллин | бактерий Клеточная стенка состоит из пептидогликана . Во время роста бактерий существующая сшивка пептидогликановых волокон нарушается, поэтому новый мономер клеточной стенки может быть интегрирован в клеточную стенку. Пенициллин действует путем ингибирования транспептидазы , которая необходима для образования поперечных связей, поэтому клеточная стенка ослабляется и разрывается из-за тургорного давления . |
| Противогрибковое средство | Азол | Эргостерин – это стерин , который образует поверхностную мембрану клеток грибов . Азол может ингибировать свой биосинтез, ингибируя альфа-деметилазу ланостерола 14 , поэтому новый эргостерин не вырабатывается, а вредный 14α-ланостерин накапливается внутри клетки. Кроме того, азол может генерировать активные формы кислорода . |
| Противовирусное средство | Саквинавир | Протеаза ВИЧ необходима для расщепления полипротеина Gag-Pol на 3 отдельных белка, чтобы они могли правильно функционировать и начать процесс упаковки вируса. Ингибиторы протеазы ВИЧ, такие как саквинавир, ингибируют ее, поэтому новые зрелые вирусные частицы не могут образовываться. |
| Инсектициды | Физостигмин | В нервной системе животных ацетилхолинэстераза необходима для расщепления нейромедиатора ацетилхолина на ацетат и холин . Физостигмин связывается с его активным центром и ингибирует его, поэтому импульсный сигнал не может передаваться по нервам. Это приводит к гибели насекомых, поскольку они теряют контроль над работой мышц и сердца. |
| Гербициды | Циклогександион | Циклогександион воздействует на ацетил-КоА-карбоксилазу , которая участвует в первом этапе синтеза жиров: АТФ-зависимом карбоксилировании ацетил -КоА в малонил-КоА . Липиды играют важную роль в формировании клеточной мембраны. |
Аллостерические сайты
[ редактировать ]
Аллостерический сайт — это участок фермента, не связанный с его активным центром, который может связывать эффекторную молекулу. Это взаимодействие является еще одним механизмом регуляции ферментов. Аллостерическая модификация обычно происходит в белках, содержащих более одной субъединицы. Аллостерические взаимодействия часто присутствуют в метаболических путях и полезны, поскольку позволяют одному этапу реакции регулировать другой этап. [ 19 ] Они позволяют ферменту иметь ряд молекулярных взаимодействий, помимо высокоспецифичного активного центра. [ 19 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Багг ТД (2004). Введение в химию ферментов и коферментов (PDF) (2-е изд.). Блэквелл Паблишинг Лимитед. ISBN 9781405114523 . Архивировано из оригинала (PDF) 22 марта 2018 года.
- ^ Jump up to: а б Шанмугам С (2009). Ферментная технология . ИК Международный издательский дом. п. 48. ИСБН 9789380026053 .
- ^ Jump up to: а б Правда Л., Берка К., Свободова Варжекова Р. и др. (2014). «Анатомия ферментных каналов» . БМК Биоинформатика . 15 (1): 379. doi : 10.1186/s12859-014-0379-x . ПМЦ 4245731 . ПМИД 25403510 .
- ^ Альбертс Б. (2010). Основная клеточная биология . Гирляндная наука. п. 91. ИСБН 9780815341291 .
- ^ Jump up to: а б Дагмар Р., Грегори А. (2008). «Как работают ферменты». Наука . 320 (5882): 1428–1429. дои : 10.1126/science.1159747 . ПМИД 18556536 . S2CID 43617575 .
- ^ Jump up to: а б с д и ж г час я Роберт А. (2000). Ферменты: практическое введение в структуру, механизм и анализ данных (PDF) (2-е изд.). Уайли-Блэквелл. ISBN 9780471359296 .
- ^ Кул ET (1984). «Плотность активного центра и соответствие субстрата репликации ДНК». Ежегодный обзор биохимии . 71 : 191–219. doi : 10.1146/annurev.biochem.71.110601.135453 . ПМИД 12045095 .
- ^ Чермели, Питер; Палотаи, Робин; Нусинов, Рут (2010). «Индуцированная подгонка, конформационный отбор и независимые динамические сегменты: расширенный взгляд на события связывания» . Тенденции биохимических наук . 35 (10): 539–546. arXiv : 1005.0348 . дои : 10.1016/j.tibs.2010.04.009 . ISSN 0968-0004 . ПМК 3018770 . ПМИД 20541943 .
- ^ Дэниел Э (1995). «Теория ключа-замка и теория вынужденного соответствия». Angewandte Chemie, международное издание . 33 (2324): 2375–2378. дои : 10.1002/anie.199423751 .
- ^ Салливан С.М. (2008). «Ферменты с закрытыми активными центрами должны действовать по механизму индуцированной подгонки, а не по конформационному отбору» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (37): 13829–13834. Бибкод : 2008PNAS..10513829S . дои : 10.1073/pnas.0805364105 . ПМЦ 2544539 . ПМИД 18772387 .
- ^ Коупленд, Роберт А. (2013). «Лекарственное средство – целевое время пребывания». Оценка ингибиторов ферментов при открытии лекарств . John Wiley & Sons, Ltd., стр. 287–344. ISBN 978-1-118-54039-8 .
- ^ Jump up to: а б Пан, Альберт К.; Борхани, Дэвид В.; Дрор, Рон О.; Шоу, Дэвид Э. (2013). «Молекулярные детерминанты кинетики связывания лекарства с рецептором». Открытие наркотиков сегодня . 18 (13–14): 667–673. дои : 10.1016/j.drudis.2013.02.007 . ISSN 1359-6446 . ПМИД 23454741 .
- ^ Миллер, Дункан С.; Ланн, Грэм; Джонс, Питер; Сабнис, Йогеш; Дэвис, Никола Л.; Дрисколл, Пол (2012). «Исследование влияния молекулярных свойств на кинетику связывания лиганда с его биологической мишенью». МедХимКомм . 3 (4): 449–452. дои : 10.1039/c2md00270a . ISSN 2040-2503 .
- ^ Флекснер С (1998). «Ингибиторы ВИЧ-протеазы». Медицинский журнал Новой Англии . 338 (18): 1281–1292. дои : 10.1056/NEJM199804303381808 . ПМИД 9562584 .
- ^ Ашраф Б., Чи-Хьюи В. (2003). «Протеаза ВИЧ-1: механизм и открытие лекарства». Органическая и биомолекулярная химия . 1 (1): 5–14. дои : 10.1039/B208248A . ПМИД 12929379 .
- ^ Грей В., Рик Дж. (1993). «Цитопротекция путем ингибирования хлоридных каналов: механизм действия глицина и стрихнина». Науки о жизни . 53 (15): 1211–1215. дои : 10.1016/0024-3205(93)90539-F . ПМИД 8412478 .
- ^ Янсен Э.Ф., Нуттиг Ф., Боллс АК (1949). «Способ ингибирования химотрипсина диизопропилфторфосфатом; введение фосфора» . Журнал биологической химии . 179 (1): 201–204. дои : 10.1016/S0021-9258(18)56828-9 . ПМИД 18119235 .
- ^ Jump up to: а б Шехтер I (2005). «Картирование активного центра протеаз в 1960-х годах и рациональный дизайн ингибиторов/лекарств в 1990-х годах». Современная наука о белках и пептидах . 6 (6): 501–512. дои : 10.2174/138920305774933286 . ПМИД 16381600 .
- ^ Jump up to: а б с ДеДекер Б.С. (2000). «Аллостерические препараты: мышление за пределами поля активного центра» . Химия и биология . 7 (5): 103–107. дои : 10.1016/S1074-5521(00)00115-0 . ПМИД 10801477 .
- ^ Цурхер М (2008). «Структурно-ориентированный дизайн лекарств: изучение правильного заполнения аполярных карманов в активных центрах ферментов». Журнал органической химии . 73 (12): 4345–4361. дои : 10.1021/jo800527n . ПМИД 18510366 .
- ^ Пауэрс Р. (2006). «Сравнение структур активных центров белков для функциональной аннотации белков и разработки лекарств» . Белки . 65 (1): 124–135. дои : 10.1002/prot.21092 . ПМИД 16862592 . S2CID 2527166 .
Дальнейшее чтение
[ редактировать ]- Алан Фершт, Структура и механизм в науке о белках: Руководство по ферментативному катализу и сворачиванию белков. У. Х. Фриман, 1998. ISBN 0-7167-3268-8
- Багг, Т. Введение в химию ферментов и коферментов. (2-е издание), Blackwell Publishing Limited, 2004 г. ISBN 1-4051-1452-5 .
| Базы данных органов управления : Национальные |
|---|