Цитохром с
| ЦИКС | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CYCS , CYC, HCS, THC4, цитохром с, соматический, цитохром с, цит с | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 123970 ; МГИ : 88578 ; Гомологен : 133055 ; Генные карты : CYCS ; ОМА : CYCS — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





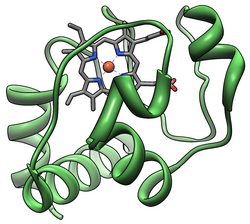
Цитохромный комплекс , или cyt c , представляет собой небольшой гемопротеин , слабо связанный с внутренней мембраной митохондрии , где он играет решающую роль в клеточном дыхании . Он переносит электроны между комплексами III (коэнзим Q – Cyt c-редуктаза) и IV (Cyt c-оксидаза). Цитохром С хорошо растворим в воде , в отличие от других цитохромов . Он способен подвергаться окислению и восстановлению , поскольку его атом железа преобразуется между двухвалентной и трехвалентной формами, но не связывает кислород . Он также играет важную роль в апоптозе клеток . У человека цитохром с кодируется CYCS геном . [ 5 ] [ 6 ]
Распространение видов
[ редактировать ]Цитохром С — высококонсервативный белок у всего спектра эукариотических видов, обнаруженный у растений, животных, грибов и многих одноклеточных организмов. Это, наряду с небольшими размерами (молекулярная масса около 12 000 дальтон ), [ 7 ] делает его полезным в исследованиях кладистики . [ 8 ] Цитохром С изучался с целью дать представление об эволюционной биологии.
Цитохром С имеет первичную структуру, состоящую из цепочки примерно из 100 аминокислот . Многие организмы высшего порядка обладают цепью из 104 аминокислот. [ 9 ] Последовательность цитохрома с у человека идентична таковой у шимпанзе (наших ближайших родственников), но отличается от таковой у лошадей. [ 10 ]
Цитохром С имеет аминокислотную последовательность, которая высоко консервативна у эукариот и варьируется всего на несколько остатков. У более чем тридцати видов, протестированных в одном исследовании, 34 из 104 аминокислот были консервативными (идентичными в своем характерном положении). [ 11 ] Например, цитохромоксидаза человека взаимодействовала с цитохромом с пшеницы in vitro ; что справедливо для всех пар протестированных видов. [ 11 ] Кроме того, окислительно-восстановительный потенциал +0,25 вольт одинаков во всех цитохрома с . изученных молекулах [ 11 ]
Структура
[ редактировать ]
Цитохром с принадлежит к классу I семейства цитохромов с-типа. [ 13 ] и содержит характерный аминокислотный мотив CXXCH (цистеин-любой-любой-цистеин-гистидин), который связывает гем. [ 14 ] Этот мотив расположен ближе к N-концу цепи пептидной и содержит гистидин в качестве 5-го лиганда железа гема. Шестой лиганд представлен остатком метионина , расположенным ближе к С-концу . Основная цепь белка свернута в пять α-спиралей , которые пронумерованы α1-α5 от N-конца до С-конца. Спирали α3, α4 и α5 называются спиралями 50, 60 и 70 соответственно, когда речь идет о митохондриальном цитохроме с. [ 15 ]
Гем с
[ редактировать ]
В то время как большинство гемовых белков прикрепляются к простетической группе посредством лигирования ионов железа и третичных взаимодействий, гемовая группа цитохрома с образует тиоэфирные связи с двумя боковыми цепями цистеина белка. [ 16 ] Одним из основных свойств гема с, позволяющих цитохрому с выполнять разнообразные функции, является его способность иметь в природе разные восстановительные потенциалы. Это свойство определяет кинетику и термодинамику реакции переноса электрона. [ 17 ]
Дипольный момент
[ редактировать ]Дипольный момент играет важную роль в ориентации белков в нужных направлениях и повышении их способности связываться с другими молекулами. [ 18 ] [ 19 ] Дипольный момент цитохрома с возникает в результате кластера отрицательно заряженных боковых цепей аминокислот на «спине» фермента. [ 19 ] Несмотря на вариации числа связанных гемовых групп и вариации последовательности, дипольный момент цитохромов с позвоночных удивительно консервативен. Например, все цитохромы с позвоночных имеют дипольный момент примерно 320 дебай , тогда как цитохромы с растений и насекомых имеют дипольный момент примерно 340 дебай. [ 19 ]
Функция
[ редактировать ]Электронно-транспортная цепь
[ редактировать ]Цитохром С является важным компонентом дыхательной цепи транспорта электронов в митохондриях. Гемовая комплекса группа цитохрома с принимает электроны от bc 1 III и транспортирует их к комплексу IV , а сам передает энергию в противоположном направлении.
Цитохром С также может катализировать некоторые окислительно-восстановительные реакции, такие как гидроксилирование и ароматическое окисление , и проявляет пероксидазную активность за счет окисления различных доноров электронов, таких как 2,2-азинобис ( 3-этилбензтиазолин-6-сульфоновая кислота) ( ABTS ), 2- кето-4-тиометилмасляная кислота и 4-аминоантипирин.
Бактериальный цитохром С действует как нитритредуктаза . [ 20 ]
Роль в апоптозе
[ редактировать ]также обнаружил, что цитохром С В 1996 году Сяодун Ван играет промежуточную роль в апоптозе , контролируемой форме гибели клеток, используемой для уничтожения клеток в процессе развития или в ответ на инфекцию или повреждение ДНК. [ 21 ]
Цитохром С связывается с кардиолипином во внутренней митохондриальной мембране, тем самым закрепляя его присутствие и не позволяя ему высвободиться из митохондрий и инициировать апоптоз. Хотя первоначальное притяжение между кардиолипином и цитохромом с является электростатическим из-за чрезвычайно положительного заряда цитохрома с, окончательное взаимодействие является гидрофобным, когда гидрофобный хвост кардиолипина вставляется в гидрофобную часть цитохрома с.
Во время ранней фазы апоптоза стимулируется продукция митохондриальных АФК и окисляется кардиолипин за счет пероксидазной функции комплекса кардиолипин-цитохром с. Затем гемопротеин отделяется от внутренней мембраны митохондрий и может быть вытеснен в растворимую цитоплазму через поры внешней мембраны. [ 22 ]
Устойчивое повышение уровня кальция предшествует высвобождению цитоцита С из митохондрий. Высвобождение небольшого количества цитоцита c приводит к взаимодействию с рецептором IP3 (IP3R) на эндоплазматическом ретикулуме (ЭР), вызывая высвобождение кальция из ЭР. Общее увеличение содержания кальция вызывает массовое высвобождение cyt c , который затем действует в петле положительной обратной связи, поддерживая высвобождение кальция из ER через IP3R. [ 23 ] Это объясняет, как высвобождение кальция из ЭР может достигать цитотоксического уровня. Это высвобождение цитохрома с, в свою очередь, активирует каспазу 9 , цистеиновую протеазу . Затем каспаза 9 может активировать каспазу 3 и каспазу 7 , которые отвечают за разрушение клетки изнутри.
Ингибирование апоптоза
[ редактировать ]Одним из способов активации апоптоза клеток является высвобождение цитохрома с из митохондрий в цитозоль. Исследование показало, что клетки способны защищаться от апоптоза, блокируя высвобождение цитохрома с с помощью Bcl- L. x [ 24 ] Другой способ, с помощью которого клетки могут контролировать апоптоз, — это фосфорилирование Tyr48, которое превращает цитохром c в антиапоптотический переключатель. [ 25 ]
В качестве антиоксидантного фермента
[ редактировать ]
В дополнение к своей хорошо известной роли в цепи переноса электронов и апоптозе клеток, согласно недавнему исследованию, цитохром С также может действовать как антиоксидантный фермент в митохондриях; он делает это путем удаления супероксида ( O - 2 ) и перекись водорода (H 2 O 2 ) из митохондрий . [ 26 ] Следовательно, цитохром С необходим не только в митохондриях для клеточного дыхания, но также необходим в митохондриях для ограничения производства О - 2 и H2O2 O2. [ 26 ]
Экстрамитохондриальная локализация
[ редактировать ]Широко распространено мнение, что цитохром с в нормальных физиологических условиях локализуется исключительно в митохондриальном межмембранном пространстве. [ 27 ] Считается, что высвобождение цитохрома с из митохондрий в цитозоль, где он активирует каспаз семейство протеаз , является основным триггером, приводящим к началу апоптоза. [ 28 ] Измерение количества цитохрома с, просачивающегося из митохондрий в цитозоль и из клетки в культуральную среду, является чувствительным методом мониторинга степени апоптоза. [ 29 ] [ 30 ] Однако подробные иммуноэлектронно-микроскопические исследования срезов тканей крыс с использованием антител, специфичных к цитохрому с, предоставили убедительные доказательства того, что цитохром с в нормальных клеточных условиях также присутствует во внемитохондриальных местах. [ 31 ] В ацинарных клетках поджелудочной железы и передней доле гипофиза сильное и специфическое присутствие цитохрома с было обнаружено в гранулах зимогена и в гранулах гормона роста соответственно. В поджелудочной железе цитохром с обнаружен также в конденсирующих вакуолях ацина и в просвете . Показано, что экстрамитохондриальная локализация цитохрома с является специфичной, поскольку она полностью устраняется при адсорбции первичного антитела с очищенным цитохромом с. [ 31 ] Помимо цитохрома с, экстрамитохондриальная локализация наблюдалась и для большого числа других белков, в том числе кодируемых митохондриальной ДНК. [ 32 ] [ 33 ] [ 34 ] Это повышает вероятность существования еще не выявленных специфических механизмов транслокации белков из митохондрий в другие клеточные направления. [ 34 ] [ 35 ]
Приложения
[ редактировать ]Обнаружение супероксида
[ редактировать ]
Цитохром с использовался для обнаружения производства пероксида в биологических системах. По мере образования супероксида количество окисленного цитохрома с 3+ увеличивается, а снижается цитохром с 2+ уменьшается. [ 36 ] Однако супероксид часто получают из оксида азота. В присутствии оксида азота происходит восстановление цитохрома с 3+ тормозится. [ 37 ] Это приводит к окислению цитохрома с. 2+ к цитохрому с 3+ , пероксиазотистой кислотой промежуточным продуктом реакции оксида азота и супероксида. [ 37 ] Присутствие пероксинитрита или H 2 O 2 и диоксида азота NO 2 в митохондриях может быть смертельным, поскольку они нитрируют тирозиновые остатки цитохрома с, что приводит к нарушению функции цитохрома с как переносчика электронов в электрон-транспортной цепи. [ 38 ]
В качестве фермента каталитической активности
[ редактировать ]Цитохром С также широко изучался как фермент с пероксидазоподобной активностью. Цитохром C был конъюгирован с заряженным полимером для проверки его пероксидазоподобной активности. [ 39 ] [ 40 ] Вдохновленный естественными примерами инкапсуляции ферментов в белковых клеточных структурах (пример: карбоксисомы, ферритин и энкапсулин), цитохром C был инкапсулирован в небольшой самособирающийся ДНК-связывающий белок размером 9 нм из белковой клетки клеток, голодающих по питательным веществам (Dps), с использованием химерной самоорганизации. -сборочный подход. Авторы наблюдали уникальное поведение каталитической активности при инкапсуляции фермента внутри белковой клетки, которое отличалось от активности фермента в растворе. Это было связано с локальным микроокружением, обеспечиваемым внутренней полостью наноклетки Dps, которая отличается от основной. [ 41 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000172115 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000063694 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ген Энтрез: цитохром с» .
- ^ Тафани М., Карпинич Н.О., Херстер К.А., Пасторино Дж.Г., Шнайдер Т., Руссо М.А. и др. (март 2002 г.). «Высвобождение цитохрома с при активации рецептора Fas зависит от транслокации полноразмерного бида и индукции перехода митохондриальной проницаемости» . Журнал биологической химии . 277 (12): 10073–82. дои : 10.1074/jbc.M111350200 . ПМИД 11790791 .
- ^ «Цитохром С – Homo sapiens (Человек)» . Р99999 . Консорциум ЮниПрот.
масса 11749 Дальтон.
- ^ Марголиаш Э. (октябрь 1963 г.). «Первичная структура и эволюция цитохрома с» . Труды Национальной академии наук Соединенных Штатов Америки . 50 (4): 672–9. Бибкод : 1963ПНАС...50..672М . дои : 10.1073/pnas.50.4.672 . ПМК 221244 . ПМИД 14077496 .
- ^ «Аминокислотные последовательности в белках цитохрома с разных видов» (PDF) . Архивировано из оригинала (PDF) 28 декабря 2013 г. , адаптировано из Стралер А.Н. (1999). Наука и история Земли: споры об эволюции и сотворении мира . Амхерст, Нью-Йорк: Книги Прометея. п. 348. ИСБН 978-1-57392-717-8 .
- ^ Луркин П.Ф., Стоун Л., Кавалли-Сфорца Л.Л. (2007). Гены, культура и эволюция человека: синтез . Оксфорд: Блэквелл. п. 79. ИСБН 978-1-4051-5089-7 .
- ^ Перейти обратно: а б с Страйер Л. (1975). Биохимия (1-е изд.). Сан-Франциско: WH Freeman and Company. п. 362 . ISBN 978-0-7167-0174-3 .
- ^ Макферсон А., ДеЛукас Л.Дж. (2015). «Кристаллизация белков в условиях микрогравитации» . npj Микрогравитация . 1 : 15010. doi : 10.1038/npjmgrad.2015.10 . ПМК 5515504 . ПМИД 28725714 .
- ^ Эмблер Р.П. (май 1991 г.). «Изменчивость последовательности бактериальных цитохромов с». Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1058 (1): 42–7. дои : 10.1016/S0005-2728(05)80266-X . ПМИД 1646017 .
- ^ Мавриду Д.А., Фергюсон С.Дж., Стивенс Дж.М. (март 2013 г.). «Сборка цитохрома С». ИУБМБ Жизнь . 65 (3): 209–16. дои : 10.1002/iub.1123 . ПМИД 23341334 . S2CID 32216217 .
- ^ Лю Дж., Чакраборти С., Хоссейнзаде П., Ю Ю, Тиан С., Петрик И. и др. (23 апреля 2014 г.). «Металлопротеины, содержащие цитохромные, железо-серные или медные окислительно-восстановительные центры» . Химические обзоры . 114 (8): 4366–4469. дои : 10.1021/cr400479b . ISSN 0009-2665 . ПМК 4002152 . ПМИД 24758379 .
- ^ Кан X, Кэри Дж (ноябрь 1999 г.). «Роль гема в структурной организации цитохрома с, исследованная методом полусинтеза». Биохимия . 38 (48): 15944–51. дои : 10.1021/bi9919089 . ПМИД 10625461 .
- ^ Чжао Ю, Ван ЗБ, Сюй JX (январь 2003 г.). «Влияние цитохрома с на образование и выведение О 2 – и H 2 O 2 в митохондриях» . Журнал биологической химии . 278 (4): 2356–60. doi : 10.1074/jbc.M209681200 . PMID 12435729 .
- ^ Коппенол WH, Марголиаш Э (апрель 1982 г.). «Асимметричное распределение зарядов на поверхности цитохрома с лошади. Функциональное значение» . Журнал биологической химии . 257 (8): 4426–37. дои : 10.1016/S0021-9258(18)34740-9 . ПМИД 6279635 .
- ^ Перейти обратно: а б с Коппенол WH, Раш Дж.Д., Миллс Дж.Д., Марголиаш Э. (июль 1991 г.). «Дипольный момент цитохрома с» . Молекулярная биология и эволюция . 8 (4): 545–58. doi : 10.1093/oxfordjournals.molbev.a040659 . ПМИД 1656165 .
- ^ Шнайдер Дж., Кронек П.М. (2014). «Производство аммиака мультигемовыми цитохромами C». В Kroneck PM, Torres ME (ред.). Металлоориентированная биогеохимия газообразных соединений в окружающей среде . Ионы металлов в науках о жизни. Том. 14. Спрингер. стр. 211–236. дои : 10.1007/978-94-017-9269-1_9 . ISBN 978-94-017-9268-4 . ПМИД 25416396 .
- ^ Лю X, Ким CN, Ян Дж, Джеммерсон Р, Ван X (июль 1996 г.). «Индукция апоптотической программы в бесклеточных экстрактах: потребность в dATP и цитохроме с» . Клетка . 86 (1): 147–57. дои : 10.1016/S0092-8674(00)80085-9 . ПМИД 8689682 . S2CID 12604356 .
- ^ Оррениус С., Животовский Б (сентябрь 2005 г.). «Окисление кардиолипина освобождает цитохром с». Химическая биология природы . 1 (4): 188–9. дои : 10.1038/nchembio0905-188 . ПМИД 16408030 . S2CID 45381495 .
- ^ Боенинг Д., Паттерсон Р.Л., Седагат Л., Глебова Н.О., Куросаки Т., Снайдер Ш.Х. (декабрь 2003 г.). «Цитохром С связывается с инозитол (1,4,5) трифосфатными рецепторами, усиливая кальций-зависимый апоптоз». Природная клеточная биология . 5 (12): 1051–61. дои : 10.1038/ncb1063 . ПМИД 14608362 . S2CID 27761335 .
- ^ Харбанда С., Пандей П., Шофилд Л., Исраэлс С., Ронцинске Р., Йошида К. и др. (июнь 1997 г.). «Роль Bcl-xL как ингибитора цитозольного накопления цитохрома C в апоптозе, вызванном повреждением ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (13): 6939–42. Бибкод : 1997PNAS...94.6939K . дои : 10.1073/pnas.94.13.6939 . ПМК 21263 . ПМИД 9192670 .
- ^ Гарсиа-Эредиа Х.М., Диас-Кинтана А., Сальсано М., Орсаес М., Перес-Пайя Э., Тейшейра М. и др. (декабрь 2011 г.). «Фосфорилирование тирозина превращает щелочной переход в биологически значимый процесс и заставляет человеческий цитохром С вести себя как антиапоптотический переключатель». Журнал биологической неорганической химии . 16 (8): 1155–68. дои : 10.1007/s00775-011-0804-9 . ПМИД 21706253 . S2CID 24156094 .
- ^ Перейти обратно: а б Боуман С.Е., Брен К.Л. (декабрь 2008 г.). «Химия и биохимия гема c: функциональные основы ковалентного присоединения» . Отчеты о натуральных продуктах . 25 (6): 1118–30. дои : 10.1039/b717196j . ПМЦ 2654777 . ПМИД 19030605 .
- ^ Нойперт В. (1997). «Импорт белка в митохондрии». Ежегодный обзор биохимии . 66 : 863–917. doi : 10.1146/annurev.biochem.66.1.863 . ПМИД 9242927 .
- ^ Кремер Г., Даллапорта Б., Реше-Ригон М. (1998). «Регулятор смерти/жизни митохондрий при апоптозе и некрозе». Ежегодный обзор физиологии . 60 : 619–42. doi : 10.1146/annurev.physiol.60.1.619 . ПМИД 9558479 .
- ^ Лоо Дж.Ф., Лау П.М., Хо Х.П., Конг С.К. (октябрь 2013 г.). «Анализ биоштрих-кода на основе аптамера с изотермической амплификацией рекомбиназной полимеразы для обнаружения цитохрома-с и скрининга противораковых препаратов». Таланта . 115 : 159–65. дои : 10.1016/j.talanta.2013.04.051 . ПМИД 24054573 .
- ^ Уотерхаус, Нью-Джерси, Трапани, Дж. А. (июль 2003 г.). «Новый количественный анализ высвобождения цитохрома с в апоптотических клетках» . Смерть клеток и дифференцировка . 10 (7): 853–5. дои : 10.1038/sj.cdd.4401263 . ПМИД 12815469 .
- ^ Перейти обратно: а б Солтис Б.Дж., Эндрюс Д.В., Джеммерсон Р., Гупта Р.С. (2001). «Цитохром-С локализуется в секреторных гранулах поджелудочной железы и передней доли гипофиза». Международная клеточная биология . 25 (4): 331–8. дои : 10.1006/cbir.2000.0651 . ПМИД 11319839 . S2CID 2106599 .
- ^ Гупта Р.С., Рамачандра Н.Б., Боуз Т., Сингх Б. (2008). «Необычное клеточное расположение митохондриальных молекулярных шаперонов Hsp60, Hsp70 и Hsp10». В Чедвик Д., Гуд Дж. (ред.). Биология внеклеточных молекулярных шаперонов . Симпозиумы Фонда Новартис. Том. 291. стр. 59–68, обсуждение 69–73, 137–40. дои : 10.1002/9780470754030.ch5 . ISBN 978-0-470-75403-0 . ПМИД 18575266 .
- ^ Садачаран С.К., Сингх Б., Боуз Т., Гупта Р.С. (ноябрь 2005 г.). «Локализация митохондриальной ДНК, кодирующей субъединицы I и II цитохром с-оксидазы, в гранулах зимогена поджелудочной железы крысы и гранулах гормона роста гипофиза». Гистохимия и клеточная биология . 124 (5): 409–21. дои : 10.1007/s00418-005-0056-2 . ПМИД 16133117 . S2CID 24440427 .
- ^ Перейти обратно: а б Солтыс Б.Дж., Гупта Р.С. (2000). Митохондриальные белки в неожиданных местах клетки: экспорт белков из митохондрий с эволюционной точки зрения . Международный обзор цитологии. Том. 194. стр. 133–96. дои : 10.1016/s0074-7696(08)62396-7 . ISBN 978-0-12-364598-2 . ПМИД 10494626 .
- ^ Солтыс Б.Дж., Гупта Р.С. (май 1999 г.). «Белки митохондриального матрикса в неожиданных местах: они экспортируются?». Тенденции биохимических наук . 24 (5): 174–7. дои : 10.1016/s0968-0004(99)01390-0 . ПМИД 10322429 .
- ^ МакКорд Дж. М., Фридович I (ноябрь 1969 г.). «Супероксиддисмутаза. Ферментативная функция эритрокупреина (гемокупреина)» . Журнал биологической химии . 244 (22): 6049–55. дои : 10.1016/S0021-9258(18)63504-5 . ПМИД 5389100 .
- ^ Перейти обратно: а б Томсон Л., Трухильо М., Теллери Р., Ради Р. (июнь 1995 г.). «Кинетика цитохрома с 2+ окисление пероксинитритом: значение для измерений супероксида в биологических системах, производящих оксид азота». Архивы биохимии и биофизики . 319 (2): 491–7. doi : 10.1006/abbi.1995.1321 . PMID 7786032 .
- ^ Домазу А.С., Гебичка Л., Дидик Дж., Гебицки Дж.Л., ван дер Мейден Б., Коппенол В.Х. (апрель 2014 г.). «Кинетика реакции диоксида азота с цитохромом с железа(II)- и железа(III)». Свободно-радикальная биология и медицина . 69 : 172–80. doi : 10.1016/j.freeradbiomed.2014.01.014 . ПМИД 24447894 .
- ^ Чжан И, Ван Ц, Хесс Х (март 2017 г.). «Увеличение пропускной способности ферментного каскада за счет pH-инженерии микроокружения отдельных ферментов». АКС-катализ . 7 (3): 2047–2051. дои : 10.1021/acscatal.7b01766 .
- ^ Бенсон К.Р., Горецкий Дж., Никифоров А., Цуй В., Каси Р.М., Кумар К.В. (апрель 2019 г.). «Конъюгаты цитохрома с-поли(акриловой кислоты) с улучшенным числом оборота пероксидазы». Органическая и биомолекулярная химия . 17 (16): 4043–4048. дои : 10.1039/c9ob00541b . ПМИД 30950479 .
- ^ Вагвани Х.К., Дуглас Т. (март 2021 г.). «Цитохром С с пероксидазоподобной активностью, инкапсулированный внутри небольшой наноклетки белка DPS» . Журнал химии материалов Б. 9 (14): 3168–3179. дои : 10.1039/d1tb00234a . ПМИД 33885621 .
Дальнейшее чтение
[ редактировать ]- Кумарсвами Р., Чандна С. (февраль 2009 г.). «Предполагаемые партнеры в высвобождении цитохрома-с, опосредованном Bax: ANT, CypD, VDAC или ни один из них?». Митохондрия . 9 (1): 1–8. дои : 10.1016/j.mito.2008.10.003 . ПМИД 18992370 .
- Скулачев В.П. (февраль 1998 г.). «Цитохром С в апоптотическом и антиоксидантном каскадах». Письма ФЭБС . 423 (3): 275–80. дои : 10.1016/S0014-5793(98)00061-1 . ПМИД 9515723 . S2CID 10267410 .
- Маннелла, Калифорния (1998). «Конформационные изменения в белке митохондриального канала, VDAC, и их функциональные последствия». Журнал структурной биологии . 121 (2): 207–18. дои : 10.1006/jsbi.1997.3954 . ПМИД 9615439 .
- Ферри К.Ф., Жакото Э., Бланко Дж., Эсте Х.А., Кремер Г. (2000). «Митохондриальный контроль гибели клеток, индуцированной белками, кодируемыми ВИЧ-1». Анналы Нью-Йоркской академии наук . 926 (1): 149–64. Бибкод : 2000NYASA.926..149F . дои : 10.1111/j.1749-6632.2000.tb05609.x . ПМИД 11193032 . S2CID 21997163 .
- Бриттон Р.С., Лестер К.Л., Бэкон Б.Р. (октябрь 2002 г.). «Токсичность железа и хелатная терапия». Международный журнал гематологии . 76 (3): 219–28. дои : 10.1007/BF02982791 . ПМИД 12416732 . S2CID 22572183 .
- Хайдер Н., Нарула Н., Нарула Дж. (декабрь 2002 г.). «Апоптоз при сердечной недостаточности представляет собой запрограммированное выживание клеток, а не смерть кардиомиоцитов и вероятность обратного ремоделирования». Журнал сердечной недостаточности . 8 (6 Дополнение): S512–7. дои : 10.1054/jcaf.2002.130034 . ПМИД 12555167 .
- Кастедо М., Перфеттини Дж.Л., Андрео К., Румье Т., Пьячентини М., Кремер Г. (декабрь 2003 г.). «Митохондриальный апоптоз, индуцированный оболочкой ВИЧ-1». Анналы Нью-Йоркской академии наук . 1010 (1): 19–28. Бибкод : 2003NYASA1010...19C . дои : 10.1196/анналы.1299.004 . ПМИД 15033690 . S2CID 37073602 .
- Нг С., Смит М.Б., Смит Х.Т., Миллетт Ф. (ноябрь 1977 г.). «Влияние модификации отдельных лизинов цитохрома с на реакцию с цитохромом b5». Биохимия . 16 (23): 4975–8. дои : 10.1021/bi00642a006 . ПМИД 199233 .
- Линч С.Р., Шерман Д., Коупленд Р.А. (январь 1992 г.). «Связывание цитохрома с влияет на конформацию цитохрома а в цитохром с оксидазе» . Журнал биологической химии . 267 (1): 298–302. дои : 10.1016/S0021-9258(18)48493-1 . ПМИД 1309738 .
- Гарбер Э.А., Марголиаш Э (февраль 1990 г.). «Взаимодействие цитохрома с с цитохром оксидазой: понимание перехода от высокого к низкому сродству». Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1015 (2): 279–87. дои : 10.1016/0005-2728(90)90032-Y . ПМИД 2153405 .
- Компакт-диск Бедетти (май 1985 г.). «Иммуноцитохимическая демонстрация цитохром-с-оксидазы иммунопероксидазным методом: специфическое окрашивание митохондрий в фиксированных формалином и залитых в парафин тканях человека» . Журнал гистохимии и цитохимии . 33 (5): 446–52. дои : 10.1177/33.5.2580882 . ПМИД 2580882 .
- Танака Ю., Асикари Т., Сибано Ю., Амачи Т., Ёшизуми Х., Мацубара Х. (июнь 1988 г.). «Конструирование гена цитохрома с человека и его функциональная экспрессия в Saccharomyces cerevisiae». Журнал биохимии . 103 (6): 954–61. doi : 10.1093/oxfordjournals.jbchem.a122393 . ПМИД 2844747 .
- Эванс MJ, Scarpulla RC (декабрь 1988 г.). «Ген соматического цитохрома с человека: два класса процессированных псевдогенов разграничивают период быстрой молекулярной эволюции» . Труды Национальной академии наук Соединенных Штатов Америки . 85 (24): 9625–9. Бибкод : 1988PNAS...85.9625E . дои : 10.1073/pnas.85.24.9625 . ПМК 282819 . ПМИД 2849112 .
- Пассон П.Г., Хультквист Д.Э. (июль 1972 г.). «Растворимая цитохром b 5 редуктаза из эритроцитов человека». Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 275 (1): 62–73. дои : 10.1016/0005-2728(72)90024-2 . hdl : 2027.42/34070 . ПМИД 4403130 .
- Доу Р.Дж., Вителло Л.Б., Эрман Дж.Э. (август 1984 г.). «Исследования седиментационного равновесия по взаимодействию цитохрома с и пероксидазы цитохрома с». Архив биохимии и биофизики . 232 (2): 566–73. дои : 10.1016/0003-9861(84)90574-5 . ПМИД 6087732 .
- Мишель Б., Bosshard HR (август 1984 г.). «Спектроскопический анализ взаимодействия цитохрома с и цитохром с оксидазой» . Журнал биологической химии . 259 (16): 10085–91. дои : 10.1016/S0021-9258(18)90932-4 . ПМИД 6088481 .
- Брогер С., Налеч М.Ю., Аззи А. (октябрь 1980 г.). «Взаимодействие цитохрома с с комплексом цитохрома bc1 дыхательной цепи митохондрий». Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 592 (3): 519–27. дои : 10.1016/0005-2728(80)90096-1 . ПМИД 6251869 .
- Смит Х.Т., Ахмед А.Дж., Миллетт Ф. (май 1981 г.). «Электростатическое взаимодействие цитохрома с с цитохромом с1 и цитохромоксидазой» . Журнал биологической химии . 256 (10): 4984–90. дои : 10.1016/S0021-9258(19)69355-5 . ПМИД 6262312 .
- Герен Л.М., Миллетт Ф. (октябрь 1981 г.). «Исследование переноса энергии флуоресценции взаимодействия адренодоксин и цитохром с» . Журнал биологической химии . 256 (20): 10485–9. дои : 10.1016/S0021-9258(19)68647-3 . ПМИД 6270113 .
- Фавр Б., Зольнерович С., Туровски П., Хеммингс Б.А. (июнь 1994 г.). «Каталитическая субъединица протеинфосфатазы 2А карбоксилметилирована in vivo» . Журнал биологической химии . 269 (23): 16311–7. дои : 10.1016/S0021-9258(17)34009-7 . ПМИД 8206937 .
- Гао Б., Айзенберг Э., Грин Л. (июль 1996 г.). «Влияние конститутивной полимеризации белка теплового шока массой 70 кДа на его взаимодействие с белковым субстратом» . Журнал биологической химии . 271 (28): 16792–7. дои : 10.1074/jbc.271.28.16792 . ПМИД 8663341 .
Внешние ссылки
[ редактировать ]- Белок Цитохром С
- Апоптоз и каспаза 3 – PMAP Карта протеолиза – анимация
- Цитохром + c в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Обзор всей структурной информации, доступной в PDB для UniProt : P99999 (Цитохром c) в PDBe-KB .
галерея PDB |
|---|

