Щавелоуксусная кислота
 | |
 | |
| Имена | |
|---|---|
| Предпочтительное название ИЮПАК 2-оксобутандиовая кислота | |
| Другие имена Щавелоуксусная кислота щавелуксусная кислота 2-оксоянтарная кислота Кетосукциновая кислота | |
| Идентификаторы | |
3D model ( JSmol ) | |
| ЧЭБИ | |
| ХимическийПаук | |
| Информационная карта ECHA | 100.005.755 |
| Номер ЕС |
|
| КЕГГ | |
ПабХим CID | |
| НЕКОТОРЫЙ | |
Панель управления CompTox ( EPA ) | |
| Характеристики | |
| С 4 Н 4 О 5 | |
| Молярная масса | 132.07 g/mol |
| Плотность | 1,6 г/см 3 |
| Температура плавления | 161 ° С (322 ° F; 434 К) |
| Термохимия | |
Стандартная энтальпия образование (Δ f H ⦵ 298 ) | -943,21 кДж/моль |
Стандартная энтальпия горение (Δ c H ⦵ 298 ) | -1205,58 кДж/моль |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
Щавелоуксусная кислота (также известная как щавелевоуксусная кислота или ОАА ) представляет собой кристаллическое органическое соединение с химической формулой HO 2 CC(O)CH 2 CO 2 H. Щавелоуксусная кислота в форме сопряженного с ней основания оксалоацетата является промежуточным продуктом метаболизма во многих процессы, происходящие у животных. Он принимает участие в глюконеогенезе , цикле мочевины , глиоксилатном цикле , синтезе аминокислот , синтезе жирных кислот и цикле лимонной кислоты . [1]
Характеристики
[ редактировать ]Щавелосуксусная кислота подвергается последовательному депротонированию с образованием дианиона :
- НО 2 CC(O)CH 2 CO 2 H ⇌ − О 2 CC(O)CH 2 CO 2 H + H + , пК а = 2,22
- − О 2 CC(O)CH 2 CO 2 H ⇌ − О 2 CC(O)CH 2 CO 2 − + Ч + , пК а = 3,89
При высоком pH енолизируемый протон ионизируется:
- − О 2 CC(O)CH 2 CO 2 − ⇌ − О 2 СС(О − )ХОЧУ 2 − + Ч + , пК а = 13,03
Енольные формы щавелевоуксусной кислоты особенно стабильны. Кето-енольная таутомеризация катализируется ферментом оксалоацетаттаутомеразой . Транс -енол-оксалоацетат также появляется, когда тартрат является субстратом для фумаразы . [2]

Биосинтез
[ редактировать ]Оксалоацетат в природе образуется несколькими способами. Основной путь – окисление L - малата , катализируемое малатдегидрогеназой , в цикле лимонной кислоты. Малат также окисляется сукцинатдегидрогеназой в медленной реакции, исходным продуктом которой является енол-оксалоацетат. [3]
Он также возникает в результате конденсации пирувата с угольной кислотой, вызванной гидролизом АТФ :
- СН 3 С(О)СО 2 − + HCO3 − + АТФ → − О 2 ССН 2 С(О)СО 2 − + АДФ + Пи
Происходя в мезофилле растений, этот процесс протекает через фосфоенолпируват , катализируемый фосфоенолпируваткарбоксилазой .
Оксалоацетат также может образовываться в результате транс- или дезаминирования аспарагиновой кислоты .
Биохимические функции
[ редактировать ]Оксалоацетат является промежуточным продуктом цикла лимонной кислоты , где он реагирует с ацетил-КоА с образованием цитрата , катализируемого цитратсинтазой . Он также участвует в глюконеогенезе , цикле мочевины , глиоксилатном цикле , синтезе аминокислот и синтезе жирных кислот . Оксалоацетат также является мощным ингибитором комплекса II .
глюконеогенез
[ редактировать ]глюконеогенез [1] представляет собой метаболический путь, состоящий из серии из одиннадцати ферментативно-катализируемых реакций, приводящих к образованию глюкозы из неуглеводных субстратов. Начало этого процесса происходит в митохондриальном матриксе , где пирувата находятся молекулы . Молекула пирувата карбоксилируется ферментом пируваткарбоксилазой , активируемым молекулой АТФ и воды. Эта реакция приводит к образованию оксалоацетата. НАДН восстанавливает оксалоацетат до малата . Эта трансформация необходима для транспортировки молекулы из митохондрий . Попав в цитозоль , малат снова окисляется до оксалоацетата с помощью НАД+. Тогда оксалоацетат остается в цитозоле, где и пройдут остальные реакции. Оксалоацетат позже декарбоксилируется и фосфорилируется фосфоенолпируваткарбоксикиназой ( GTP и превращается в 2-фосфоенолпируват с использованием гуанозинтрифосфата ) в качестве источника фосфата. Глюкозу получают после дальнейшей переработки.
Цикл мочевины
[ редактировать ]Цикл мочевины представляет собой метаболический путь, который приводит к образованию мочевины с использованием одной молекулы аммония из расщепленных аминокислот, другой группы аммония из аспартата и одной молекулы бикарбоната. [1] Этот путь обычно происходит в гепатоцитах . Реакции, связанные с циклом мочевины, производят НАДН , а НАДН можно получить двумя разными способами. Один из них использует оксалоацетат . В цитозоле имеются молекулы фумарата . Фумарат может превращаться в малат под действием фермента фумаразы . Малат под действием малатдегидрогеназы превращается в оксалоацетат, образуя молекулу НАДН. После этого оксалоацетат будет переработан в аспартат , поскольку трансаминазы предпочитают именно эти кетокислоты остальным. Эта рециркуляция поддерживает приток азота в клетку.

Глиоксилатный цикл
[ редактировать ]Глиоксилатный цикл является разновидностью цикла лимонной кислоты. [4] Это анаболический путь, происходящий в растениях и бактериях с использованием ферментов изоцитратлиазы и малатсинтазы . Некоторые промежуточные этапы цикла немного отличаются от цикла лимонной кислоты; тем не менее, оксалоацетат выполняет одну и ту же функцию в обоих процессах. [1] Это означает, что оксалоацетат в этом цикле также выступает в качестве основного реагента и конечного продукта. Фактически оксалоацетат является чистым продуктом глиоксилатного цикла , поскольку его петля цикла включает две молекулы ацетил-КоА.
Синтез жирных кислот
[ редактировать ]На предыдущих стадиях ацетил-КоА переносится из митохондрий в цитоплазму, где находится синтаза жирных кислот . Ацетил-КоА транспортируется в виде цитрата, который ранее образовался в митохондриальном матриксе из ацетил-КоА и оксалоацетата. Эта реакция обычно инициирует цикл лимонной кислоты, но когда нет необходимости в энергии, она транспортируется в цитоплазму, где расщепляется до цитоплазматического ацетил-КоА и оксалоацетата.
Другая часть цикла требует НАДФН для синтеза жирных кислот. [5] Часть этой восстанавливающей способности генерируется, когда цитозольный оксалоацетат возвращается в митохондрии, пока внутренний слой митохондрий непроницаем для оксалоацетата. Сначала оксалоацетат восстанавливают до малата с помощью НАДН. Затем малат декарбоксилируется до пирувата. Теперь этот пируват может легко проникать в митохондрии, где он снова карбоксилируется до оксалоацетата пируваткарбоксилазой. Таким образом, перенос ацетил-КоА из митохондрий в цитоплазму приводит к образованию молекулы НАДН. Общую реакцию, которая является спонтанной, можно резюмировать следующим образом:
- HCOHCO3 – + АТФ + ацетил-КоА → АДФ + Ф i + малонил-КоА
Синтез аминокислот
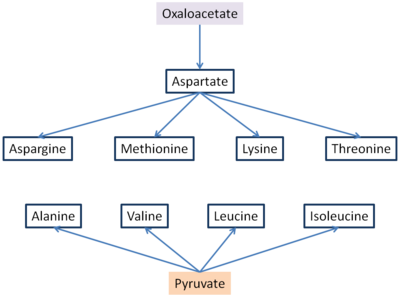
[ редактировать ]Шесть незаменимых аминокислот и три заменимые синтезируются из оксалоацетата и пирувата . [6] Аспартат и аланин образуются из оксалоацетата и пирувата соответственно путем переаминирования из глутамата . Аспарагин синтезируется путем амидирования аспартата, при этом глутамин отдает NH4.Это заменимые аминокислоты, и их простые пути биосинтеза встречаются во всех организмах. Метионин, треонин, лизин, изолейцин, валин и лейцин являются незаменимыми аминокислотами для человека и большинства позвоночных. Пути их биосинтеза у бактерий сложны и взаимосвязаны.
Биосинтез оксалата
[ редактировать ]Оксалоацетат образует оксалат путем гидролиза. [7]
- оксалоацетат + H 2 O ⇌ оксалат + ацетат
Этот процесс катализируется ферментом оксалоацетазой . Этот фермент встречается у растений, но неизвестен в животном мире. [8]
Интерактивная карта маршрутов
[ редактировать ]| Нажмите на гены, белки и метаболиты ниже, чтобы перейти к соответствующим статьям. [§ 1] Гликолиз и глюконеогенез
| Нажмите на гены, белки и метаболиты ниже, чтобы перейти к соответствующим статьям. [§ 1] TCACycle_WP78 редактировать
|


См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д Нельсон, Дэвид Л.; Кокс, Майкл М. (2005). Принципы биохимии (4-е изд.). Нью-Йорк: WH Freeman. ISBN 0-7167-4339-6 .
- ^ ван Вугт-Люссенбург, BMA; ван дер Вель, Л; Хаген, WR; Хагедорн, Польша (26 февраля 2021 г.), «Биохимические сходства и различия между каталитическим кластером [4Fe-4S], содержащим фумаразы FumA и FumB из Escherichia coli », PLOS ONE , 8 (2) (опубликовано 6 февраля 2013 г.): e55549, doi : 10.1371/journal.pone.0055549 , PMC 3565967 , PMID 23405168
- ^ М.В. Панченко; А.Д. Виноградов (1991). «Прямая демонстрация енол-оксалоацетата как непосредственного продукта окисления малата сукцинатдегидрогеназой млекопитающих» . Письма ФЭБС . 286 (1–2): 76–78. Бибкод : 1991FEBSL.286...76P . дои : 10.1016/0014-5793(91)80944-X . ПМИД 1864383 .
- ^ «Добро пожаловать в Химический зал» . www.pearsonhighered.com . Проверено 5 апреля 2018 г.
- ^ «синтез жирных кислот» . www.rpi.edu .
- ^ «Анимокислоты, синтезированные из оксалоацетата и пирувата» . факультет.ksu.edu.sa . Архивировано из оригинала (PPTX) 21 октября 2013 года . Проверено 21 октября 2013 г.
- ^ Гэдд, Джеффри М. «Грибковое производство лимонной и щавелевой кислоты: значение в видообразовании металлов, физиологии и биогеохимических процессах» Достижения в микробной физиологии (1999), 41, 47-92.
- ^ Сюй, Хуа-Вэй. «Накопление и регуляция оксалатов не зависят от гликолатоксидазы в листьях риса» Журнал экспериментальной ботаники, том 57, № 9, стр. 1899–1908, 2006 г.









