Тургорное давление
Тургорное давление – это сила внутри клетки, которая прижимает плазматическую мембрану к клеточной стенке . [1]
Его также называют гидростатическим давлением и определяют как давление в жидкости, измеренное в определенной точке внутри нее, когда она находится в равновесии. [2] Обычно тургорное давление обусловлено осмотическим потоком воды и встречается у растений , грибов и бактерий . Это явление также наблюдается у протистов , имеющих клеточные стенки. [3] Эта система не наблюдается в клетках животных, поскольку отсутствие клеточной стенки может привести к лизису клетки под слишком большим давлением. [4] Давление, оказываемое осмотическим потоком воды, называется тургидностью. Это вызвано осмотическим потоком воды через избирательно проницаемую мембрану . Движение воды через полупроницаемую мембрану из объема с низкой концентрацией растворенного вещества в объем с более высокой концентрацией растворенного вещества называется осмотическим потоком. клетки У растений это означает, что вода перемещается из растворенного вещества с низкой концентрацией вне клетки в вакуоль . [ нужна ссылка ]
Этимология
[ редактировать ]1610-е годы, от латинского turgidus «опухший, раздутый, раздутый», от turgere «набухать», неизвестного происхождения. Образное использование в прозе относится к 1725 году. Связано: Напыщенно; напыщенность.
Механизм
[ редактировать ]
Осмос – это процесс, при котором вода вытекает из объема с низкой концентрацией растворенного вещества (осмолярностью), [5] в соседнюю область с более высокой концентрацией растворенного вещества, пока не будет достигнуто равновесие между двумя областями. [6] Обычно это сопровождается благоприятным увеличением энтропии растворителя . Все клетки окружены двухслойной липидной клеточной мембраной, которая обеспечивает поступление воды в клетку и из нее, ограничивая при этом поток растворенных веществ. Когда клетка находится в гипертоническом растворе, вода вытекает из клетки, что уменьшает ее объем. В гипотоническом растворе вода поступает в мембрану и увеличивает объем клетки, тогда как в изотоническом растворе вода поступает в клетку и выходит из нее с одинаковой скоростью. [4]
Тургидность — это точка, в которой клеточная мембрана прижимается к клеточной стенке, когда тургорное давление высокое. Когда клетка имеет низкое тургорное давление, она вялая. У растений это проявляется в виде увядания анатомических структур. Это более конкретно известно как плазмолиз. [7]

Объем и геометрия клетки влияют на величину тургорного давления и на то, как оно может влиять на пластичность клеточной стенки. Исследования показали, что более мелкие клетки испытывают более сильные упругие изменения по сравнению с более крупными клетками. [3]
Тургорное давление также играет ключевую роль в росте растительных клеток, когда клеточная стенка подвергается необратимому расширению из-за силы тургорного давления, а также структурных изменений в клеточной стенке, которые изменяют ее растяжимость. [8]
Тургорное давление у растений
[ редактировать ]Тургорное давление внутри клеток регулируется осмосом, что также приводит к расширению клеточной стенки во время роста. Помимо размера, жесткость клетки обусловлена тургорным давлением; более низкое давление приводит к увяданию клеток или структуры растения (например, листа, стебля). Одним из механизмов регулирования тургорного давления у растений является полупроницаемая мембрана клетки, которая позволяет только некоторым растворенным веществам проникать в клетку и выходить из нее, поддерживая минимальное давление. Другие механизмы включают транспирацию , которая приводит к потере воды и уменьшению тургидности клеток. [9] Тургорное давление также является важным фактором транспорта питательных веществ по растению. Клетки одного и того же организма могут иметь разное тургорное давление по всей структуре организма. У сосудистых растений тургорное давление отвечает за апикальный рост таких элементов, как кончики корней. [10] и пыльцевые трубки . [11]
Рассредоточение
[ редактировать ]Транспортные белки , которые перекачивают растворенные вещества в клетку, могут регулироваться тургорным давлением клетки. Более низкие значения позволяют увеличить перекачку растворенных веществ, что, в свою очередь, увеличивает осмотическое давление. Эта функция важна как реакция растений в условиях засухи. [12] (так как сохраняется тургорное давление), и для клеток, которым необходимо накапливать растворенные вещества (т.е. развивающихся плодов ). [13]
Органы цветения и размножения
[ редактировать ]Было зарегистрировано, что лепестки Gentiana kochiana и Kalanchoe blossfeldiana поверхности растения цветут за счет изменчивого тургорного давления клеток на адаксиальной . [11] во время таких процессов, как пыльника раскрытие Было замечено, что , высыхающие клетки эндотеция вызывают силу изгиба наружу, что приводит к высвобождению пыльцы. Это означает, что в этих структурах наблюдается пониженное тургорное давление из-за того, что они обезвожены. Пыльцевые трубки — это клетки, которые удлиняются, когда пыльца попадает на рыльце пестика на кончике запястья. Эти клетки довольно быстро растут кончиками из-за увеличения тургорного давления. Пыльцевая трубка лилий при росте в ходе этого процесса имеет среднее тургорное давление 0,21 МПа. [14]
Распространение семян
[ редактировать ]
В таких плодах, как Impatiens parviflora , Oxalia acetosella и Ecballium elaterium , тургорное давление является методом диспергирования семян. [15] У Ecballium elaterium , или брызгающего огурца, тургорное давление в плодах возрастает до такой степени, что они агрессивно отрываются от плодоножки, а семена и вода разбрызгиваются повсюду, когда плод падает на землю. Тургорное давление внутри плода колеблется от 0,003 до 1,0 МПа. [16]
Рост
[ редактировать ]Обычно считается, что движущей силой роста внутри клетки является действие тургорного давления на растяжимые клеточные стенки. [17] Увеличение тургорного давления вызывает расширение клеток и удлинение апикальных клеток, пыльцевых трубок и других структур растения, таких как кончики корней. Расширение клетки и повышение тургорного давления происходит за счет диффузии воды внутрь клетки, а тургорное давление увеличивается за счет увеличения объема вакуолярного сока . Тургорное давление растущей клетки корня может достигать 0,6 МПа, что более чем в три раза превышает давление автомобильной шины. Эпидермальные клетки листа могут иметь давление от 1,5 до 2,0 МПа. [18] Такое высокое давление может объяснить, почему растения могут расти сквозь асфальт и другие твердые поверхности. [17]
Напухлость
[ редактировать ]Набухание наблюдается в клетке, где клеточная мембрана прижимается к клеточной стенке. У некоторых растений клеточные стенки разрыхляются быстрее, чем вода может проникнуть через мембрану, что приводит к образованию клеток с более низким тургорным давлением. [3]
устьица
[ редактировать ]
Тургорное давление внутри устьиц регулирует, когда устьица могут открываться и закрываться, что играет роль в скорости транспирации растения. Это также важно, поскольку эта функция регулирует потерю воды внутри растения. Более низкое тургорное давление может означать, что в клетке низкая концентрация воды, и закрытие устьиц поможет сохранить воду. Высокое тургорное давление сохраняет устьица открытыми для газообмена, необходимого для фотосинтеза. [9]
Мимоза стыдливая
[ редактировать ]
Сделан вывод, что потеря тургорного давления в листьях Mimosa pudica ответственна за реакцию растения при прикосновении. другие факторы, такие как изменения осмотического давления, сокращение протоплазмы и увеличение клеточной проницаемости Было обнаружено, что на этот ответ влияют и . Установлено также, что тургорное давление в верхних и нижних легочных клетках растения различно, а движение ионов калия и кальция по клеткам вызывает повышение тургорного давления. При прикосновении пульвинус активируется и выделяет сократительные белки, что, в свою очередь, повышает тургорное давление и смыкает листья растения. [19]
Функция в других таксонах
[ редактировать ]Как уже говорилось ранее, тургорное давление можно обнаружить не только у растений, но и у других организмов, и оно может играть большую роль в развитии, движении и природе указанных организмов.
Грибы
[ редактировать ]
У грибов тургорное давление является важным фактором проникновения субстрата . У таких видов, как Saprolegniaferax, Magnaporthe grisea и Aspergillus oryzae , наблюдалось огромное тургорное давление в их гифах . Исследование показало, что они могут проникать в такие вещества, как растительные клетки и синтетические материалы, такие как поливинилхлорид . [20] При наблюдении этого явления отмечается, что инвазивный рост гиф обусловлен тургорным давлением, а также коферментами, секретируемыми грибами для проникновения в указанные субстраты. [21] Рост гиф напрямую связан с тургорным давлением, и рост замедляется по мере снижения тургорного давления. В Magnaporthe grisea наблюдалось давление до 8 МПа. [22]
Протисты
[ редактировать ]Некоторые протисты не имеют клеточных стенок и не могут испытывать тургорное давление. Эти немногие протисты используют свою сократительную вакуоль для регулирования количества воды внутри клетки. Клетки протистов избегают лизиса в гипотоническом растворе, используя вакуоль, которая выкачивает воду из клеток для поддержания осмотического равновесия. [23]
Животные
[ редактировать ]тургорное давление не наблюдается, В животных клетках поскольку у них отсутствует клеточная стенка. У организмов с клеточными стенками клеточная стенка предотвращает лизис клетки под действием высокого тургорного давления. [1]
Диатомовые водоросли
[ редактировать ]У диатомовых водорослей Heterokontophyta имеют полифилетическую тургор-резистентную клеточную стенку. На протяжении всего жизненного цикла этих организмов тщательно контролируемое тургорное давление отвечает за расширение клеток и высвобождение сперматозоидов, но не за такие процессы, как рост щетинок . [24]
Цианобактерии
[ редактировать ]газовакуумный [ проверьте орфографию ] Цианобактерии обычно ответственны за цветение воды . Они обладают способностью плавать благодаря накоплению газов внутри вакуолей, а о роли тургорного давления и его влиянии на емкость этих вакуолей сообщалось в различных научных статьях. [25] [26] Отмечено, что чем выше тургорное давление, тем ниже емкость газовакуолей у разных цианобактерий. Эксперименты, проведенные для корреляции осмоса и тургорного давления у прокариот, были использованы, чтобы показать, как диффузия растворенных веществ в клетку влияет на тургорное давление внутри клетки. [27]
Измерения
[ редактировать ]При измерении тургорного давления у растений приходится учитывать множество факторов. Обычно утверждается, что полностью набухшие клетки имеют тургорное давление, равное тургорному давлению самой клетки, а вялые клетки имеют значение тургора, равное или близкое к нулю. Другие клеточные механизмы, которые следует принимать во внимание, включают протопласт , растворенные вещества внутри протопласта (потенциал растворенного вещества), скорость транспирации клетки и натяжение клеточных стенок. Измерение ограничено в зависимости от используемого метода, некоторые из которых рассмотрены и объяснены ниже. Не все методы можно использовать для всех организмов из-за размера или других свойств. Например, диатомея не обладает теми же свойствами, что и растение, что налагает ограничения на методы, которые можно использовать для определения тургорного давления. [28]
Единицы
[ редактировать ]Единицы измерения тургорного давления не зависят от мер, используемых для определения его значений. Общие единицы включают бары , МПа или ньютоны на квадратный метр. 1 бар равен 0,1 МПа. [29]
Методы
[ редактировать ]Уравнение водного потенциала
[ редактировать ]Тургорное давление можно определить, если общий водный потенциал Ψ w и осмотический потенциал Ψ s . в уравнении водного потенциала известны [30] Эти уравнения используются для измерения общего водного потенциала растения с использованием таких переменных, как матричный потенциал, осмотический потенциал, потенциал давления, гравитационные эффекты и тургорное давление. [31] После взятия разницы между Ψ s и Ψ w получают значение тургорного давления. При использовании этого метода гравитация и матричный потенциал считаются пренебрежимо малыми, поскольку их значения, как правило, либо отрицательны, либо близки к нулю. [30]
Техника «бомбы давления»
[ редактировать ]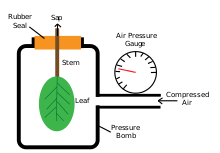
Техника напорной бомбы была разработана Шоландером и др., рассмотренная Тайри и Хаммелом в их публикации 1972 года, для проверки движения воды через растения. Прибор используется для измерения тургорного давления путем помещения листа (с прикрепленным стеблем) в закрытую камеру, куда постепенно добавляется газ под давлением. Измерения проводят в момент выхода сока ксилемы из поверхности разреза и в месте, где он не скапливается и не отступает обратно на поверхность разреза. [32]
Атомно-силовой микроскоп
[ редактировать ]В атомно-силовых микроскопах используется разновидность сканирующей зондовой микроскопии (СЗМ). Небольшие зонды вводятся в интересующую область, а пружина внутри зонда измеряет значения путем перемещения. [33] Этот метод можно использовать для измерения тургорного давления организмов. При использовании этого метода дополнительная информация, такая как уравнения механики сплошной среды , кривые глубины одиночной силы и геометрия клеток, может использоваться для количественной оценки тургорного давления в пределах заданной области (обычно клетки).
Датчик давления
[ редактировать ]Первоначально эта машина использовалась для измерения отдельных клеток водорослей , но теперь ее можно использовать и для более крупных образцов клеток. Обычно его используют на тканях высших растений , но не использовали для измерения тургорного давления до тех пор, пока Хюскен и Циммерман не усовершенствовали этот метод. [34] Датчики давления измеряют тургорное давление посредством смещения. В клетку вставляют стеклянную микрокапиллярную трубку, и все, что клетка выделяет в трубку, наблюдают под микроскопом. Затем подключенное устройство измеряет, какое давление необходимо, чтобы вытолкнуть выброс обратно в ячейку. [32]
Микроманипуляционный зонд
[ редактировать ]Они используются для точной количественной оценки размеров более мелких клеток. В эксперименте Вебера, Смита и его коллег отдельные клетки томата были сжаты между микроманипуляционным зондом и стеклом, чтобы позволить микрокапилляру зонда давления определить тургорное давление клетки. [35]
Теоретические размышления
[ редактировать ]Отрицательное тургорное давление
[ редактировать ]Было замечено, что значение Ψ w уменьшается по мере того, как клетка становится более обезвоженной, [30] но ученые предполагают, будет ли это значение продолжать уменьшаться, но никогда не упадет до нуля, или же оно может быть меньше нуля. Были исследования [36] [37] которые показывают, что отрицательное клеточное давление может существовать в ксерофитных растениях, но статья М.Т. Тайри исследует, возможно ли это, или же вывод основан на неверно истолкованных данных. Он заключает, что утверждения об отрицательных значениях тургорного давления были неверными и возникли в результате неправильной классификации «связанной» и «свободной» воды в клетке. Анализируя изотермы апопластной и симпластической воды, он показывает, что отрицательное тургорное давление не может присутствовать у засушливых растений из-за чистой потери воды экземпляром во время засухи. Несмотря на этот анализ и интерпретацию данных, в научном сообществе до сих пор используются отрицательные значения тургорного давления. [38]
Рост кончиков у высших растений
[ редактировать ]Гипотеза, представленная М. Гарольдом и его коллегами, предполагает, что рост кончиков у высших растений имеет амебную природу и не вызван тургорным давлением, как широко распространено мнение, а это означает, что растяжение вызывается актиновым цитоскелетом в этих растительных клетках. Предполагается, что регуляция роста клеток осуществляется цитоплазматическими микротрубочками, которые контролируют ориентацию фибрилл целлюлозы, которые откладываются в соседней клеточной стенке и приводят к росту. У растений клетки окружены клеточными стенками и нитчатыми белками, которые сохраняют и регулируют рост и форму растительной клетки. Сделан вывод, что низшие растения растут за счет апикального роста, который отличается тем, что клеточная стенка расширяется только на одном конце клетки. [39]
Ссылки
[ редактировать ]- ^ Jump up to: а б Причард, Джереми (2001). «Тургорное давление». Энциклопедия наук о жизни . Американское онкологическое общество. дои : 10.1038/npg.els.0001687 . ISBN 9780470015902 .
- ^ Фрике, Виланд (январь 2017 г.). «Тургорное давление». Энциклопедия наук о жизни . стр. 1–6. дои : 10.1002/9780470015902.a0001687.pub2 . ISBN 9780470015902 .
- ^ Jump up to: а б с Штойдле, Эрнст (февраль 1977 г.). «Влияние тургорного давления и размера клеток на эластичность стенок растительных клеток» . Физиология растений . 59 (2): 285–9. дои : 10.1104/стр.59.2.285 . ПМК 542383 . ПМИД 16659835 .
- ^ Jump up to: а б «Осмос и тоничность» . Ханская академия . Проверено 27 апреля 2017 г.
- ^ Кеппен, Брюс М.; Стэнтон, Брюс А. (2013). Физиология почек (Пятое изд.). Филадельфия, Пенсильвания. ISBN 978-0-323-08825-1 . OCLC 815507871 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ «GCSE Bitesize: осмос в клетках» . Би-би-си .
- ^ «Плазмолиз в клетках растений элодеи - Science NetLinks» . sciencenetlinks.com . Проверено 27 апреля 2017 г.
- ^ Джордан, БМ; Дюмэ, Дж. (2010). «Биомеханика роста растительных клеток» . Энциклопедия наук о жизни .
- ^ Jump up to: а б Ваггонер, Пол Э.; Зелич, Израиль (10 декабря 1965 г.). «Транспирация и устьица листьев». Наука . 150 (3702): 1413–1420. Бибкод : 1965Sci...150.1413W . дои : 10.1126/science.150.3702.1413 . ПМИД 17782290 .
- ^ Симадзаки, Юми; Оокава, Тайитиро; Хирасава, Тадаши (1 сентября 2005 г.). «Кончик корня и ускоряющая область подавляют удлинение замедляющейся области без какого-либо влияния на тургор клеток в первичных корнях кукурузы в условиях водного стресса» . Физиология растений . 139 (1): 458–465. дои : 10.1104/стр.105.062091 . ПМЦ 1203394 . ПМИД 16100358 .
- ^ Jump up to: а б Бозами, Лена; Накаяма, Наоми; Будауд, Арезки (1 ноября 2014 г.). «Цветы под давлением: тонкости регуляции тургора в развитии» . Анналы ботаники . 114 (7): 1517–1533. дои : 10.1093/aob/mcu187 . ПМК 4204789 . ПМИД 25288632 .
- ^ Фишер, Дональд Б.; Кэш-Кларк, Кора Э. (27 апреля 2017 г.). «Градиенты водного потенциала и тургорного давления на пути транслокации во время налива зерна у нормально поливаемых и испытывающих водный стресс растений пшеницы» . Физиология растений . 123 (1): 139–148. дои : 10.1104/стр.123.1.139 . ПМК 58989 . ПМИД 10806232 .
- ^ Келлер, Маркус; Шреста, Прадип М. (2014). «Накопление растворенных веществ различается в вакуолях и апопласте созревающих ягод винограда». Планта . 239 (3): 633–642. дои : 10.1007/s00425-013-2004-z . ПМИД 24310282 . S2CID 15443543 .
- ^ Бенкерт, Райнер; Обермайер, Герхард; Бентруп, Фридрих-Вильгельм (1 марта 1997 г.). «Тургорное давление растущих пыльцевых трубок лилии». Протоплазма . 198 (1–2): 1–8. дои : 10.1007/BF01282125 . S2CID 23911884 .
- ^ Хаяши, М.; Фейлих, КЛ; Эллерби, диджей (1 мая 2009 г.). «Механика взрывного распространения семян оранжевой драгоценности ( Impatiens capensis )» . Журнал экспериментальной ботаники . 60 (7): 2045–2053. дои : 10.1093/jxb/erp070 . ПМЦ 2682495 . ПМИД 19321647 .
- ^ Козловский, Т.Т. (2012). Биология семян: значение, развитие и прорастание . Том. 1. Академическая пресса. стр. 195–196.
- ^ Jump up to: а б Крегер, Йенс Х.; Зерзур, Рабах; Гейтманн, Аня (25 апреля 2011 г.). «Регулятор или движущая сила? Роль тургорного давления в колебательном росте растительных клеток» . ПЛОС ОДИН . 6 (4): e18549. Бибкод : 2011PLoSO...618549K . дои : 10.1371/journal.pone.0018549 . ПМК 3081820 . ПМИД 21541026 .
- ^ Серпе, Марсело Д.; Мэтьюз, Марк А. (1 января 1994 г.). «Рост, давление и напряжение стенок эпидермальных клеток листьев бегонии argenteo-guttata L. во время развития». Международный журнал наук о растениях . 155 (3): 291–301. дои : 10.1086/297168 . JSTOR 2475182 . S2CID 84209016 .
- ^ Аллен, Роберт Д. (1 августа 1969 г.). «Механизм сейсмонастической реакции у Mimosa pudica1» . Физиология растений . 44 (8): 1101–1107. дои : 10.1104/стр.44.8.1101 . ПМК 396223 . ПМИД 16657174 .
- ^ Ховард, Ричард (декабрь 1991 г.). «Проникновение грибка в твердые субстраты с применением огромного тургорного давления» . Учеб. Натл. акад. Наука . 88 (24): 11281–11284. Бибкод : 1991PNAS...8811281H . дои : 10.1073/pnas.88.24.11281 . ПМЦ 53118 . ПМИД 1837147 .
- ^ Жерве, Патрик; Абади, Кристоф; Молин, Пол (1999). «Тургорное давление грибковых клеток: теоретический подход и измерение» . Журнал научных и промышленных исследований . 58 (9): 671–677.
- ^ Деньги, Николас П. (31 декабря 1995 г.). «Тургорное давление и механика проникновения грибов». Канадский журнал ботаники . 73 (С1): 96–102. дои : 10.1139/b95-231 .
- ^ «Пирсон - Место биологии» . www.phschool.com . Проверено 27 апреля 2017 г.
- ^ Рэйвен, Дж.А.; Уэйт, AM (1 апреля 2004 г.). «Эволюция окварцевания диатомовых водорослей: неизбежное погружение и погружение как спасение?» . Новый фитолог . 162 (1): 45–61. дои : 10.1111/j.1469-8137.2004.01022.x .
- ^ Кинсман, Р. (январь 1991 г.). «Коллапс газовых пузырьков под действием тургорного давления и его роль в регуляции плавучести Anabaena flos-aquae » . Журнал общей микробиологии . 143 (3): 1171–1178. дои : 10.1099/00221287-137-5-1171 .
- ^ Рид, Р.Х.; Уолсби, AE (1 декабря 1985 г.). «Изменение тургорного давления в ответ на увеличение внешней концентрации NaCl у газовакуолятивной цианобактерии Microcystis sp». Архив микробиологии . 143 (3): 290–296. дои : 10.1007/BF00411252 . S2CID 25006411 .
- ^ Оливер, Родерик Льюис (1 апреля 1994 г.). «Плавающие и тонущие газовакуолятивные цианобактерии1». Журнал психологии . 30 (2): 161–173. дои : 10.1111/j.0022-3646.1994.00161.x . S2CID 83747596 .
- ^ Томос, АД; Ли, РА; Шоу, Калифорния; Джонс, RGW (1 ноября 1984 г.). «Сравнение методов измерения тургорного давления и осмотического давления клеток запасающей ткани столовой свеклы». Журнал экспериментальной ботаники . 35 (11): 1675–1683. дои : 10.1093/jxb/35.11.1675 .
- ^ «Что такое единица давления «бар» (б)» . www.aqua-calc.com . Проверено 27 апреля 2017 г.
- ^ Jump up to: а б с Крамер, Пол (2012). Водные отношения растений . Эльзевир Наука . ISBN 978-0124250406 . ОСЛК 897023594 .
- ^ Безграничный (26 мая 2016 г.). «Давление, гравитация и матричный потенциал» . Безграничный .
- ^ Jump up to: а б Тайри, Монтана; Хаммель, ХТ (1972). «Измерение тургорного давления и водных отношений растений методом аэробомбы». Журнал экспериментальной ботаники . 23 (1): 267–282. дои : 10.1093/jxb/23.1.267 .
- ^ Бозами, Лена (май 2015 г.). «Количественное определение гидростатического давления в растительных клетках с помощью вдавливания с помощью атомно-силового микроскопа» . Биофизический журнал . 108 (10): 2448–2456. Бибкод : 2015BpJ...108.2448B . дои : 10.1016/j.bpj.2015.03.035 . ПМК 4457008 . ПМИД 25992723 .
- ^ Хюскен, Дитер; Штойдле, Эрнст; Циммерманн, Ульрих (1 февраля 1978 г.). «Техника измерения водного баланса клеток высших растений с помощью зонда давления» . Физиология растений . 61 (2): 158–163. дои : 10.1104/стр.61.2.158 . ПМЦ 1091824 . ПМИД 16660252 .
- ^ Вебер, Ален; Брейбрук, Шивон; Хуфлейт, Михал; Моска, Габриэлла; Рутье-Кержковска, Анн-Лиза; Смит, Ричард С. (1 июня 2015 г.). «Измерение механических свойств растительных клеток путем сочетания микроиндентирования с осмотической обработкой» . Журнал экспериментальной ботаники . 66 (11): 3229–3241. дои : 10.1093/jxb/erv135 . ПМЦ 4449541 . ПМИД 25873663 .
- ^ Ян, Дунмей; Ли, Цзюньхуэй; Дин, Итин; Тайри, Мелвин Т. (1 марта 2017 г.). «Экспериментальные доказательства отрицательного тургорного давления в мелких клетках листа Robinia pseudoacacia L по сравнению с крупными клетками Metasequoia glyptostroboides Hu et WC Cheng. 2. Диаграммы Хефлера ниже объема нулевого тургора и теоретическое значение для кривых давление-объем живых клеток» . Растение, клетка и окружающая среда . 40 (3): 340–350. дои : 10.1111/шт.12860 . ПМИД 27861986 .
- ^ Оэртли, Джей-Джей (июль 1986 г.). «Влияние размера клеток на коллапс клеток под отрицательным тургорным давлением». Журнал физиологии растений . 124 (3–4): 365–370. дои : 10.1016/S0176-1617(86)80048-7 .
- ^ Тайри, М. (январь 1976 г.). «Отрицательное тургорное давление в растительных клетках: факт или заблуждение?». Канадский журнал ботаники . 54 (23): 2738–2746. дои : 10.1139/b76-294 .
- ^ Пикетт-Хипс, доктор юридических наук; Кляйн, АГ (1998). «Рост кончиков растительных клеток может быть амебоидным и не вызываться тургорным давлением» . Труды: Биологические науки . 265 (1404): 1453–1459. дои : 10.1098/rspb.1998.0457 . ПМК 1689221 .
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| растений Группы | |||||||||||
| Анатомия растений |
| ||||||||||
| Физиология растений Материалы | |||||||||||
| Рост растений и привычка | |||||||||||
| Воспроизведение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||