Меристема
В клеточной биологии меристема встречающийся — это тип ткани, в растениях. Он состоит из недифференцированных клеток ( меристематических клеток ), способных к клеточному делению . Клетки меристемы могут развиваться во все другие ткани и органы растений. Эти клетки продолжают делиться, пока не станут дифференцированными и не потеряют способность делиться.
Дифференцированные растительные клетки обычно не могут делиться или производить клетки другого типа. Меристематические клетки недифференцированы или неполностью дифференцированы. Они тотипотентны и способны к продолжению клеточного деления . Деление меристематических клеток обеспечивает появление новых клеток для разрастания и дифференцировки тканей и зарождения новых органов, обеспечивающих основную структуру тела растения. Клетки мелкие, с небольшими вакуолями или без них, протоплазма полностью заполняет клетку. Пластиды хлоропласты ( пропластиды или хромопласты ) недифференцированы, но присутствуют в рудиментарной форме ( ) . Меристематические клетки плотно прилегают друг к другу, без межклеточных пространств. Клеточная стенка представляет собой очень тонкую первичную клеточную стенку.
Термин меристема впервые был использован в 1858 году швейцарским ботаником Карлом Вильгельмом фон Нэгели (1817–1891) в его книге «Вклад в научную ботанику» . [1] Оно происходит от греческого μερίζειν (меризеин) «разделять» в знак признания присущей ему функции. [ нужна ссылка ]
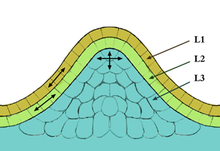
Существует три типа меристематических тканей: апикальные (на кончиках), интеркалярные или базальные (в середине) и латеральные (по бокам, также известные как камбий). На вершине меристемы находится небольшая группа медленно делящихся клеток, которую принято называть центральной зоной. Клетки этой зоны выполняют функцию стволовых клеток и необходимы для поддержания меристемы. Скорость пролиферации и роста на вершине меристемы обычно значительно отличается от таковой на периферии.
Первичные меристемы
[ редактировать ]Апикальные меристемы дают начало первичному телу растения и отвечают за первичный рост или увеличение длины или высоты. [2] [3] Апикальные меристемы могут различаться на три типа первичной меристемы:
- Протодерма : лежит вокруг стебля снаружи и развивается в эпидермис .
- Прокамбий : лежит внутри протодермы и развивается в первичную ксилему и первичную флоэму . Он также производит сосудистый камбий и пробковый камбий (вторичные меристемы). Пробковый камбий далее дифференцируется на феллодерму (внутри) и феллему, или пробку (наружу). Все три слоя (пробковый камбий, феллема и феллодерма) составляют перидерму . В корнях прокамбий также может давать начало перициклу, который дает боковые корни у эвдикотов. [4]
- Основная меристема : состоит из паренхимы , колленхимы и склеренхимы. клеток [4] которые развиваются в кору и сердцевину .
Вторичные меристемы
[ редактировать ]После первичного роста боковые меристемы развиваются как вторичный рост растения. Этот рост увеличивает диаметр растения по сравнению с укоренившимся стеблем, но не все растения демонстрируют вторичный рост. Различают два типа вторичных меристем: сосудистый камбий и пробковый камбий.
- Сосудистый камбий , образующий вторичную ксилему и вторичную флоэму. Этот процесс может продолжаться на протяжении всей жизни растения. Именно это дает начало древесине у растений. Такие растения называются древесными . Этого не происходит у растений, которые не проходят вторичный рост (так называемые травянистые растения).
- Пробковый камбий , дающий начало перидерме, заменяющей эпидермис.
Апикальные меристемы
Апикальные меристемы — это совершенно недифференцированные (индетерминированные) меристемы растения. Они различаются на три типа первичных меристем. Первичные меристемы, в свою очередь, производят два типа вторичных меристем. Эти вторичные меристемы также известны как латеральные меристемы, поскольку они участвуют в латеральном росте.

- Центральная зона
- Периферийная зона
- Медуллярная (т.е. центральная) меристема
- Медуллярная ткань
Существует два типа ткани апикальной меристемы: апикальная меристема побега ( SAM ), которая дает начало таким органам, как листья и цветы, и апикальная меристема корня ( RAM ), которая обеспечивает меристематические клетки для будущего роста корня. Клетки SAM и RAM быстро делятся и считаются неопределенными, поскольку они не обладают каким-либо определенным конечным статусом. В этом смысле меристематические клетки часто сравнивают со стволовыми клетками животных, которые имеют аналогичное поведение и функции.
Апикальные меристемы многослойные, количество слоев варьируется в зависимости от типа растения. Обычно самый внешний слой называется оболочкой , а самые внутренние слои — телом . У однодольных растений оболочка определяет физические характеристики края и края листа. У двудольных второй слой корпуса определяет характеристики края листа. Тело и оболочка играют важную роль в физическом облике растения, поскольку все растительные клетки образуются из меристем. Апикальные меристемы встречаются в двух местах: в корне и стебле. Некоторые арктические растения имеют апикальную меристему в нижней/средней части растения. Считается, что такая меристема возникла потому, что она выгодна в арктических условиях. [ нужна ссылка ]
Стрелять апикальные меристемы
[ редактировать ]
Апикальные меристемы побега являются источником всех надземных органов, таких как листья и цветы. Клетки на вершине апикальной меристемы побега служат стволовыми клетками для окружающей периферической области, где они быстро пролиферируют и включаются в дифференцирующиеся зачатки листьев или цветков.
Апикальная меристема побега является местом большей части эмбриогенеза цветковых растений. [ нужна ссылка ] Зачатки листьев, чашелистиков, лепестков, тычинок и завязей закладываются здесь со скоростью один за каждый интервал времени, называемый пластохроном . Именно здесь проявляются первые признаки развития цветка. Одним из этих признаков может быть потеря апикального доминирования и высвобождение спящих клеток для развития в качестве вспомогательных меристем побега у некоторых видов в пазухах зачатков, расположенных на расстоянии двух или трех от апикального купола.
Апикальная меристема побега состоит из четырех отдельных групп клеток:
- Стволовые клетки
- Непосредственные дочерние клетки стволовых клеток
- Нижележащий организационный центр
- Клетки-основатели для инициации органов в прилегающих регионах
Эти четыре отдельные зоны поддерживаются сложным сигнальным путем. У Arabidopsis thaliana 3 взаимодействующих гена CLAVATA необходимы для регулирования размера резервуара стволовых клеток в апикальной меристеме побега путем контроля скорости клеточного деления . [5] CLV1 Предполагается, что и CLV2 образуют рецепторный комплекс (семейства рецептороподобных киназ LRR которого является CLV3 ), лигандом . [6] [7] [8] CLV3 имеет некоторую гомологию с белками ESR кукурузы, при этом 14 аминокислот . короткая область из сохраняется между белками [9] [10] Белки, содержащие эти консервативные области, были сгруппированы в семейство белков CLE. [9] [10]
Было показано, что CLV1 взаимодействует с несколькими цитоплазматическими белками, которые, скорее всего, участвуют в последующей передаче сигналов . Например, было обнаружено, что комплекс CLV связан с небольшими белками, родственными ГТФазе Rho/Rac . [5] Эти белки могут выступать в качестве промежуточного звена между комплексом CLV и митоген-активируемой протеинкиназой (MAPK), которая часто участвует в сигнальных каскадах. [11] KAPP представляет собой киназу-ассоциированную протеинфосфатазу , которая, как было показано, взаимодействует с CLV1. [12] Считается, что KAPP действует как негативный регулятор CLV1, дефосфорилируя его. [12]
Другим важным геном в поддержании меристемы растений является WUSCHEL (сокращенно WUS ), который является мишенью передачи сигналов CLV в дополнение к положительной регуляции CLV, образуя таким образом петлю обратной связи. [13] WUS экспрессируется в клетках ниже стволовых клеток меристемы, и его присутствие предотвращает дифференцировку стволовых клеток. [13] CLV1 способствует клеточной дифференцировке, подавляя активность WUS за пределами центральной зоны, содержащей стволовые клетки. [5]
Функция WUS в апикальной меристеме побега связана с фитогормоном цитокинином . Цитокинин активирует гистидинкиназы , которые затем фосфорилируют белки-фосфопереносчики гистидина. [14] Впоследствии фосфатные группы переносятся на два типа регуляторов ответа Arabidopsis (ARR): ARRS типа B и ARR типа A. ARR типа B действуют как факторы транскрипции, активируя гены, расположенные ниже цитокинина , включая A-ARR. A-ARR по структуре аналогичны B-ARR; однако A-ARR не содержат ДНК-связывающих доменов, которые есть у B-ARR и которые необходимы для функционирования в качестве факторов транскрипции. [15] Следовательно, A-ARR не способствуют активации транскрипции и, конкурируя за фосфаты с белками-фосфопереносчиками, ингибируют функцию B-ARR. [16] В SAM B-ARR индуцируют экспрессию WUS , которая индуцирует идентичность стволовых клеток. [17] Затем WUS подавляет A-ARR. [18] В результате B-ARRs больше не ингибируются, вызывая устойчивую передачу сигналов цитокининов в центре апикальной меристемы побега. Вместе с сигнализацией CLAVATA эта система работает как петля отрицательной обратной связи . Передача сигналов цитокинина положительно усиливается WUS, чтобы предотвратить ингибирование передачи сигналов цитокинина, в то время как WUS способствует развитию собственного ингибитора в форме CLV3, который в конечном итоге удерживает передачу сигналов WUS и цитокинина под контролем. [19]
Корневая апикальная меристема
[ редактировать ]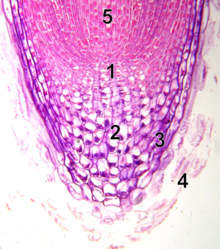
- спокойный центр
- калиптроген (живые клетки корневого чехлика)
- корневой чехлик
- отшелушивание мертвых клеток корневого чехлика
- прокамбий
В отличие от апикальной меристемы побега, апикальная меристема корня производит клетки в двух измерениях. Он содержит два пула стволовых клеток вокруг организующего центра, называемого клетками покоящегося центра (QC), и вместе производит большую часть клеток взрослого корня. [20] [21] На вершине корневая меристема покрыта корневым чехликом, который защищает и направляет траекторию ее роста. Клетки постоянно отслаиваются от внешней поверхности корневого чехлика . Клетки ПЦ характеризуются низкой митотической активностью. Имеющиеся данные свидетельствуют о том, что QC поддерживает окружающие стволовые клетки, предотвращая их дифференцировку с помощью сигналов, которые еще предстоит обнаружить. Это обеспечивает постоянный приток новых клеток в меристему, необходимый для непрерывного роста корня. Недавние результаты показывают, что КК также может выступать в качестве резервуара стволовых клеток для восполнения всего, что было потеряно или повреждено. [22] Апикальная меристема корня и структура тканей формируются у эмбриона в случае первичного корня и в новом зачатке бокового корня в случае вторичных корней.
Интеркалярная меристема
[ редактировать ]У покрытосеменных вставочные (иногда называемые базальными) меристемы встречаются в стеблях однодольных (в частности, травяных ) у основания узлов и листовых пластинок. Хвощи и вельвичия также демонстрируют вставочный рост. Интеркалярные меристемы способны к делению клеток и обеспечивают быстрый рост и возобновление роста многих однодольных растений. Интеркалярные меристемы в узлах бамбука обеспечивают быстрое удлинение стебля, а меристемы у основания большинства листовых пластинок травы позволяют поврежденным листьям быстро отрастать. Такое возобновление роста листьев трав развилось в ответ на ущерб, нанесенный травоядными животными и/или лесными пожарами.
Цветочная меристема
[ редактировать ]Когда растения начинают цвести, апикальная меристема побега преобразуется в меристему соцветия, из которой образуется цветочная меристема, из которой образуются чашелистики, лепестки, тычинки и плодолистики цветка.
В отличие от вегетативных апикальных меристем и некоторых меристем цветения, цветочные меристемы не могут продолжать расти бесконечно. Их рост ограничивается цветком определенного размера и формы. Для перехода от меристемы побега к меристеме цветка необходимы гены, идентифицирующие меристему цветка, которые одновременно определяют цветочные органы и вызывают прекращение производства стволовых клеток. AGAMOUS ( AG ) – это цветочный гомеотический ген, необходимый для терминации меристемы цветка и необходимый для правильного развития тычинок и плодолистиков . [5] AG необходим для предотвращения превращения цветочных меристем в меристемы побегов соцветия, но является тождественным геном LEAFY ( LFY ) и WUS и ограничен центром цветочной меристемы или двумя внутренними мутовками. [23] Таким образом достигается цветочная идентичность и региональная специфика. WUS активирует AG путем связывания с консенсусной последовательностью во втором интроне AG, а LFY связывается с соседними сайтами узнавания. [23] Как только AG активируется, он подавляет экспрессию WUS, что приводит к прекращению меристемы. [23]
На протяжении многих лет ученые манипулировали цветочными меристемами по экономическим причинам. Примером может служить мутантное растение табака «Мэрилендский мамонт». В 1936 году департамент сельского хозяйства Швейцарии провел с этим растением несколько научных испытаний. «Мэрилендский мамонт» необычен тем, что растет гораздо быстрее других табачных растений.
Апикальное доминирование
[ редактировать ]Апикальное доминирование – это когда одна меристема предотвращает или подавляет рост других меристем. В результате у растения будет один четко выраженный главный ствол. Например, у деревьев верхушка основного ствола несет доминантную меристему побега. Поэтому кончик ствола быстро растет и не затеняется ветвями. Если доминантную меристему отрезать, один или несколько кончиков ветвей возьмут на себя доминирование. Филиал начнет расти быстрее, и новый рост будет вертикальным. С годами ветвь может начать все больше напоминать продолжение основного ствола. Часто такое поведение наблюдается в нескольких ветвях после удаления апикальной меристемы, что приводит к густому росту.
Механизм апикального доминирования основан на ауксинах — регуляторах роста растений. Они производятся в апикальной меристеме и транспортируются к корням в камбии . Если апикальное доминирование полное, они предотвращают образование ветвей, пока апикальная меристема активна. Если доминирование неполное, будут развиваться боковые ветви. [ нужна ссылка ]
Недавние исследования апикального доминирования и контроля ветвления выявили новое семейство растительных гормонов, названное стриголактонами . Ранее было известно, что эти соединения участвуют в прорастании семян и взаимодействии с микоризными грибами , а теперь показано, что они участвуют в ингибировании ветвления. [24]
Разнообразие меристемной архитектуры
[ редактировать ]SAM содержит популяцию стволовых клеток , которые также производят боковые меристемы, пока стебель удлиняется. Оказывается, механизм регуляции количества стволовых клеток может быть эволюционно консервативным. Ген CLAVATA ответственный CLV2, за поддержание популяции стволовых клеток Arabidopsis thaliana, очень тесно связан с кукурузы геном FASCIATED EAR 2 ( FEA2 ), также участвующим в той же функции. [25] Аналогично, у риса система FON1-FON2, по-видимому, имеет тесную связь с сигнальной системой CLV у Arabidopsis thaliana . [26] Эти исследования показывают, что регуляция количества, идентичности и дифференцировки стволовых клеток может быть эволюционно консервативным механизмом у однодольных , если не у покрытосеменных . Рис также содержит еще одну генетическую систему, отличную от FON1-FON2 , которая участвует в регуляции количества стволовых клеток . [26] Этот пример подчеркивает инновации , которые постоянно происходят в живом мире.
Роль генов семейства KNOX
[ редактировать ]

Генетический скрининг выявил гены, принадлежащие к семейству KNOX , выполняющие эту функцию. Эти гены по существу поддерживают стволовые клетки в недифференцированном состоянии. Семейство KNOX претерпело значительную эволюционную диверсификацию, сохранив при этом общий механизм более или менее схожим. Члены семейства KNOX были обнаружены в таких разнообразных растениях, как Arabidopsis thaliana , рис, ячмень и томаты. KNOX-подобные гены также присутствуют в некоторых водорослях , мхах, папоротниках и голосеменных растениях . Неправильная экспрессия этих генов приводит к формированию интересных морфологических особенностей. Например, среди представителей Antirrineae только виды рода Antirhinum лишены структуры, называемой шпорой, в цветочной области. Шпора считается эволюционной инновацией , поскольку она определяет опылителя специфичность и привлекательность . Исследователи провели мутагенез транспозонов у Antirhinum majus и увидели, что некоторые вставки приводили к образованию шпор, очень похожих на шпоры у других представителей Antirhineae. , [27] что указывает на то, что потеря шпоры в диких популяциях Antirhinum majus, вероятно, могла быть эволюционным нововведением.
Семейство KNOX также принимало участие в листьев эволюции формы (более подробное обсуждение см. ниже) . В одном исследовании изучался характер экспрессии гена KNOX у A. thaliana , имеющего простые листья, и Cardamine hirsuta , растения со сложными листьями . У A. thaliana гены KNOX полностью отключаются в листьях, но у C.hirsuta экспрессия продолжается, образуя сложные листья. [28] Кроме того, было высказано предположение, что механизм действия гена KNOX консервативен для всех сосудистых растений , поскольку существует тесная корреляция между экспрессией KNOX и сложной морфологией листьев . [29]
Неопределенный рост меристем
[ редактировать ]Хотя каждое растение растет по определенному набору правил, каждый новый корень и меристема побега могут продолжать расти, пока они живы. У многих растений меристематический рост потенциально неопределенен , поэтому общая форма растения не определена заранее. Это основной рост . Первичный рост приводит к удлинению тела растения и формированию органов. Все органы растения в конечном итоге возникают в результате деления клеток апикальной меристемы с последующим расширением и дифференцировкой клеток. Первичный рост дает начало верхушечной части многих растений.
Рост азотфиксирующих корневых клубеньков на бобовых растениях, таких как соя и горох, может быть детерминантным или индетерминантным. Таким образом, соя (или фасоль и Lotus japonicus) образуют детерминированные клубеньки (сферические) с разветвленной сосудистой системой, окружающей центральную зону заражения. Часто клетки, инфицированные Rhizobium, имеют лишь небольшие вакуоли. Напротив, клубеньки на горохе, клевере и Medicago truncatula являются недетерминированными, чтобы поддерживать (по крайней мере, в течение некоторого времени) активную меристему, которая дает новые клетки для инфекции Rhizobium. Таким образом, в клубеньке существуют зоны зрелости. Инфицированные клетки обычно имеют большую вакуоль. Сосудистая система растения разветвленная и периферическая.
Клонирование
[ редактировать ]При соответствующих условиях каждая меристема побега может развиться в полноценное новое растение или клон . Такие новые растения можно вырастить из черенков побегов, содержащих апикальную меристему. Однако апикальные меристемы корня клонировать нелегко. Такое клонирование называется бесполым размножением или вегетативным размножением и широко практикуется в садоводстве для массового производства растений желаемого генотипа . Было показано, что этот процесс, известный как мериклонирование, уменьшает или устраняет вирусы, присутствующие в родительском растении у многих видов растений. [30] [31]
Размножение черенками - это еще одна форма вегетативного размножения, которая инициирует образование корней или побегов из вторичных меристематических камбиальных клеток. Это объясняет, почему базальное «ранение» побеговых черенков часто способствует образованию корней. [32]
Индуцированные меристемы
[ редактировать ]Меристемы также могут индуцироваться в корнях бобовых, таких как соя , Lotus japonicus , горох и Medicago truncatula, после заражения почвенными бактериями, обычно называемыми ризобиями . [ нужна ссылка ] Клетки внутренней или внешней коры головного мозга в так называемом «окне узелков» сразу за развивающимся кончиком корня начинают делиться. Критическим сигнальным веществом является липололигосахарид Nod -фактор , украшенный боковыми группами, обеспечивающими специфичность взаимодействия. Белки рецептора Nod-фактора NFR1 и NFR5 были клонированы из нескольких бобовых, включая Lotus japonicus , Medicago truncatula и соевые бобы ( Glycine max ). Регуляция меристем клубеньков использует регуляцию на расстоянии, известную как авторегуляция клубеньков (АОН). , локализованные в сосудистой ткани листа LRR рецепторов Этот процесс включает в себя киназы (LjHAR1, GmNARK и MtSUNN), передачу сигналов пептида CLE и взаимодействие KAPP, подобное тому, которое наблюдается в системе CLV1,2,3. регуляции узелков, LjKLAVIER также демонстрирует фенотип хотя пока неизвестно, как это связано с другими киназами рецептора AON.
Боковые меристемы
[ редактировать ]Боковые меристемы, форма вторичного роста растений, увеличивают диаметр растений. В первую очередь это наблюдается у многолетних двудольных растений, выживающих из года в год. Различают два типа латеральных меристем: сосудистый камбий и пробковый камбий.
В сосудистом камбии первичная флоэма и ксилема образуются апикальной меристемой. После этого первоначального развития латеральная меристема образует вторичную флоэму и ксилему. Они соединены тонким слоем паренхиматозных клеток, которые дифференцируются в пучок камбия. Пучок камбия делится, образуя новую вторичную флоэму и ксилему. Вслед за этим кортикальная паренхима между сосудистыми цилиндрами дифференцирует межпучковый камбий. Этот процесс повторяется для неопределенного роста. [33]
Пробковый камбий создает защитное покрытие вокруг растения снаружи. Это происходит после того, как вторичная ксилема и флоэма уже разрослись. Кортикальные паренхиматозные клетки дифференцируются в пробковый камбий вблизи эпидермиса, который образует новые клетки, называемые феллодермой и пробковыми клетками. Эти пробковые клетки непроницаемы для воды и газов из-за вещества под названием суберин, которое их покрывает. [34]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Галун, Эсра (2007). Рисунок растений: структурные и молекулярно-генетические аспекты . Мировое научное издательство. п. 333. ISBN 9789812704085
- ^ Баушер, Мари; АльмДжазири, Мондер; Вандепут, Оливье. «От первичного роста к вторичному: возникновение и развитие сосудистой системы» . Academic.oup.com . Проверено 18 марта 2023 г.
- ^ Тогнетти, Ванеса Б.; Белах, Агнешка; Хртян, Моника (октябрь 2017 г.). «Окислительно-восстановительная регуляция в месте первичного роста: перекрестные помехи ауксина, цитокинина и АФК: пластичность апикальной меристемы в ответ на стресс» . Растение, клетка и окружающая среда . 40 (11): 2586–2605. дои : 10.1111/шт.13021 . ПМИД 28708264 .
- ^ Jump up to: а б Эверт, Рэй и Сьюзен Эйххорн. Ворон Биология растений. Нью-Йорк: WH Freeman and Company, 2013. Печать.
- ^ Jump up to: а б с д Флетчер, Дж. К. (2002). «Поддержание побегов и цветочной меристемы арабидопсиса». Анну. Преподобный Плант Биол. 53 : 45–66. doi : 10.1146/annurev.arplant.53.092701.143332 . ПМИД 12221985 .
- ^ Кларк С.Е., Уильямс Р.В., Мейеровиц Э. (1997). «Ген CLAVATA1 кодирует предполагаемую рецепторную киназу, которая контролирует размер побегов и цветочной меристемы арабидопсиса» . Клетка . 89 (4): 575–85. дои : 10.1016/S0092-8674(00)80239-1 . ПМИД 9160749 . S2CID 15360609 .
- ^ Чон С., Тротошо А.Е., Кларк С. (1999). «Ген CLAVATA2 Arabidopsis кодирует рецептороподобный белок, необходимый для стабильности рецептороподобной киназы CLAVATA1» . Растительная клетка . 11 (10): 1925–33. дои : 10.1105/tpc.11.10.1925 . ПМК 144110 . ПМИД 10521522 .
- ^ Флетчер Дж. К., Брэнд Ю, член парламента, Саймон Р., Мейеровиц Э. М. (1999). «Сигнализация решений судьбы клеток с помощью CLAVATA3 в меристемах побегов Arabidopsis». Наука . 283 (5409): 1911–14. Бибкод : 1999Sci...283.1911F . дои : 10.1126/science.283.5409.1911 . ПМИД 10082464 .
- ^ Jump up to: а б Дж. Марк Кок; Шейла МакКормик (июль 2001 г.). «Большое семейство генов, гомологичных с CLAVATA3» . Физиология растений . 126 (3): 939–942. дои : 10.1104/стр.126.3.939 . ПМК 1540125 . ПМИД 11457943 .
- ^ Jump up to: а б Карстен Олкерс, Николя Гоффар, Георг Ф. Вайлер, Питер М. Грессхофф, Ульрике Матезиус и Танкред Фрики (3 января 2008 г.). «Биоинформатический анализ семейства сигнальных пептидов CLE» . Биология растений BMC . 8 :1. дои : 10.1186/1471-2229-8-1 . ПМК 2254619 . ПМИД 18171480 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Вальстер, АХ; и др. (2000). «Растительные ГТФазы: Рос в цвету». Тенденции клеточной биологии . 10 (4): 141–146. дои : 10.1016/s0962-8924(00)01728-1 . ПМИД 10740268 .
- ^ Jump up to: а б Стоун, Дж. М.; и др. (1998). «Контроль развития меристемы с помощью киназы рецептора CLAVATA1 и взаимодействий протеинфосфатазы, связанной с киназой» . Физиология растений . 117 (4): 1217–1225. дои : 10.1104/стр.117.4.1217 . ПМК 34886 . ПМИД 9701578 .
- ^ Jump up to: а б Майер, К.Ф. X; и др. (1998). «Роль WUSCHEL в регулировании судьбы стволовых клеток в меристеме побегов Arabidopsis» . Клетка . 95 (6): 805–815. дои : 10.1016/S0092-8674(00)81703-1 . ПМИД 9865698 . S2CID 18995751 .
- ^ Шин, Джен ; Хван, Ильду (сентябрь 2001 г.). «Двухкомпонентная схема передачи сигнала цитокинина арабидопсиса». Природа . 413 (6854): 383–389. Бибкод : 2001Natur.413..383H . дои : 10.1038/35096500 . ISSN 1476-4687 . ПМИД 11574878 . S2CID 4418158 .
- ^ Ломанн, Ян У.; Кибер, Джозеф Дж.; Демар, Моника; Андреас Келе; Стелинг, Сандра; Буш, Вольфганг; Дженнифер ПК; Лейбфрид, Андреа (декабрь 2005 г.). «WUSCHEL контролирует функцию меристемы путем прямой регуляции цитокинин-индуцируемых регуляторов ответа». Природа . 438 (7071): 1172–1175. Бибкод : 2005Natur.438.1172L . дои : 10.1038/nature04270 . ISSN 1476-4687 . ПМИД 16372013 . S2CID 2401801 .
- ^ Кибер, Джозеф Дж.; Экер, Джозеф Р.; Алонсо, Хосе М.; Шаллер, Г. Эрик; Мейсон, Майкл Г.; Дерюэр, Жан; Феррейра, Фернандо Х.; Хаберер, Георг; Дженнифер ПК (01 марта 2004 г.). «Регуляторы ответа арабидопсиса типа А являются частично избыточными негативными регуляторами передачи сигналов цитокининов» . Растительная клетка . 16 (3): 658–671. дои : 10.1105/tpc.018978 . ISSN 1040-4651 . ПМЦ 385279 . ПМИД 14973166 .
- ^ Юргенс, Г.; Бергер, Дж.; Майер, К.Ф.; Ло, Т. (1 января 1996 г.). «Ген WUSCHEL необходим для целостности побегов и цветочной меристемы арабидопсиса» . Разработка . 122 (1): 87–96. дои : 10.1242/dev.122.1.87 . ISSN 0950-1991 . PMID 8565856 .
- ^ Джексон, Дэвид; Саймон, Рюдигер; Дже, Бён Иль; Сомссич, Марк (15 сентября 2016 г.). «Передача сигналов CLAVATA-WUSCHEL в меристеме побега» . Разработка . 143 (18): 3238–3248. дои : 10.1242/dev.133645 . ISSN 0950-1991 . ПМИД 27624829 .
- ^ Гордон, СП; Чикармане, В.С.; Оно, К.; Мейеровиц, Э.М. (26 августа 2009 г.). «Множественные петли обратной связи через передачу сигналов цитокининов контролируют количество стволовых клеток в меристеме побега арабидопсиса» . Труды Национальной академии наук . 106 (38): 16529–16534. Бибкод : 2009PNAS..10616529G . дои : 10.1073/pnas.0908122106 . ISSN 0027-8424 . ПМЦ 2752578 . ПМИД 19717465 .
- ^ Себастьян, Хосе; Ли, Джи Ён (2013). «Корневые апикальные меристемы». ЭЛС . дои : 10.1002/9780470015902.a0020121.pub2 . ISBN 978-0470016176 .
- ^ Беннетт, Том; Шерес, Бен (2010). «Развитие корня – две меристемы по цене одной?». Актуальные темы биологии развития . 91 : 67–102. дои : 10.1016/S0070-2153(10)91003-X . ISBN 9780123809100 . ПМИД 20705179 .
- ^ Хейдстра, Ренце; Сабатини, Сабрина (2014). «Стволовые клетки растений и животных: похожие, но разные». Nature Reviews Молекулярно-клеточная биология . 15 (5): 301–12. дои : 10.1038/nrm3790 . ПМИД 24755933 . S2CID 34386672 .
- ^ Jump up to: а б с Ломанн, Ю. и др. (2001) Молекулярная связь между регуляцией стволовых клеток и формированием цветочного рисунка в клетках арабидопсиса 105: 793-803
- ^ «Разветвление: новый класс растительных гормонов подавляет образование ветвей» . Природа . 455 (7210). 11 сентября 2008 г. Проверено 30 апреля 2009 г.
- ^ Тагучи-Шиобара; Юань, Z; Хек, С; Джексон, Д; и др. (2001). «Ген fasciated Ear2 кодирует богатый лейцином повторяющийся рецептор-подобный белок, который регулирует пролиферацию меристемы побега кукурузы» . Гены и развитие . 15 (20): 2755–2766. дои : 10.1101/gad.208501 . ПМК 312812 . ПМИД 11641280 .
- ^ Jump up to: а б Сузаки Т.; Ториба, Т; Фудзимото, М; Цуцуми, Н; Китано, Х; Хирано, Хай (2006). «Сохранение и диверсификация механизма поддержания меристемы у Oryza sativa: функция гена ЦВЕТОЧНОГО ОРГАНА НОМЕР2» . Физиология растений и клеток . 47 (12): 1591–1602. дои : 10.1093/pcp/pcl025 . ПМИД 17056620 .
- ^ Гольц Ю.Ф.; Кек, Эмма Дж.; Хадсон, Эндрю (2002). «Спонтанные мутации в генах KNOX приводят к возникновению новой цветочной структуры у антирринума» . Курс. Биол . 12 (7): 515–522. Бибкод : 2002CBio...12..515G . дои : 10.1016/S0960-9822(02)00721-2 . ПМИД 11937019 . S2CID 14469173 .
- ^ Хэй и Циантис; Циантис, М (2006). «Генетическая основа различий в форме листьев между Arabidopsis thaliana и его диким родственником Cardamine hirsuta ». Нат. Жене . 38 (8): 942–947. дои : 10.1038/ng1835 . ПМИД 16823378 . S2CID 5775104 .
- ^ Бхаратан Дж. и др. (2002). «Гомологии в форме листа, выведенные из экспрессии гена KNOXI во время развития». Наука . 296 (5574): 1858–1860. Бибкод : 2002Sci...296.1858B . дои : 10.1126/science.1070343 . ПМИД 12052958 . S2CID 45069635 .
- ^ Адамс, Алекса (апрель 2013 г.). «Уничтожение вирусов из хмеля (Humulus lupulus) путем термотерапии и меристемной культуры» . Журнал садоводческих наук . 50 (2): 151–160. дои : 10.1080/00221589.1975.11514616 . Проверено 24 января 2023 г.
- ^ Алам, я; Шармин, ЮАР; Нахер, МК; Алам, MJ; Анисуззаман, М; Алам, МФ (апрель 2013 г.). «Уничтожение и обнаружение вирусов в ростках сладкого картофеля, полученных из меристемы, как недорогой вариант коммерциализации» . 3 Биотехнологии . 3 (2): 53–164. дои : 10.1007/s13205-012-0080-6 . ПМЦ 3597136 . ПМИД 8324570 .
- ^ Маккензи, Канада; Ховард, Б.Х. (1986). «Анатомическая связь между камбиальной регенерацией и закладыванием корней у раненых зимних черенков подвоя яблони M.26». Анналы ботаники . 58 (5): 649–661. doi : 10.1093/oxfordjournals.aob.a087228 .
- ^ Ниеминен, Кайса; Бломстер, Тиина; Хелариутта, Йа; Мяхёнен, Ари Пекка (январь 2015 г.). «Развитие сосудистого камбия» . Книга «Арабидопсис» . 13 : e0177. дои : 10.1199/tab.0177 . ISSN 1543-8120 . ПМЦ 4463761 . ПМИД 26078728 .
- ^ «Развитие растений II: первичный и вторичный рост | Органическая биология» . organismalbio.biosci.gatech.edu . Проверено 8 апреля 2024 г.
Источники
[ редактировать ]- Лаборатория анатомии растений Техасского университета ; лаборатория Дж. Д. Маузета. Микрофотографии растительных клеток и тканей с пояснительным текстом.
- Шуф, Хейко; Ленхард, М; Хеккер, А; Майер, К.Ф.; Юргенс, Г; Ло, Т (2000). « Меристемы побегов арабидопсиса поддерживаются регуляторной петлей между генами Clavata и Wuschel» . Клетка . 100 (6): 635–644. doi : 10.1016/S0092-8674(00)80700-X . ПМИД 10761929 . S2CID 8963007 .
- Скофилд и Мюррей (2006). Развивающаяся концепция меристемы. Молекулярная биология растений 60:v–vii.
Внешние ссылки
[ редактировать ]| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| растений Группы | |||||||||||
| Анатомия растений |
| ||||||||||
| Физиология растений Материалы | |||||||||||
| Рост растений и привычка | |||||||||||
| Воспроизведение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||