Корневой узелок

Корневые клубеньки встречаются на корнях растений бобовых , прежде всего , образующих симбиоз с азотфиксирующими бактериями . [1] В условиях ограничения азота способные растения образуют симбиотические отношения со специфичным для хозяина штаммом бактерий, известным как ризобии . [2] Этот процесс неоднократно развивался у бобовых, а также у других видов семейства Rosid . [3] К бобовым культурам относятся фасоль , горох и соевые бобы .
В корневых клубеньках бобовых культурный азот (N 2 ) из атмосферы превращается в аммиак (NH 3 ), который затем усваивается в аминокислоты (строительные блоки белков), нуклеотиды (строительные блоки ДНК и РНК , а также важная энергетическая молекула АТФ ) и другие клеточные компоненты, такие как витамины , флавоны и гормоны . [ нужна ссылка ] Их способность фиксировать газообразный азот делает бобовые идеальными сельскохозяйственными организмами, поскольку их потребность в азотных удобрениях снижается. Действительно, высокое содержание азота блокирует развитие клубеньков, поскольку от формирования симбиоза растению нет никакой пользы. Энергия для расщепления газообразного азота в клубеньке поступает из сахара, который переносится из листа (продукт фотосинтеза ). Малат как продукт распада сахарозы является прямым источником углерода для бактероида. Фиксация азота в узле очень чувствительна к кислороду. Клубеньки бобовых содержат железосодержащий белок, называемый леггемоглобином , тесно связанным с миоглобином животных , который облегчает диффузию газообразного кислорода, используемого при дыхании.
Симбиоз
[ редактировать ]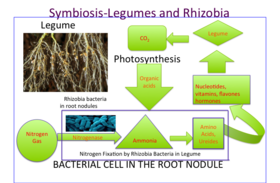
Семья бобовых
[ редактировать ]Растения, которые способствуют фиксации N2, включают семейство бобовых – Fabaceae – с такими таксонами, как кудзу , клевер , соевые бобы , люцерна , люпин , арахис и ройбуш . В клубеньках они содержат симбиотические бактерии, называемые ризобиями , которые производят соединения азота, которые помогают растению расти и конкурировать с другими растениями. Когда растение умирает, фиксированный азот высвобождается, делая его доступным для других растений, и это помогает удобрять почву . [4] [5] Подавляющее большинство бобовых имеют такую связь, но некоторые роды (например, Styphnolobium ) ее не имеют. Во многих традиционных методах ведения сельского хозяйства на полях чередуются различные типы культур, в том числе культуры, состоящие в основном или полностью из бобовых культур, таких как клевер, чтобы воспользоваться этим.
Незерновые
[ редактировать ]Хотя подавляющее большинство растений, способных образовывать азотфиксирующие корневые клубеньки, относятся к семейству бобовых Fabaceae , есть несколько исключений:
- Актиноризные растения, такие как ольха и бейберри, могут образовывать (менее сложные) азотфиксирующие клубеньки благодаря симбиотической ассоциации с Frankia бактериями . Эти растения принадлежат к 25 родам, распределенным по 8 семействам растений. [6] По подсчетам 1998 г., он включает около 200 видов и обеспечивает примерно такую же степень азотфиксации, как и ризобиальные симбиозы. Важным структурным отличием является то, что в этих симбиозах бактерии никогда не освобождаются из нити инфекции. [7]
- Parasponia , тропический род Cannabaceae, также способен взаимодействовать с ризобиями и образовывать азотфиксирующие клубеньки. Поскольку родственные растения являются актиноризными, считается, что растение «поменяло партнера» в своей эволюции. [8]
Способность фиксировать азот в этих семьях присутствует далеко не повсеместно. Например, из 122 родов розоцветных только 4 рода способны фиксировать азот. Все эти семейства относятся к порядкам Cucurbitales , Fagales и Rosales , которые вместе с Fabales образуют азотфиксирующую кладу (NFC) евросид . В этой кладе Fabales были первой ответвленной линией; таким образом, способность фиксировать азот может быть плезиоморфной и впоследствии утрачиваться у большинства потомков исходного азотфиксирующего растения; однако возможно, что основные генетические и физиологические потребности присутствовали в зачаточном состоянии у последних общих предков всех этих растений, но развились до полной функции лишь у некоторых из них:
| Семья: Генера Betulaceae : Alnus (ольха) | ...... | ...... | ...... |
| ...... |
|
Классификация
[ редактировать ]
У бобовых описаны два основных типа клубеньков: детерминантный и индетерминантный. [9]
Детерминированные клубеньки встречаются на некоторых племенах тропических бобовых, например, родов Glycine (соевые бобы), Phaseolus (фасоль обыкновенная) и Vigna . и на некоторых бобовых умеренного пояса, таких как лотос . Эти детерминированные узелки теряют меристематическую активность вскоре после возникновения, поэтому рост происходит за счет расширения клеток, в результате чего образуются зрелые узелки, имеющие сферическую форму. Другой тип детерминированных клубеньков встречается у широкого спектра трав, кустарников и деревьев, таких как арахис ( арахис ). Они всегда связаны с пазухами боковых или придаточных корней и образуются в результате заражения через трещины, в которых выходят эти корни, без использования корневых волосков . Их внутреннее строение сильно отличается от клубеньков соевого типа. [10]
Неопределенные клубеньки встречаются у большинства бобовых всех трех подсемейств как в регионах с умеренным климатом, так и в тропиках. Их можно увидеть в бобовых Faboideae , таких как Pisum (горох), Medicago (люцерна), Trifolium (клевер) и Vicia (вика), а также во всех мимозоидных бобовых, таких как акация , и в немногих цезальпиниоидных бобовых с клубеньками, таких как горох куропатки . Они получили название «индетерминантные», потому что поддерживают активную апикальную меристему , которая производит новые клетки для роста в течение жизни узелка. В результате узелок имеет, как правило, цилиндрическую форму, которая может быть сильно разветвленной. [10] Поскольку неопределенные клубеньки активно растут, они образуют зоны, разграничивающие разные стадии развития/симбиоза: [11] [12] [13]

- Зона I — активная меристема . Здесь формируется новая ткань узла, которая позже дифференцируется в другие зоны узла.
- Зона II – зона заражения . Эта зона пронизана инфекционными нитями, полными бактерий. Растительные клетки крупнее, чем в предыдущей зоне, и деление клеток прекращается.
- Межзона II–III — здесь бактерии проникли в растительные клетки, содержащие амилопласты . Они удлиняются и начинают окончательно дифференцироваться в симбиотические азотфиксирующие бактероиды .
- Зона III — зона азотфиксации . Каждая клетка в этой зоне содержит большую центральную вакуоль , а цитоплазма заполнена полностью дифференцированными бактероидами, активно фиксирующими азот . Растение снабжает эти клетки леггемоглобином , в результате чего они приобретают отчетливый розовый цвет.
- Зона IV – зона старения . Здесь происходит разложение растительных клеток и их бактероидного содержимого. Распад гемового компонента легоглобина приводит к видимому зеленению у основания узелка.
Это наиболее широко изученный тип клубеньков, но детали совершенно различны у арахиса и его родственников, а также у некоторых других важных культур, таких как люпин, у которых клубеньки образуются в результате прямого заражения ризобиями через эпидермис и где инфекционные нити никогда не образуются. . Вокруг корня растут узелки, образующие воротниковидную структуру. В этих клубеньках и в типе арахиса центральная инфицированная ткань однородна, в ней отсутствуют неинфицированные клетки, наблюдаемые в клубеньках сои и многих неопределенных типов, таких как горох и клевер.


Узелки актиноризного типа представляют собой заметно отличающиеся структуры, встречающиеся у небобовых растений. При этом типе клетки, происходящие из коры корня, образуют инфицированную ткань, а предклубеньк становится частью зрелого клубенька. Несмотря на это, казалось бы, существенное различие, такие клубеньки у бобовых можно получить в результате единственной гомеотической мутации. [14]
Узелок
[ редактировать ]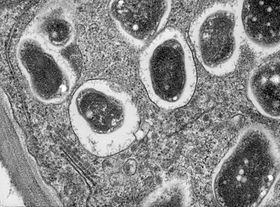

Бобовые выделяют из своих корней органические соединения в виде вторичных метаболитов , называемых флавоноидами , которые привлекают к себе ризобии, а также активируют nod- гены в бактериях, производя nod-факторы и инициируя образование клубеньков. [15] [16] Эти завивание корней факторы вызывают волос . Завивка начинается с того, что самый кончик корневого волоска завивается вокруг ризобия . Внутри кончика корня образуется небольшая трубочка, называемая инфекционной нитью, которая обеспечивает путь для проникновения ризобий в клетки эпидермиса корня, в то время как корневые волосы продолжают скручиваться. [17]
Частичного завивки можно добиться даже одним лишь фактором кивка . [16] Это было продемонстрировано путем выделения no -факторов и их применения к частям корневых волос. Корневые волоски скручивались в направлении нанесения, демонстрируя действие корневых волос, пытающихся обвиться вокруг бактерии. Даже нанесение на боковые корни вызывало скручивание. Это продемонстрировало, что именно фактор кивка , а не бактерия, вызывает стимуляцию завивки. [16]
Когда корень ощущает фактор кивка, происходит ряд биохимических и морфологических изменений: в корне запускается деление клеток , образуя узелок, а рост корневых волосков перенаправляется так, что несколько раз обвивает бактерии, пока они полностью не инкапсулируют одну из них. или более бактерий. Инкапсулированные бактерии многократно делятся, образуя микроколонии . Из этой микроколонии бактерии попадают в развивающийся узелок через инфекционную нить, которая прорастает через корневой волос в базальную часть клетки эпидермиса и далее в кору корня ; затем они окружаются симбиосомной мембраной растительного происхождения и дифференцируются в бактероиды, фиксирующие азот . [18]
Эффективное образование клубеньков происходит примерно через четыре недели после посадки культуры , при этом размер и форма клубеньков зависят от культуры. Такие культуры, как соя или арахис, будут иметь более крупные клубеньки, чем кормовые бобовые культуры, такие как красный клевер или люцерна, поскольку их потребность в азоте выше. Количество клубеньков и их внутренний цвет будут указывать на состояние фиксации азота в растении. [19]
Клубенькование контролируется множеством процессов, как внешних (жара, кислые почвы, засуха, нитраты), так и внутренних (ауторегуляция клубенькования, этилен). Ауторегуляция узелкового процесса [20] контролирует количество клубеньков на растении посредством системного процесса, затрагивающего лист. Ткань листа воспринимает ранние события клубеньков в корне посредством неизвестного химического сигнала, а затем ограничивает дальнейшее развитие клубеньков во вновь развивающейся ткани корня. Киназы рецептора богатых лейцином повторов (LRR) (NARK в соевых бобах ( Glycine max ); HAR1 в Lotus japonicus , SUNN в Medicago truncatula ) необходимы для ауторегуляции клубеньков (AON). Мутация, приводящая к потере функции этих киназ рецептора AON, приводит к супернодуляции или гипернодуляции. Часто нарушения роста корней сопровождают потерю киназной активности рецептора AON, что позволяет предположить, что рост клубеньков и развитие корней функционально связаны. Исследования механизмов образования клубеньков показали, что ген ENOD40 , кодирующий белок из 12–13 аминокислот [41], активируется во время образования клубеньков [3].
Подключение к корневой структуре
[ редактировать ]Корневые клубеньки, по-видимому, развивались трижды внутри семейства Fabaceae , но редко встречаются за пределами этого семейства. Склонность этих растений к образованию корневых клубеньков, по-видимому, связана со структурой их корней. В частности, тенденция к развитию боковых корней в ответ на абсцизовую кислоту может способствовать более позднему развитию корневых клубеньков. [21]
Узелкообразные структуры
[ редактировать ]Некоторые грибы узелковые структуры, известные как бугорчатая эктомикориза образуют на корнях своих растений-хозяев . Suillus tomentosus , например, производит эти структуры с помощью растения-хозяина сосны ложчатой ( Pinus contorta var. latifolia ). В свою очередь, было показано, что в этих структурах обитают азотфиксирующие бактерии , которые вносят значительное количество азота и позволяют соснам колонизировать участки с низким содержанием питательных веществ. [22]
Галерея
[ редактировать ] Узелки на корнях Vicia Faba .
Узелки на корнях Vicia Faba .
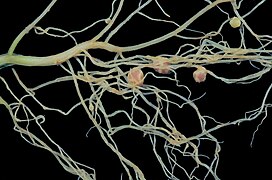 Робинии псевдоакации Клубеньки
Робинии псевдоакации Клубеньки Крупный план рассеченного Medicago корневого узелка семейства растений Fabaceae .
Крупный план рассеченного Medicago корневого узелка семейства растений Fabaceae . Fabaceae . Корневые клубеньки семейства
Fabaceae . Корневые клубеньки семейства Узелки Medicago italica .
Узелки Medicago italica .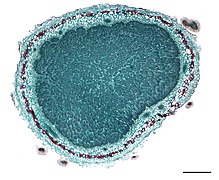 Поперечный разрез узла.
Поперечный разрез узла. Корни вигны ( Vigna unguiculata spp. ).
Корни вигны ( Vigna unguiculata spp. ).








См. также
[ редактировать ]- Корневая галловая нематода
- Ризобий
- Синоризобиум
- Брадирризобий
- Неоризобий
- Параризобий
- Общий симбиотический сигнальный путь
Ссылки
[ редактировать ]- ^ Вагнер, Стивен К. (2011). «Биологическая фиксация азота» . Знания о природном образовании . 3 (10): 15.
- ^ Ван, Ци; Ян, Шэнмин (2017). «Секретируемый хозяином антимикробный пептид усиливает симбиотическую селективность у Medicago truncatula » . ПНАС . 114 (26): 6854–6859. Бибкод : 2017PNAS..114.6854W . дои : 10.1073/pnas.1700715114 . ПМЦ 5495241 . ПМИД 28607058 .
- ^ Дойл, Джефф Дж.; Лакоу, Мелисса А. (2003). «Остальная часть айсберга. Разнообразие и эволюция бобовых в филогенетическом контексте» . Физиология растений . 131 (3): 900–910. дои : 10.1104/стр.102.018150 . ISSN 1532-2548 . ПМК 1540290 . ПМИД 12644643 .
- ^ Постгейт, Джон (1998). Азотфиксация (3-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 9780521648530 .
- ^ Смиль, Вацлав (2000). Циклы жизни: цивилизация и биосфера . Научная американская библиотека. ISBN 9780716750796 .
- ^ Доусон, Дж. О. (2008). «Экология актиноризных растений». Азотфиксирующие актиноризные симбиозы . Фиксация азота: происхождение, применение и прогресс исследований. Том. 6. Спрингер. стр. 199–234. дои : 10.1007/978-1-4020-3547-0_8 . ISBN 978-1-4020-3540-1 . S2CID 85913801 .
- ^ Дойл, Джефф Дж. (1998). «Филогенетические взгляды на клубеньки: развивающиеся взгляды на растения и симбиотические бактерии» . Тенденции в науке о растениях . 3 (12): 473–778. дои : 10.1016/S1360-1385(98)01340-5 .
- ^ Оп-ден-Кэмп, Рик; Стренг, Аренд; Де Мита, Стефан; Цао, Цинцинь; Полоне, Элиза; Лю, Вэй; Аммираджу, пристань СС; Кудрна, Дэйв; Крыло, Род; Унтергассер, Андреас; Бисселинг, Тон; Гертс, Рене (18 февраля 2011 г.). «Микоризный рецептор LysM-типа, задействованный в симбиозе ризобий у небобовых параспоний » . Наука . 331 (6019): 909–912. Бибкод : 2011Sci...331..909O . дои : 10.1126/science.1198181 . ISSN 0036-8075 . ПМИД 21205637 . S2CID 20501765 .
- ^ Креспи, Мартин; Гальвес, Сусана (1 июня 2000 г.). «Молекулярные механизмы развития корневых клубеньков». Журнал регулирования роста растений . 19 (2): 155–166. дои : 10.1007/s003440000023 . ISSN 1435-8107 . ПМИД 11038225 . S2CID 22216527 .
- ^ Jump up to: а б Срент, Джанет И. (2009). Клубенькование бобовых: глобальная перспектива . Уайли-Блэквелл. дои : 10.1002/9781444316384 . ISBN 9781444316384 .
- ^ Фуше, Фабрис; Кондороси, Ева (1 августа 2000 г.). «Регуляция клеточного цикла в ходе органогенеза клубеньков Medicago ». Молекулярная биология растений . 43 (5): 773–786. дои : 10.1023/А:1006405029600 . ISSN 1573-5028 . ПМИД 11089876 . S2CID 11658948 .
- ^ Монахан-Джованелли, Ханна; Пинедо, Каталина Аранго; Гейдж, Дэниел Дж. (2006). «Архитектура сетей инфекционных нитей при развитии корневых клубеньков, индуцированных симбиотической бактерией Sinorhizobium meliloti на Medicago truncatula » . Физиология растений . 140 (2): 661–670. дои : 10.1104/стр.105.072876 . ПМЦ 1361332 . ПМИД 16384905 .
- ^ Ван де Вельде, Виллем; Герра, Хуан Карлос Перес; Кейзер, Анник Де; Де Райк, Рит; Ромбо, Стефан; Монури, Николас; Мергерт, Питер; Кондороси, Ева; Кобуры, Марсель; Гурмахтиг, Софи (28 апреля 2006 г.). «Старение при симбиозе бобовых. Молекулярный взгляд на старение клубеньков у Medicago truncatula » . Физиология растений . 141 (2): 711–720. дои : 10.1104/стр.106.078691 . ISSN 1532-2548 . ПМЦ 1475454 . ПМИД 16648219 .
- ^ Шен, Дефэн; Сяо, Тин Тин; ван Вельзен, Робин; Куликова, Ольга; Гун, Сяоюнь; Гертс, Рене; Павловский, Катарина; Бисселинг, Тон (июнь 2020 г.). «Гомеотическая мутация превращает онтогенез клубеньков бобовых в онтогенез актиноризного типа» . Растительная клетка . 32 (6): 1868–1885. дои : 10.1105/tpc.19.00739 . ПМЦ 7268803 . ПМИД 32276984 .
- ^ Экардт, Нэнси А. (июнь 2006 г.). «Роль флавоноидов в развитии корневых клубеньков и транспорте ауксина у Medicago truncatula» . Растительная клетка . 18 (7): 1539–1540. дои : 10.1105/tpc.106.044768 . ПМК 1488913 .
- ^ Jump up to: а б с Эсселинг, Джон Дж.; Люасье, Франк GP; Эмонс, Энн Ми К. (август 2003 г.). «Завивка корневых волос, вызванная Nod Factor: непрерывный полярный рост к точке применения Nod Factor» . Физиология растений . 132 (4): 1982–1988. дои : 10.1104/стр.103.021634 . ISSN 1532-2548 . ПМК 181283 . ПМИД 12913154 .
- ^ Слончевски, Джоан; Фостер, Джон Уоткинс (2017). Микробиология: развивающаяся наука (Четвертое изд.). Нью-Йорк. ISBN 978-0393614039 . OCLC 951925510 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Мергерт, П.; Утиуми, Т.; Алунни, Б.; Эванно, Г.; Черон, А.; Катрис, О.; и др. (2006). «Эукариотический контроль бактериального клеточного цикла и дифференцировки в симбиозе ризобий-бобовых» . ПНАС . 103 (13): 5230–35. Бибкод : 2006PNAS..103.5230M . дои : 10.1073/pnas.0600912103 . ISSN 1091-6490 . ПМЦ 1458823 . ПМИД 16547129 . Онлайн.
- ^ Аджей, М.Б. «Азотфиксация и инокуляция кормовых бобовых» (PDF) . Университет Флориды . Архивировано из оригинала (PDF) 2 декабря 2016 года . Проверено 1 декабря 2016 г.
- ^ Рид, Делавэр; Фергюсон, Би Джей; Хаяши, С; Лин, Ю.Х.; Грессхофф, премьер-министр (октябрь 2011 г.). «Молекулярные механизмы, контролирующие ауторегуляцию клубеньков в бобовых» . Анналы ботаники . 108 (5): 789–95. дои : 10.1093/aob/mcr205 . ПМК 3177682 . ПМИД 21856632 .
- ^ Лян, Ян; Харрис, Жанна М. (2005). «Реакция разветвления корней на абсцизовую кислоту коррелирует с образованием клубеньков как у бобовых, так и у небобовых растений» . Американский журнал ботаники . 92 (10): 1675–1683. дои : 10.3732/ajb.92.10.1675 . ISSN 0002-9122 . ПМИД 21646084 .
- ^ Пол, ЛР; Чепмен, БК; Чанвей, CP (2007). «Азотфиксация, связанная с Suillus tomentosus Tuberculate Ectomycorizae на Pinus contorta var. latifolia » . Анналы ботаники . 99 (6): 1101–1109. дои : 10.1093/aob/mcm061 . ПМЦ 3243579 . ПМИД 17468111 .
Внешние ссылки
[ редактировать ]- Корневые клубеньки бобовых в веб-проекте «Древо жизни»
- Видео и комментарий о клубеньках белого клевера
| Базы данных органов управления : Национальные |
|---|