Общий сигнальный путь симбиоза


Общий сигнальный путь симбиоза ( CSSP ) представляет собой сигнальный каскад у растений, который позволяет им взаимодействовать с симбиотическими микробами. Это соответствует наследственному пути, который растения используют для взаимодействия с арбускулярными микоризными грибами (АМФ) . Он известен как «общий», потому что различные эволюционно более молодые симбиозы также используют этот путь, особенно симбиоз корневых клубеньков с азотфиксирующими бактериями- ризобиями . Этот путь активируется как восприятием Nod-фактора (для ризобий клубенькообразующих ) , так и восприятием Myc-фактора , высвобождаемого из АМФ. Этот путь отличается от путей распознавания патогенов , но может иметь некоторые общие рецепторы, участвующие как в распознавании патогенов, так и в CSSP. Недавняя работа [ 1 ] Кевин Коуп и его коллеги показали, что эктомикоризы (другой тип микоризы ) также используют компоненты CSSP, такие как распознавание Myc-фактора.
требует Колонизация AMF следующей цепочки [ 2 ] событий, которые можно условно разделить на следующие этапы:
1: Предварительная контактная сигнализация
2: CSSP
2: А: Восприятие
2: Б: Передача
2: С: Транскрипция
3: Программа размещения
Контур
[ редактировать ]Чтобы точно распознать инфекционную нить другого вида организма и установить взаимовыгодную ассоциацию, требуется надежная передача сигналов. [ 3 ] AM-грибы также являются жирных кислот ауксотрофами ; [ 4 ] [ 2 ] поэтому они зависят от растений в поставке жирных кислот. [ 5 ]
При пресимбиотической передаче сигналов растения и AMF выделяют химические факторы в свое окружение, чтобы партнеры могли узнавать и находить друг друга. [ 6 ] играют Корневые выделения растений роль в сложном микробном взаимодействии. [ 7 ] путем высвобождения различных соединений, [ 7 ] [ 8 ] [ 9 ] среди которых стриголактон , способствующий как колонизации АМФ, так и инфицированию патогеном. был идентифицирован [ 8 ]
Фосфатное голодание у растений вызывает выработку стриголактона , а также колонизацию АМФ. [ 8 ] Растения выделяют стриголактон, класс растительных гормонов на основе каротиноидов, который также привлекает грибных симбионтов и стимулирует окислительный метаболизм грибов наряду с ростом и ветвлением грибкового партнера. [ 2 ] Стриголактон способствует ветвлению гиф в прорастающих спорах AMF и облегчает колонизацию. [ 9 ] [ 10 ]
Общий сигнальный путь симбиоза назван так потому, что он имеет общие компоненты как для грибного симбиоза, так и для ризобиального симбиоза. Общий сигнальный путь, вероятно, развился, когда существующий путь арбускулярной микоризы был использован ризобиями. [ 2 ] [ 11 ]
Восприятие происходит, когда растение обнаруживает грибковый фактор Myc. Факторы Myc сравнимы с ризобиальными nod-факторами . была раскрыта химическая природа некоторых Myc-факторов: Myc , действуют ( липохитоолигосахаридов - LCOs ( Myc - Недавно ) и хитоолигосахаридов COs ) которые как симбиотический сигнал. [ 10 ] [ 12 ] [ 13 ]
Присутствие стриголактона усиливает продукцию Myc- CO AMF. [ 10 ]
Рецептор Myc-фактора ( MFR ) все еще остается предполагаемым. Однако белок под названием DMI2 (или SYMRK ) играет важную роль в процессе восприятия и считается корецептором MFR. Другие растения, такие как рис, могут использовать другие механизмы, используя Os CERK1 и Os CEBiP для обнаружения олигомеров хитина. [ 2 ] [ 14 ] [ 15 ] Однако недавняя работа показала, что рис SYMRK необходим для симбиоза AM. [ 16 ]
Передача происходит, когда сигнал после обнаружения передается в ядро растения. Этот процесс опосредован двумя нуклеопоринами NUP85 и NUP133 . [ 11 ] Альтернативно, другая гипотеза гласит, что редуктаза HMG-CoA активируется при восприятии, которая затем преобразует HMG-CoA в мевалонат. Этот мевалонат действует как вторичный мессенджер и активирует ядерный катионный канал K+ ( DMI-1 или Pollux ). [ 2 ] [ 17 ] Стадия передачи заканчивается созданием «кальциевого всплеска» в ядре. [ 18 ]
Стадия транскрипции начинается, когда кальций- и кальмодулин-зависимая киназа ( CCaMK) . активируется [ 2 ] Эта киназа стимулирует целевой белок CYCLOPS . [ 2 ] CCaMK и CYCLOPS, вероятно, образуют комплекс, который вместе с белком DELLA регулирует экспрессию гена RAM1 ( уменьшенная арбускулярная микориза1 ). [ 2 ]
Процесс аккомодации включает обширное ремоделирование корковых клеток хозяина. Это включает инвагинацию плазмалеммы хозяина, пролиферацию эндоплазматического ретикулума , аппарата Гольджи , сети транс-Гольджи и секреторных пузырьков . Пластиды размножаются и образуют «стромулы». Вакуоли также подвергаются обширной реорганизации. [ 11 ]
 |
 |
|---|
Предконтактная сигнализация
[ редактировать ]Химическая передача сигналов начинается до того, как два симбионта вступают в контакт. Со стороны растения-хозяина оно синтезирует и выделяет ряд фитогормонов на основе каротиноидов, называемых стриголактонами. [ 2 ] Они имеют консервативную структуру трициклического лактона, также известную как кольца ABC . [ 19 ] Биосинтез стриголактона происходит преимущественно в пластидах, [ 20 ] где D27 ( DWARF 27 риса ; ортолог ATD27 Arabidopsis ), железосвязывающая бета-каротин-изомераза, действует выше биосинтеза стриголактона. [ 20 ] Затем ферменты диоксигеназы каротиноидного расщепления CCD7 и CCD8 модифицируют структуру, которая имеет следующие ортологи:
| Имя гена | Локализация | функция | Ортолог риса | Ортолог гороха | Ортолог петунии | Ортолог арабидопсиса |
|---|---|---|---|---|---|---|
| ПЗС7 | Пластидные белки | участвует в биосинтезе стриголактона | Д17/ХТД1 | RMS5 | ПАПА3 | МАКС3 |
| ПЗС8 | Пластидные белки | участвует в биосинтезе стриголактона | Д10 | среднеквадратичное значение1 | ПАПА4 | МАКС4 |
| Альфа/бета-складчатая гидролаза | Ядерные белки | участвует в восприятии стриголактона | Д14 | среднеквадратичное значение3 | ПАПА2 | ? |
Сообщается, что альфа/бета-гидролаза D3 , а также D14L ( D14-Like ) (более поздний имеет ортолог Arabidopsis, KAI2 или KARRIKIN INSENSITIVE-2 ) играют важную роль в микоризном симбиозе. [ 21 ] в частности, D3 , D14 и D14L локализованы в ядре. [ 2 ]
NOPE1 или « NO PERCEPTION 1» — это белок-переносчик риса ( Oryza sativa ) и кукурузы ( Zea mays ), который также необходим на стадии прайминга для колонизации грибком. NOPE1 является членом семейства транспортных белков Major Facilitator Super, способных транспортировать N-ацетилглюкозамин. Поскольку корневые экссудаты мутантов nope1 не вызывают транскрипционных ответов у грибов, вполне вероятно, что NOPE1 секретирует что-то (еще не охарактеризованное), что способствует грибковой реакции. [ 22 ]
Восприятие
[ редактировать ]
Существует два основных типа корневого симбиоза; один - это симбиоз корневых клубеньков с ризобиями ( RN-тип ), а другой - арбускулярная микориза ( AM-тип ). Между этими двумя путями присутствуют общие гены. [ 23 ] эти ключевые общие компоненты образуют путь общего симбиоза (CSP или CSSP). [ 23 ] Было высказано предположение, что симбиоз RN произошел от симбиоза AM. [ 11 ] Восприятие присутствия грибкового симбионта происходит главным образом через химические выделения грибов, обычно называемые Myc-факторами. Рецепторы Myc-факторов еще не идентифицированы. Однако DMI2/SYMRK , вероятно, действует как корецептор рецептора фактора Myc (MFR). Материалами, секретируемыми АМ-грибами, имеющими отношение к симбиозу, являются Myc -LCOs , Myc- COs , N-ацетилглюкозамин. [ 2 ] [ 24 ]
| Myc-фактор | Растительный белок, на который он в основном действует |
|---|---|
| Myc -LCO | LYS11 у Lotus japonicus |
| Короткоцепочечные олигомеры хитина ( COs ) | Ос CERK1 и Ос CEBiP в рисе |
| N-ацетилглюкозамин | NOPE-1 в кукурузе |
Грибковые молекулы, вызывающие CSSP
[ редактировать ]Myc -LCO (липохитоолигосахариды)
[ редактировать ]Подобно ризобиальным LCO (факторы Nod); Myc - LCO играют важную роль на стадии восприятия. собой своего рода секретируемые соединения AM-грибов, в основном ( липохитоолигосахаридов Они представляют смеси LCO Myc - ) . У Lotus japonicus LYS11 , рецептор LCO , экспрессируется в клетках коры корня, связанных с внутрирадикальными колонизациями арбускулярных микоризных грибов. [ 24 ]
Короткоцепочечные олигомеры хитина ( Myc- COs)
[ редактировать ]Растения-хозяева AM демонстрируют симбиотически активируемые волны кальция при воздействии короткоцепочечных олигомеров хитина . Сообщалось, что производство этих молекул AM-грибом Rhizophagus нерегулярной сильно стимулируется при воздействии стриголактонов. [ 2 ] Это говорит о том, что растения выделяют стриголактоны, и в ответ гриб увеличивает количество олигомеров хитина с короткой цепью, что, в свою очередь, вызывает реакцию растения на приспособление к грибу. Считается, что лизиновый мотивный домен Os CERK1 и Os CEBiP участвует в восприятии короткоцепочечных олигомеров хитина. [ 2 ]
NOPE-1 – транспортер (описан выше). NOPE-1 также демонстрирует сильную активность поглощения N-ацетилглюкозамина и, как полагают, связан с распознаванием присутствия грибкового симбионта. [ 2 ]
Предполагается, что некоторые растительные белки распознают Myc-факторы, и лизинового мотива Os CERK1 риса ( LysM ), является одной из них. киназа, подобная рецептору [ 15 ]
Рецепторы клеточной поверхности
[ редактировать ]
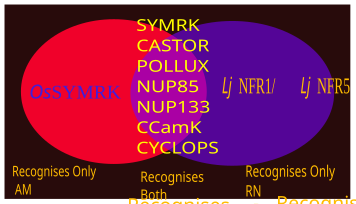
Существует множество семейств рецепторов распознавания образов и корецепторов, участвующих в распознавании микробных патогенов и симбионтов. Некоторыми из соответствующих семейств, участвующих в CSSP, являются мембраносвязанные LysM ( LYM ), растворимые рецепторы LysM, подобные белку, LYK (рецепторы LysM с активным киназным доменом), LYR (белки LysM с неактивным киназным доменом) и т. д.
По-видимому, разные комбинации рецепторов LYK и LYR воспринимают и генерируют дифференциальные сигналы, например, некоторые комбинации генерируют сигнал распознавания патогена, тогда как некоторые комбинации генерируют симбиотические сигналы. [ 25 ] [ 26 ] [ 27 ] [ 28 ]
Рецептоподобные киназы (RLK)
[ редактировать ]DMI2/SYMRK представляет собой рецептороподобную киназу, важный белок в восприятии сигналов эндосимбиоза, обнаруженный у нескольких растений ( Mt- DMI2 или Mt- NORK у Medicago trancatula ; Lj -SYMRK у Lotus japonicas; Ps -SYM19 у Pisum sativum; Os SYMRK). в Райсе). Os SYMRK не имеет N-концевого домена и регулирует исключительно АМ-симбиоз (не участвует в RN-симбиозе). [ 29 ] Примечательно, что было обнаружено, что ген, индуцируемый Nod-фактором, MtENOD11 , активируется в присутствии экссудатов АМФ; об этом явлении известно немного. [ 30 ] [ 31 ]
Киназа, подобная рецептору LysM
[ редактировать ]Рецепторно-подобные киназы Lysin Motif (LysM) представляют собой подсемейство, связанное с мембраносвязанными рецептороподобными киназами (RLK) с внеклеточной областью, состоящей из 3 лизиновых мотивов . У них есть несколько важных ортологов у разных растений, которые различаются по своим функциям . У одних видов растений они участвуют в симбиозе АМ, у других — нет. Помидор ( Solanum lycopersicum ), небобовый эвдикот , также имеет аналогичный рецептор LysM, Sl LYK10, который способствует симбиозу AM. Есть некоторые корецепторы рецептора Myc-фактора, а именно Os CEBiP в рисе, мембранный белок LysM может функционировать как корецептор Os CERK1 , но участвует в другом пути. [ 32 ] [ 33 ] [ 34 ]
Большинство этих киназ представляют собой серин/треониновые киназы , некоторые — тирозинкиназы . [ 27 ] Кроме того, они представляют собой трансмембранные белки типа 1 , что указывает на то, что их N-концевой домен направлен наружу клетки, а С-концевой домен — внутрь клетки. [ 27 ]

| Медикаго усеченный | Лотос японский | Растение гороха
(вероятно) |
Чернослива персидская | Арабидопсис Таллиана | Брассика рапа | Пасленум томатный
(Помидор) |
Брахиподий дистахион | Ориза сатива
(Рис) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Лизиновый мотив
Рецептор-подобная киназа и Рецептор лизинового мотива, подобный белку |
КЛЕЙ | LYMI | ЛИМ1 | ПП LYM1 | В LYM1
В LYM3 |
Сл LYM1 | Д-р ЛИМ1
Д-р ЛИМ3 |
Ось LYP6
Ос LYP5, Ос LYP4 | |||||
| LYMII | ЛИМ2 | ПП LYM3
ПП LYM2 |
В ЛИМ2 | Сл LYM3
Сл LYM2 |
Д-р ЛИМ2
Д-р ЛИМ4 |
ЦЭБиП
ОС LYP3 | |||||||
| ЛИРА | ЛИР 1 | ЛИРИЯ | Гора НФП
Гора LYR1 |
ЖЖ НФР5
ЖЖ LYS11 |
ПП LYR1 | Сл Lyk10 | BD LYR1 | Кость NFR5 | |||||
| ТЕКСТ | Гора LYR8 | ПП LYR2 | Сл LYK9 | Д-р ЛИР2 | |||||||||
| ЛИР 2 | ЛИРИЯ | Гора LYR10 | ЖЖ LYS16 | ПП LYR6 | Это LYK2 | Сл LYK2 | |||||||
| ЛИРИИБ | МтЛИР9 LYR9 | ЖЖ LYS15 | ПП LYR7 | Сл LYK15 | |||||||||
| ЛИР 3 | ЛИРИЯ | МтЛИР3 LYR3 | ЖЖ LYS12 | ПП LYR3 | Это LYK4 | Сл LYK4 | BD LYR4 | ОС ЛИК6 | |||||
| ЛИРИИИБ | МтЛИР2 LYR2 | ПП LYR4 | Сл LYK7
Сл LYK6 |
||||||||||
| ЛИРИКА | Гора LYR4
МтЛИР7 LYR7 |
ЖЖ LYS13
ЖЖ LYS14 |
В LYK5 | Том LYR3 | Ос LYK3
Ос LYK2, Ос LYK4 | ||||||||
| ЛИР 4 | ЛИРИВ | Гора LYR5
Гора LYR6 |
ЖЖ LYS20 | ПП LYR5 | |||||||||
| КАЖЕТСЯ | КЛЮЧ | ЛИК1, ЛИК4, ЛИК5, ЛИК6, ЛИК7, ЛИК2, ЛИК3, ЛИК9, ЛИК8 | ЖЖ LYS2
Lj LYS1, Lj NFR1, Lj LYS6, Lj LYS7 |
ПП LYK2
ПП Lyk1 |
В LYK1/
В CERK1 |
Сл LYK13
Сл LYK1/ Сл Bti9, Сл LYK12, Сл LYK11 |
Бдлык1 LYK1 | ОС CERK1 | |||||
| ЛИКИЙ | ЛИК10 | ЖЖ LYS3/
ЭПР3 |
ПП LYK3
ПП LYK4 |
||||||||||
| ЛИКИЙ | ПП LYK5 | Это LYK3 | Сл LYK3 | Д- ЛИК3 | |||||||||
| Рецептор, подобный киназе | РЛК | Мт- ДМИ2/
Мт- ВОЗ |
Lj- СИМРК | Ps-SYM19 | Ос СИМРК | ||||||||
Передача инфекции
[ редактировать ]Передача сигнальных каскадов в ядро недостаточно изучена. Однако эта передача включает в себя передачу сообщения до ядерной мембраны и генерацию кальциевой волны. [ 35 ] Некоторые элементы, участвующие в этом процессе:
Нуклеопорины
[ редактировать ]Lotus japonicus Нуклеопорины Lj NUP85 и Lj NUP133 потенциально играют роль в передаче сигнала. [ 36 ] Lj -NENA – еще один важный нуклеопорин, играющий роль в симбиозе АМ. [ 29 ]
HMGR и мевалонат.
[ редактировать ]Было высказано предположение, что фермент 3-гидрокси-3-метилглутарил-КоА-редуктаза ( HMG-CoA-редуктаза или HMGR ) может играть потенциальную роль на стадии передачи. Фермент активируется SYMRK/DMI2 и образует мевалонат. [ 37 ] [ 38 ] Этот мевалонат действует как вторичный мессенджер и активирует ядерный калиевый канал, DMI1 или POLLUX . [ 38 ]
| Ядерная оболочка Белок | Функция | Рис | Лотос японский | Медикаго усеченный | Писум |
| CNGC15 | Циклически-нуклеотидный кальциевый канал | Мт -CNGC15 | |||
| Кастор | Катионный канал калия | -Кастор | ЖЖ - Кастор | ||
| ПОЛЛЮКС или ДМИ1 | Катионный канал калия | Ос ПОЛЛУКС | Элджей Поллюкс | Мт- ДМИ1 | Ps- SYM8 |
Катионные каналы ядерной мембраны.
[ редактировать ]Ядерный кальциевый канал CNGC15 , который представляет собой ионный канал, управляемый циклическими нуклеотидами; опосредует симбиотический ядерный Ca 2+ приток, и ему противостоит K + отток по DMI1. [ 37 ]
Транскрипция
[ редактировать ]| Белок | Функция | Название завода | |||
| Рис | Лотос японский | Медикаго Трункатула | Растение гороха | ||
| CCamK | Кальций-кальмодулин-зависимая киназа, играющая роль в симбиозе АМФ | Ос- DMI3 или
Ос -ССаМК |
Lj- CCaMK | Мт- ДМИ3 | Ps- SYM9 |
| ЦИКЛОП | Спиральный домен, содержащий белки, которые реагируют на CCamK и способствуют симбиозу AMF. | Ос -ЦИКЛОП | Lj- ЦИКЛОП | Мт- ИПД3 | Ps- SYM33 |
| ОТ | Содействие симбиозу AMF | Ос- SLR1 | Гора- DELLA1
Гора -DELLA2 |
Ps- Лос-Анджелес
Ps- CRY | |
Кальмодулин — широко распространенный регуляторный белок, функционирующий совместно с Са. 2+ в различных биологических процессах. В передаче сигналов AM-симбиоза он модулирует CCaMK . [ 37 ] CCaMK или DMI3 представляет собой кальций- и кальмодулин-зависимую киназу ( CCaMK ), которая считается ключевым декодером Ca. 2+ колебания и важный регуляторный белок-киназа. Ядерный кальций 2+ добавление способствует связыванию Ca 2+ кальмодулин с CCaMK . [ 37 ] Связывание Ca 2+ кальмодулин с CCaMK вызывает конформационные изменения CCaMK , которые стимулируют белок-мишень CYCLOPS, имеющий разные ортологи. [ 37 ] ЦИКЛОП — это спиральный домен, содержащий белок. [ 37 ] возможно, образует комплекс с CCaMK [ 37 ] который работает вместе с белками DELLA . Белки DELLA представляют собой разновидность белка GRAS -домена, первоначально идентифицированного как репрессоры сигнального пути гиббереллина, однако теперь видно, что DELLA участвует во многих сигнальных путях. [ 40 ] есть два У Medicago trancatula и Pisum sativum белка DELLA , которые играют роль в симбиозе, тогда как у риса только один белок DELLA. эту задачу выполняет [ 37 ] Уменьшение арбускулярной микоризы или RAM1 [ 37 ] это ГРАС [ 41 ] белок, ген которого напрямую регулируется DELLA и CCaMK/CYCLOPS . [ 37 ] С помощью анализов иммунопреципитации хроматина было показано, что RAM1 связывается с RAM2 . промотором гена [ 37 ] RAM1 также регулирует многие гены растений, которые участвуют в аккомодации AMF.
Некоторые белки GRAS играют роль в симбиозе AM, но эти роли еще не до конца изучены. К ним относятся RAM1 , RAD1 (ТРЕБУЕТСЯ ДЛЯ РАЗВИТИЯ АРБУСКУЛА 1 ), MIG1 (GRAS, ВЫЗВАННЫЙ МИКОРИЗОЙ 1 ), NSP1 и NSP2 . [ 42 ] Считается, что гены транскрипционных факторов WRKY играют очень важную роль в установлении микоризного симбиоза и, возможно, действуют посредством регуляции защитных генов растений. [ 43 ]
Программа проживания
[ редактировать ]Клетки коры корня претерпевают важные изменения, чтобы приспособиться к грибному эндосимбионту. а также перестроить цитоплазму растительной клетки . Необходимо сформировать аппарат предварительного проникновения (PPA) во внешних слоях клеток и периарбускулярную мембрану, окружающую арбускулы во внутренних слоях клеток, [ 36 ] вакуоль уменьшается в размерах, ядро и ядрышко увеличиваются в размерах, а хроматин деконденсируется, что указывает на повышенную транскрипционную активность. [ 36 ] Пластиды размножаются и остаются связанными со «стромулюсом». [ 36 ] Более того, было высказано предположение, что апопластный продольный рост гиф, вероятно, регулируется растительными генами, такими как taci1 и CDPK1 . [ 44 ]
Гены и белки играют роль в программе аккомодации
[ редактировать ]Хотя были идентифицированы различные белки, которые могут играть роль в том, как происходит этот процесс аккомодации, подробный сигнальный каскад до конца не изучен. Некоторые из белков и механизмов, участвующих в отложении на периарбускулярной мембране, включают комплекс EXOCYST, субъединицу EXO70 , специфичный для симбиоза вариант сплайсинга SYP132 , VAPYRIN и два варианта VAMP721 . [ 37 ] Растительные ферменты FatM и RAM2 [ 45 ] и ABC-транспортер STR/STR2 предположительно участвуют в синтезе и доставке липидного β-моноацилглицерина 16:0 к АМ-грибам. [ 45 ] [ 46 ] Недавно обнаруженные киназы, которые регулируют программу аккомодации AMF, включают ADK1 , [ 47 ] AMK8 , AMK24 , [ 48 ] АРК1 [ 49 ] и АРК2 . [ 50 ]
Белковый состав периарбускулярной мембраны сильно отличается от плазматической мембраны. Он включает в себя некоторые специальные переносчики, такие как переносчики фосфатов ( Mt -PT4 , Os -PT11 , Os -PT13 ) и переносчики аммония ( Mt -AMT2 и 3 ). Он также включает транспортеры ABC, такие как STR/STR2, предположительно участвующие в транспорте липидов. [ 37 ] [ 51 ]
Эволюционное значение
[ редактировать ]АМ-грибы и растения эволюционировали совместно и развили очень сложное взаимодействие, которое позволяет растению приспосабливаться к АМ-грибу-хозяину. [ 52 ] [ 53 ] [ 54 ] Было высказано предположение, что симбиоз RN произошел от симбиоза AM. [ 36 ] [ 29 ]
См. также
[ редактировать ]- Рецептор тирозинкиназы
- Серин-треонин киназа
- Рецепторы распознавания образов
- Моноглицериды
- Стриголактон
- Интеллект растений
- Передача сигналов ячейки
- Преобразование сигнала
- Молекулярная картина, связанная с патогеном
- Молекулярный паттерн, связанный с микробами
- мутуализм
- Сигнализация с тележками
- Мевалонатный путь
Ссылки
[ редактировать ]- ^ Коуп, Кевин Р.; Баскюль, Аделина; Ирвинг, Томас Б.; Венкатешваран, Мутусубраманиан; Маэда, Джунко; Гарсия, Кевин; Раш, Томас А.; Ма, Кэтлин; Лаббе, Джесси; Джоуди, Сара; Штайгервальд, Эдвард; Сецке, Джонатан; Фунг, Эммелин; Шнелл, Кимберли Г.; Ван, Юньцянь; Шлейф, Натаниэль; Бюкинг, Хайке; Штраус, Стивен Х.; Майе, Фабьен; Джаргит, Патрисия; Бекар, Гийом; Пуэх-Пажес, Вирджиния; Ане, Жан-Мишель (октябрь 2019 г.). «Эктомикоризный гриб Laccaria bicolor продуцирует липохитоолигосахариды и использует общий путь симбиоза для колонизации корней Populus» . Растительная клетка . 31 (10): 2386–2410. дои : 10.1105/tpc.18.00676 . ПМК 6790088 . ПМИД 31416823 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Маклин, Эллисон М.; Браво, Армандо; Харрисон, Мария Дж. (октябрь 2017 г.). «Сигнальные сигналы растений и метаболические пути, обеспечивающие арбускулярный микоризный симбиоз» . Растительная клетка . 29 (10): 2319–2335. дои : 10.1105/tpc.17.00555 . ПМК 5940448 . ПМИД 28855333 .
- ^ Бармен Ягнасен; Саманта, Авик; Саха, Бабита; Датта, Сирадж (декабрь 2016 г.). «Микориза: старейшая ассоциация между растениями и грибами». Резонанс . 21 (12): 1093–1104. дои : 10.1007/ s12045-016-0421-6 ISSN 0971-8044 . S2CID 109932925 .
- ^ Лугинбюль, Леони Х.; Менар, Уильям Н.; Куруп, Смит; Ван Эрп, Гарри; Радхакришнан, Гуру В.; Брейкспир, Эндрю; Олдройд, Джайлз ЭД; Истмонд, Питер Дж. (16 июня 2017 г.). «Жирные кислоты, синтезируемые растением-хозяином в арбускулярных микоризных грибах» . Наука 356 (6343): 1175–1178. дои : 10.1126/science.aan0081 . ПМИД 28596311 . S2CID 206658467 .
- ^ Браво, Армандо; Брэндс, Матиас; Вевер, Вера; Дорманн, Питер; Харрисон, Мария Дж. (июнь 2017 г.). «Специфические для арбускулярной микоризы ферменты FatM и RAM 2 тонко настраивают биосинтез липидов, способствуя развитию арбускулярной микоризы» . Новый фитолог . 214 (4): 1631–1645. дои : 10.1111/nph.14533 . ПМИД 28380681 . S2CID 3481181 .
- ^ Пашковский, Ута (октябрь 2006 г.). «Путешествие по передаче сигналов в арбускулярных микоризных симбиозах, 2006 г.» . Новый фитолог . 172 (1): 35–46. дои : 10.1111/j.1469-8137.2006.01840.x . ПМИД 16945087 .
- ^ Jump up to: а б Байс, Харш П.; Вейр, Тиффани Л.; Перри, Лаура Г.; Гилрой, Саймон; Виванко, Хорхе М. (1 июня 2006 г.). «Роль корневых выделений во взаимодействии ризосферы с растениями и другими организмами». Ежегодный обзор биологии растений . 57 (1): 233–266. doi : 10.1146/annurev.arplant.57.032905.105159 . ПМИД 16669762 .
- ^ Jump up to: а б с Бадри, Даякар В.; Виванко, Хорхе М. (июнь 2009 г.). «Регуляция и функция корневых экссудатов» . Растение, клетка и окружающая среда . 32 (6): 666–681. дои : 10.1111/j.1365-3040.2009.01926.x . ПМИД 19143988 .
- ^ Jump up to: а б Де-ла-Пенья, Клелия; Бадри, Даякар В.; Лойола-Варгас, Виктор М. (2012). «Выделения корней растений и их взаимодействие с соседями». Секреции и экссудаты в биологических системах . Сигнализация и связь у растений. Том. 12. стр. 1–26. дои : 10.1007/978-3-642-23047-9_1 . ISBN 978-3-642-23046-2 .
- ^ Jump up to: а б с Жанр, Андреа; Шабо, Мирей; Бальзерг, Колин; Пуэх-Пажес, Вирджиния; Новеро, Мара; Рей, Томас; Фурнье, Жоэль; Рошанж, Сойзич; Бекар, Гийом; Бонфанте, Паола; Баркер, Дэвид Г. (апрель 2013 г.). «Короткоцепочечные олигомеры хитина из арбускулярных микоризных грибов вызывают всплеск ядерного C a 2+ в корнях Medicago truncatula, и их производство усиливается стриголактоном» . Новый фитолог . 198 (1): 190–202. дои : 10.1111/nph.12146 . hdl : 2318/134858 . ПМИД 23384011 . S2CID 34009711 .
- ^ Jump up to: а б с д Бонфанте, Паола; Жанр, Андреа (27 июля 2010 г.). «Механизмы, лежащие в основе полезных взаимодействий растений и грибов в микоризном симбиозе» . Природные коммуникации . 1 (1): 48. Бибкод : 2010NatCo...1...48B . дои : 10.1038/ncomms1046 . hdl : 2318/77771 . ПМИД 20975705 .
- ^ Майе, Фабьен; Пуансо, Верена; Андре, Оливье; Пуэх-Пажес, Вирджиния; Хауи, Александра; Генье, Моник; Кромер, Лоуренс; Жироде, Дельфин; Форми, Дэмиен; Нибель, Андреас; Мартинес, Эдуардо Андрес; Дригес, Хьюг; Бекар, Гийом; Денарие, Жан (2011). «Симбиотические сигналы грибковых липохитоолигосахаридов при арбускулярной микоризе». Природа . 469 (7328): 58–63. Бибкод : 2011Природа.469...58М . дои : 10.1038/nature09622 . ПМИД 21209659 . S2CID 4373531 .
- ^ Сун, Чонхо; Миллер, Дж. Бенджамин; Гранквист, Эмма; Уайли-Карлил, Одри; Гоббато, Энрико; Майе, Фабьен; Коттаз, Сильвен; Самаин, Эрик; Венкатешваран, Мутусубраманиан; Форт, Себастьян; Моррис, Ричард Дж.; Энн, Джон Майкл; Денарий, Джон; Олдройд, Джайлз, Эд (9 апреля 2015 г.). «Активация передачи сигналов симбиоза арбускулярными микоризными грибами в бобовых и рисе» . Растительная клетка 27 (3): 823–838. дои : 10.1105/tpc.114.131326 . ПМЦ 4558648 . ПМИД 25724637 .
- ^ Жанр, А.; Бонфанте, П. (февраль 2020 г.). «Рисовый рецептор сигналов микоризных грибов открывает новые возможности для развития устойчивых методов ведения сельского хозяйства» . Молекулярный завод . 13 (2): 181–183. дои : 10.1016/j.molp.2020.01.009 . hdl : 2318/1726475 . ПМИД 31981734 . S2CID 210912425 .
- ^ Jump up to: а б Мията, Козаки, Юсуке; Одзава, Кэндзиро; Асамидзу, Окабе, Ёсуке; Миямото, Аяно; Акияма, Коки; Ёко, Наото, Накагава, Томоми (ноябрь); 2014). , регулирует как хитиновый иммунитет, так и арбускулярный микоризный симбиоз риса» Физиология растений и клеток . 55 (11): 1864–1872. : 10.1093 /pcp/pcu129 . doi «Бифункциональный растительный рецептор, OsCERK1 .
- ^ Мията, Кана; Акамацу, Акира; Цзян, Венди; Такаока, Рё; Абэ, Сацуки; Каку, Ханаэ; « OsSYMRK играет важную роль в симбиозе AM у риса (Oryza sativa)» . Физиология растений и клеток . 64 ): 378–391 doi : pcp/ pcad006 ISSN 0032-0781 . / 10.1093 ( 4
- ^ Венкатешваран, Мутусубраманиан; Джаяраман, Дхилипкумар; Шабо, Мирей; Жанр, Андреа; Баллон, Эллисон Дж.; Маэда, Джунко; Форши, Кэрри; Оса — Дезире; Квечен, Николас В.; Кун, Джошуа Дж.; Баркер, Дэвид Г.; Анн, Жан-Мишель (4 августа 2015 г.). «Роль мевалонатного пути в ранней симбиотической передаче сигналов растений» . Труды Национальной академии наук . Откр. 112 (31): 9781–9786. Бибкод : 2015PNAS..112.9781V . дои : 10.1073/pnas.1413762112 . ПМЦ 4534228 . ПМИД 26199419 .
- ^ Шарпантье, Мириам; Олдройд, Джайлз ЭД (8 октября 2013 г.). «Передача сигналов ядерного кальция в растениях» . Физиология растений . 163 (2): 496–503. дои : 10.1104/стр.113.220863 . ПМЦ 3793031 . ПМИД 23749852 .
- ^ Jump up to: а б Саид, Ваджиха; Насим, Саадия; Али, Захид (28 августа 2017 г.). «Биосинтез стриголактонов и их роль в устойчивости растений к абиотическому стрессу: критический обзор» . Границы в науке о растениях . 8 : 1487. doi : 10.3389/fpls.2017.01487 . ПМК 5581504 . ПМИД 28894457 .
- ^ Jump up to: а б Аль-Бабили, Салим; Бауместер, Харро Дж. (29 апреля 2015 г.). «Стриголактоны, новый растительный гормон каротиноидного происхождения». Ежегодный обзор биологии растений . 66 (1): 161–186. doi : 10.1146/annurev-arplant-043014-114759 . ПМИД 25621512 .
- ^ Гутжар, Кэролайн; Гоббато, Энрико; Чхве, Чонмин; Риман, Майкл; Джонстон, Мэтью Г.; Саммерс, Уильям; Карбоннел, Сэми; Мэнсфилд, Кэтрин; Ян, Шу-И; Надаль, ВМФ; Акоста, Иван; Такано, Макото; Цзяо, Вэнь-Бяо; Шнебергер, Корбиниан; Келли, Кристина А. (18 декабря 2015 г.). «Восприятие рисом симбиотических арбускулярных микоризных грибов требует рецепторного комплекса каррикина» . Наука 350 (6267): 1521–1524. дои : 10.1126/science.aac9715 . ISSN 0036-8075 . ПМИД 26680197 . S2CID 206641200 .
- ^ Надаль, Марина; Сойерс, Руайрид; Насим, Шамун; Бассин, Барбара; Кулицке, Коринна; Шарман, Эбигейл; Ан, Гюнхын; Ан, Кёнсук; Ахерн, Кевин Р.; Ромаг, Аманда; Брутнелл, Томас П.; Гутжар, Кэролайн; Гельднер, Нико; Ру, Кристоф; Мартинойя, Энрико (26 мая 2017 г.). «Переносчик N-ацетилглюкозамина, необходимый для арбускулярного микоризного симбиоза риса и кукурузы» . Природные растения . 3 (6): 17073. doi : 10.1038/nplants.2017.73 . ISSN 2055-0278 . ПМЦ 5685555 . ПМИД 28548655 .
- ^ Jump up to: а б Накагава, Томоми; Имаидзуми-Анраку, Харуко (декабрь 2015 г.). «Арбускулярная микориза риса как инструмент изучения молекулярных механизмов грибкового симбиоза и потенциальная мишень для повышения продуктивности» . Рис . 8 (1): 32. дои : 10.1186/s12284-015-0067-0 . ПМЦ 4626465 . ПМИД 26516078 .
- ^ Jump up to: а б Расмуссен, СР; Фюхтбауэр, В.; Новеро, М.; Вольпе, В.; Малков Н.; Жанр, А.; Бонфанте, П.; Стугард, Дж.; Радутойу, С. (20 июля 2016 г.). «Внутрирадикальная колонизация арбускулярными микоризными грибами вызывает индукцию липохитоолигосахаридного рецептора» . Научные отчеты . 6 (1): 29733. Бибкод : 2016NatSR...629733R . дои : 10.1038/srep29733 . ПМЦ 4951684 . ПМИД 27435342 .
- ^ Жанр, А.; Бонфанте, П. (февраль 2020 г.). «Рисовый рецептор сигналов микоризных грибов открывает новые возможности для развития устойчивых методов ведения сельского хозяйства» . Молекулярный завод . 13 (2): 181–183. дои : 10.1016/j.molp.2020.01.009 . hdl : 2318/1726475 . ПМИД 31981734 . S2CID 210912425 .
- ^ Мията, Кана; Козаки, Тошинори; Козай, Юсуке; Одзава, Кенджиро; Исии, Кадзуо; Асамидзу, Эрика; Окабе, Ёсихиро; Умехара, Ёске; Миямото, Аяно; Кобаэ, Ёсихиро; Акияма, Коки; Каку, Ханаэ; Нисидзава, Йоко; Сибуя, Наото; Накагава, Томоми (ноябрь 2014 г.). «Бифункциональный растительный рецептор, OsCERK1, регулирует как хитин-запускаемый иммунитет, так и арбускулярный микоризный симбиоз у риса» . Физиология растений и клеток . 55 (11): 1864–1872. дои : 10.1093/pcp/pcu129 . ISSN 1471-9053 . ПМИД 25231970 .
- ^ Jump up to: а б с д Буэндиа, Луис; Жирарден, Ариана; Ван, Тунмин; Коттрет, Людовик; Лефевр, Бенуа (24 октября 2018 г.). «Семейства рецептороподобных киназ LysM и рецептороподобных белков LysM: обновленная информация о филогении и функциональной характеристике» . Границы в науке о растениях . 9 : 1531. doi : 10.3389/fpls.2018.01531 . ISSN 1664-462X . ПМК 6207691 . ПМИД 30405668 .
- ^ Хэ, Цзян, Чи; Лю, Хуань, Ян, Чэнь, Си; Ван, Дапэн; Ли, Вэйчао; Ю, Цзухуа (декабрь 2019 г.). рецептора LysM опосредует восприятие арбускулярного микоризного симбиотического сигнала в рисе» 12 . Гетеромер ( ): 1561–1576. 12 « Гуойонг ; 2019.10.015 . ПМИД 31706032. S2CID 207966375 .
- ^ Jump up to: а б с д Накагава, Томоми; Имаидзуми-Анраку, Харуко (декабрь 2015 г.). «Арбускулярная микориза риса как инструмент изучения молекулярных механизмов грибкового симбиоза и потенциальная мишень для повышения продуктивности» . Рис . 8 (1): 32. дои : 10.1186/s12284-015-0067-0 . ISSN 1939-8425 . ПМЦ 4626465 . ПМИД 26516078 .
- ^ Пашковский, Ута (октябрь 2006 г.). «Путешествие по передаче сигналов в арбускулярных микоризных симбиозах, 2006 г.» . Новый фитолог . 172 (1): 35–46. дои : 10.1111/j.1469-8137.2006.01840.x . ISSN 0028-646X . ПМИД 16945087 .
- ^ Косута, Соня; Шабо, Мирей; Луньон, Жеральдин; Гоф, Клэр; Денари, Жан; Баркер, Дэвид Г.; Бекар, Гийом (01 марта 2003 г.). «Диффузионный фактор арбускулярных микоризных грибов индуцирует симбиоз-специфическую экспрессию MtENOD11 в корнях Medicago truncatula» . Физиология растений . 131 (3): 952–962. дои : 10.1104/стр.011882 . ISSN 1532-2548 . ПМК 166861 . ПМИД 12644648 .
- ^ Маклин, Эллисон М.; Браво, Армандо; Харрисон, Мария Дж. (октябрь 2017 г.). «Сигнальные сигналы растений и метаболические пути, обеспечивающие арбускулярный микоризный симбиоз» . Растительная клетка . 29 (10): 2319–2335. дои : 10.1105/tpc.17.00555 . ISSN 1040-4651 . ПМК 5940448 . ПМИД 28855333 .
- ^ Жанр, А.; Бонфанте, П. (февраль 2020 г.). «Рисовый рецептор сигналов микоризных грибов открывает новые возможности для развития устойчивых методов ведения сельского хозяйства» . Молекулярный завод . 13 (2): 181–183. дои : 10.1016/j.molp.2020.01.009 . hdl : 2318/1726475 . ПМИД 31981734 . S2CID 210912425 .
- ^ Мията, Кана; Козаки, Тошинори; Козай, Юсуке; Одзава, Кенджиро; Исии, Кадзуо; Асамидзу, Эрика; Окабе, Ёсихиро; Умехара, Ёске; Миямото, Аяно; Кобаэ, Ёсихиро; Акияма, Коки; Каку, Ханаэ; Нисидзава, Йоко; Сибуя, Наото; Накагава, Томоми (ноябрь 2014 г.). «Бифункциональный растительный рецептор, OsCERK1, регулирует как хитин-запускаемый иммунитет, так и арбускулярный микоризный симбиоз у риса» . Физиология растений и клеток . 55 (11): 1864–1872. дои : 10.1093/pcp/pcu129 . ISSN 1471-9053 . ПМИД 25231970 .
- ^ 3. Маклин А.М., Браво А., Харрисон М.Дж. Сигнальные и метаболические пути растений, обеспечивающие арбускулярномикоризный симбиоз. Растительная клетка. 2017 год; 29(10):2319-2335. https://doi.org/10.1105/tpc.17.00555
- ^ Jump up to: а б с д и Бонфанте, Паола; Жанр, Андреа (27 июля 2010 г.). «Механизмы, лежащие в основе полезных взаимодействий растений и грибов в микоризном симбиозе» . Природные коммуникации . 1 (1): 48. Бибкод : 2010NatCo...1...48B . дои : 10.1038/ncomms1046 . hdl : 2318/77771 . ISSN 2041-1723 . ПМИД 20975705 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Маклин, Эллисон М.; Браво, Армандо; Харрисон, Мария Дж. (октябрь 2017 г.). «Сигнальные сигналы растений и метаболические пути, обеспечивающие арбускулярный микоризный симбиоз» . Растительная клетка . 29 (10): 2319–2335. дои : 10.1105/tpc.17.00555 . ISSN 1040-4651 . ПМК 5940448 . ПМИД 28855333 .
- ^ Jump up to: а б Венкатешваран, Мутусубраманиан; Джаяраман, Дхилипкумар; Шабо, Мирей; Жанр, Андреа; Баллон, Эллисон Дж.; Маэда, Джунко; Форши, Кэрри; Оса — Дезире; Квечен, Николас В.; Кун, Джошуа Дж.; Баркер, Дэвид Г.; Год, Жан-Мишель (04 августа 2015 г.). «Роль мевалонатного пути в ранней симбиотической передаче сигналов растений» . Труды Национальной академии наук . Откр. 112 (31): 9781–9786. Бибкод : 2015PNAS..112.9781V . дои : 10.1073/pnas.1413762112 . ISSN 0027-8424 . ПМЦ 4534228 . ПМИД 26199419 .
- ^ Косута, Соня; Шабо, Мирей; Луньон, Жеральдин; Гоф, Клэр; Денари, Жан; Баркер, Дэвид Г.; Бекар, Гийом (01 марта 2003 г.). «Диффузионный фактор арбускулярных микоризных грибов индуцирует симбиоз-специфическую экспрессию MtENOD11 в корнях Medicago truncatula» . Физиология растений . 131 (3): 952–962. дои : 10.1104/стр.011882 . ISSN 1532-2548 . ПМК 166861 . ПМИД 12644648 .
- ^ Пимприкар, Прия; Гутяхр, Кэролайн (01 апреля 2018 г.). «Транкрипционная регуляция развития арбускулярной микоризы» . Физиология растений и клеток . 59 (4): 678–695. дои : 10.1093/pcp/pcy024 . ISSN 0032-0781 . ПМИД 29425360 .
- ^ Хирш, Сибилла; Олдройд, Джайлз ЭД (август 2009 г.). «Факторы транскрипции GRAS-домена, регулирующие развитие растений» . Сигнализация и поведение растений . 4 (8): 698–700. дои : 10.4161/psb.4.8.9176 . ISSN 1559-2324 . ПМК 2801379 . ПМИД 19820314 .
- ^ Хо-Плагаро, Таня; Гарсиа-Гарридо, Хосе Мануэль (2022). «Многообразные и интерактивные роли факторов транскрипции GRAS во время развития арбускулярной микоризы» . Границы в науке о растениях . 13 . дои : 10.3389/fpls.2022.836213 . ISSN 1664-462X . ПМЦ 8996055 . ПМИД 35419017 .
- ^ Моханта, Тапан Кумар; Бэ, Ханхун (январь 2015 г.). «Функциональная геномика и сигнальные события в микоризном симбиозе» . Журнал взаимодействия растений . 10 (1): 21–40. дои : 10.1080/17429145.2015.1005180 . ISSN 1742-9145 . S2CID 85269506 .
- ^ Пашковский, Ута (октябрь 2006 г.). «Путешествие по передаче сигналов в арбускулярных микоризных симбиозах, 2006 г.» . Новый фитолог . 172 (1): 35–46. дои : 10.1111/j.1469-8137.2006.01840.x . ISSN 0028-646X . ПМИД 16945087 .
- ^ Jump up to: а б Лугинбюль, Леони Х.; Менар, Уильям Н.; Куруп, Смит; Ван Эрп, Гарри; Радхакришнан, Гуру В.; Брейкспир, Эндрю; Олдройд, Джайлз ЭД; Истмонд, Питер Дж. (16 июня 2017 г.). «Жирные кислоты, синтезируемые растением-хозяином в арбускулярных микоризных грибах» . Наука 356 (6343): 1175–1178. дои : 10.1126/science.aan0081 . ISSN 0036-8075 . ПМИД 28596311 . S2CID 206658467 .
- ^ Браво, Армандо; Брэндс, Матиас; Вевер, Вера; Дорманн, Питер; Харрисон, Мария Дж. (июнь 2017 г.). «Специфические для арбускулярной микоризы ферменты FatM и RAM 2 тонко настраивают биосинтез липидов, способствуя развитию арбускулярной микоризы» . Новый фитолог . 214 (4): 1631–1645. дои : 10.1111/nph.14533 . ISSN 0028-646X . ПМИД 28380681 . S2CID 3481181 .
- ^ Го, Руй; Ву, Я-Нан; Лю, Ченг-Чен; Лю, Ин-На; Тянь, Ли; Ченг, Цзянь-Фей; Пан, Чжиюн; Ван, Донг; Ван, Бин (2022). «OsADK1, новая киназа, регулирующая арбускулярный микоризный симбиоз риса». Новый фитолог . 234 (1): 256–268. дои : 10.1111/nph.17979 . ISSN 0028-646X . ПМИД 35133010 .
- ^ Ленг, Сяотун; Ван, Лунсян, Цзоу, Цзычао; Чжао, Нинкан; Дуанму, Децян; Марсель; , Хайтао ; Бухер : 10.1093 /plcell/ 10226568. Получено 30 koad050 номер PMC октября 2023 г.
- ^ Рот, Ронель; Кьяпелло, Марко; Монтеро, Эктор; Гериг, Питер; Гроссманн, Йонас; О'Холлеран, Кевин; Харткен, Дениз; Уолтерс, Фергюс; Ян, Шу-И; Хиллмер, Стефан; Шумахер, Карин; Боуден, Сара; Безумие, Мелани; Уоллингтон, Эмма Дж.; Мияо, Акио (08.11.2018). «Киназа, подобная серин/треониновому рецептору риса, регулирует арбускулярный микоризный симбиоз на периарбускулярной мембране» . Природные коммуникации . 9 (1): 4677. doi : 10.1038/s41467-018-06865-z . ISSN 2041-1723 . ПМК 6224560 . ПМИД 30410018 .
- ^ Монтеро, Эктор; Ли, Так; Пакер, Боас; Феррерас-Гарручо, Габриэль; Олдройд, Джайлз; Брокингтон, Сэмюэл Ф.; Мияо, Акио; Пашковский, Ута (22 июня 2021 г.). «Связанная с микоризой рецептороподобная киназа древнего происхождения из зеленой линии» . Труды Национальной академии наук . 118 (25). дои : 10.1073/pnas.2105281118 . ISSN 0027-8424 . ПМЦ 8237591 . ПМИД 34161289 .
- ^ Ши , Цзиньцай , Ваньсяо : 10.1016 j.molp.2017.07.012 / . PMID 28782719 Ван
- ^ Бармен Ягнасен; Саманта, Авик; Саха, Бабита; Датта, Сирадж (декабрь 2016 г.). «Микориза: древнейшая ассоциация растений и грибов» . Резонанс . 21 (12): 1093–1104. дои : 10.1007/ s12045-016-0421-6 ISSN 0971-8044 . S2CID 109932925 .
- ^ Эрколин, Флавия; Рейнхардт, Дидье (июль 2011 г.). «Успешные совместные предприятия растений: арбускулярная микориза и не только» (PDF) . Тенденции в науке о растениях . 16 (7): 356–362. doi : 10.1016/j.tplants.2011.03.006 . ПМИД 21459657 .
- ^ Де-ла-Пенья, Клелия; Бадри, Даякар В.; Лойола-Варгас, Виктор М. (2012), Виванко, Хорхе М.; Балушка, Франтишек (ред.), «Выделения корней растений и их взаимодействие с соседями» , Секреции и экссудаты в биологических системах , Передача сигналов и связь в растениях, том. 12, Берлин, Гейдельберг: Springer Berlin Heidelberg, стр. 1–26, doi : 10.1007/978-3-642-23047-9_1 , ISBN 978-3-642-23046-2 , получено 24 октября 2023 г.