Транспортер ABC
| ABC Транспортер, НБД | |||
|---|---|---|---|
 Транспортер витамина В12 , BtuCD PDB 1l7v | |||
| Идентификаторы | |||
| Символ | ABC_tran | ||
| Пфам | PF00005 | ||
| ИнтерПро | ИПР003439 | ||
| PROSITE | PDOC00185 | ||
| СКОП2 | 1b0u / SCOPe / СУПФАМ | ||
| TCDB | 3.A.1 | ||
| OPM superfamily | 17 | ||
| OPM protein | 3g5u | ||
| |||
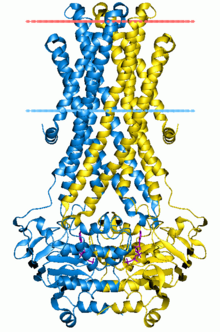
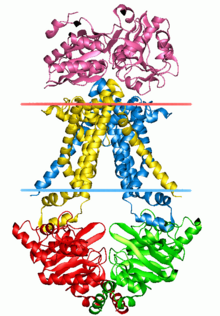
Транспортеры ABC , кассетные транспортеры, связывающие АТФ-синтазу (АТФ), представляют собой суперсемейство транспортных систем, которое является одним из крупнейших и, возможно, одним из старейших семейств генов . Он представлен во всех современных типах , от прокариотов до человека . [1] [2] [3] Транспортеры ABC относятся к транслоказам .
Транспортеры ABC часто состоят из множества субъединиц, одна или две из которых представляют собой трансмембранные белки , а одна или две из них представляют собой мембраносвязанные AAA- АТФазы . [ нужна ссылка ] Субъединицы АТФазы используют энергию связывания и гидролиза аденозинтрифосфата (АТФ) для обеспечения энергии, необходимой для перемещения субстратов через мембраны либо для поглощения, либо для экспорта субстрата.
Большинство систем поглощения также имеют экстрацитоплазматический рецептор, белок, связывающий растворенные вещества. Некоторые гомологичные АТФазы участвуют в процессах, не связанных с транспортом, таких как трансляция РНК и репарация ДНК . [4] [5] Транспортеры ABC считаются суперсемейством ABC на основании сходства последовательности и организации их доменов АТФ-связывающей кассеты (ABC), хотя интегральные мембранные белки, по-видимому, несколько раз эволюционировали независимо и, таким образом, включают разные семейства белков. [6] Like the ABC exporters, it is possible that the integral membrane proteins of ABC uptake systems also evolved at least three times independently, based on their high resolution three-dimensional structures.[7] Портеры поглощения ABC поглощают большое количество питательных веществ, предшественников биосинтеза, микроэлементов и витаминов , а экспортеры транспортируют липиды , стерины , лекарства и большое количество первичных и вторичных метаболитов. Некоторые из этих экспортеров в организме человека участвуют в резистентности опухолей, муковисцидозе и ряде других наследственных заболеваний человека. Высокий уровень экспрессии генов, кодирующих некоторые из этих экспортеров, как у прокариотических, так и у эукариотических организмов (включая человека), приводит к развитию устойчивости к множеству лекарств, таких как антибиотики и противораковые средства.
Hundreds of ABC transporters have been characterized from both prokaryotes and eukaryotes.[8] ABC genes are essential for many processes in the cell, and mutations in human genes cause or contribute to several human genetic diseases.[9] Forty eight ABC genes have been reported in humans. Among these, many have been characterized and shown to be causally related to diseases present in humans such as cystic fibrosis, adrenoleukodystrophy, Stargardt disease, drug-resistant tumors, Dubin–Johnson syndrome, Byler's disease, progressive familiar intrahepatic cholestasis, X-linked sideroblastic anemia, ataxia, and persistent and hyperinsulimenic hypoglycemia.[8] ABC transporters are also involved in multiple drug resistance, and this is how some of them were first identified. When the ABC transport proteins are overexpressed in cancer cells, they can export anticancer drugs and render tumors resistant.[10]
Function
[edit]ABC transporters utilize the energy of ATP binding and hydrolysis to transport various substrates across cellular membranes. They are divided into three main functional categories. In prokaryotes, importers mediate the uptake of nutrients into the cell. The substrates that can be transported include ions, amino acids, peptides, sugars, and other molecules that are mostly hydrophilic. The membrane-spanning region of the ABC transporter protects hydrophilic substrates from the lipids of the membrane bilayer thus providing a pathway across the cell membrane. Eukaryotes do not possess any importers. Exporters or effluxers, which are present both in prokaryotes and eukaryotes, function as pumps that extrude toxins and drugs out of the cell. In gram-negative bacteria, exporters transport lipids and some polysaccharides from the cytoplasm to the periplasm. The third subgroup of ABC proteins do not function as transporters, but are rather involved in translation and DNA repair processes.[4]
Prokaryotic
[edit]Bacterial ABC transporters are essential in cell viability, virulence, and pathogenicity.[1][4] Iron ABC uptake systems, for example, are important effectors of virulence.[11] Pathogens use siderophores, such as Enterobactin, to scavenge iron that is in complex with high-affinity iron-binding proteins or erythrocytes. These are high-affinity iron-chelating molecules that are secreted by bacteria and reabsorb iron into iron-siderophore complexes. The chvE-gguAB gene in Agrobacterium tumefaciens encodes glucose and galactose importers that are also associated with virulence.[12][13] Transporters are extremely vital in cell survival such that they function as protein systems that counteract any undesirable change occurring in the cell. For instance, a potential lethal increase in osmotic strength is counterbalanced by activation of osmosensing ABC transporters that mediate uptake of solutes.[14] Other than functioning in transport, some bacterial ABC proteins are also involved in the regulation of several physiological processes.[4]
In bacterial efflux systems, certain substances that need to be extruded from the cell include surface components of the bacterial cell (e.g. capsular polysaccharides, lipopolysaccharides, and teichoic acid), proteins involved in bacterial pathogenesis (e.g. hemolysis, heme-binding protein, and alkaline protease), heme, hydrolytic enzymes, S-layer proteins, competence factors, toxins, antibiotics, bacteriocins, peptide antibiotics, drugs and siderophores.[15] They also play important roles in biosynthetic pathways, including extracellular polysaccharide biosynthesis[16] and cytochrome biogenesis.[17]
Eukaryotic
[edit]Although most eukaryotic ABC transporters are effluxers, some are not directly involved in transporting substrates. In the cystic fibrosis transmembrane regulator (CFTR) and in the sulfonylurea receptor (SUR), ATP hydrolysis is associated with the regulation of opening and closing of ion channels carried by the ABC protein itself or other proteins.[5]
Human ABC transporters are involved in several diseases that arise from polymorphisms in ABC genes and rarely due to complete loss of function of single ABC proteins.[18] Such diseases include Mendelian diseases and complex genetic disorders such as cystic fibrosis, adrenoleukodystrophy, Stargardt disease, Tangier disease, immune deficiencies, progressive familial intrahepatic cholestasis, Dubin–Johnson syndrome, Pseudoxanthoma elasticum, persistent hyperinsulinemic hypoglycemia of infancy due to focal adenomatous hyperplasia, X-linked sideroblastosis and anemia, age-related macular degeneration, familial hypoapoproteinemia, Retinitis pigmentosum, cone rod dystrophy, and others.[5] The human ABCB (MDR/TAP) family is responsible for multiple drug resistance (MDR) against a variety of structurally unrelated drugs. ABCB1 or MDR1 P-glycoprotein is also involved in other biological processes for which lipid transport is the main function. It is found to mediate the secretion of the steroid aldosterone by the adrenals, and its inhibition blocked the migration of dendritic immune cells,[19] possibly related to the outward transport of the lipid platelet activating factor (PAF). It has also been reported that ABCB1 mediates transport of cortisol and dexamethasone, but not of progesterone in ABCB1 transfected cells. MDR1 can also transport cholesterol, short-chain and long-chain analogs of phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidylserine (PS), sphingomyelin (SM), and glucosylceramide (GlcCer). Multispecific transport of diverse endogenous lipids through the MDR1 transporter can possibly affect the transbilayer distribution of lipids, in particular of species normally predominant on the inner plasma membrane leaflet such as PS and PE.[18]
More recently, ABC-transporters have been shown to exist within the placenta, indicating they could play a protective role for the developing fetus against xenobiotics.[20] Evidence has shown that placental expression of the ABC-transporters P-glycoprotein (P-gp) and breast cancer resistance protein (BCRP) are increased in preterm compared to term placentae, with P-gp expression further increased in preterm pregnancies with chorioamnionitis.[21] To a lesser extent, increasing maternal BMI also associated with increased placental ABC-transporter expression, but only at preterm.[21]
Structure
[edit]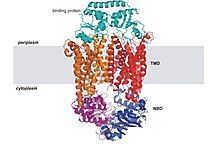
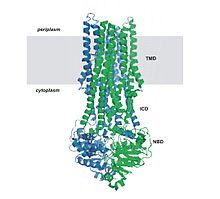
All ABC transport proteins share a structural organization consisting of four core domains.[22] These domains consist of two trans-membrane (T) domains and two cytosolic (A) domains. The two T domains alternate between an inward and outward facing orientation, and the alternation is powered by the hydrolysis of adenosine triphosphate or ATP. ATP binds to the A subunits and it is then hydrolyzed to power the alternation, but the exact process by which this happens is not known. The four domains can be present in four separate polypeptides, which occur mostly in bacteria, or present in one or two multi-domain polypeptides.[10] When the polypeptides are one domain, they can be referred to as a full domain, and when they are two multi-domains they can be referred to as a half domain.[9] The T domains are each built of typically 10 membrane spanning alpha helices, through which the transported substance can cross through the plasma membrane. Also, the structure of the T domains determines the specificity of each ABC protein. In the inward facing conformation, the binding site on the A domain is open directly to the surrounding aqueous solutions. This allows hydrophilic molecules to enter the binding site directly from the inner leaflet of the phospholipid bilayer. In addition, a gap in the protein is accessible directly from the hydrophobic core of the inner leaflet of the membrane bilayer. This allows hydrophobic molecules to enter the binding site directly from the inner leaflet of the phospholipid bilayer. After the ATP powered move to the outward facing conformation, molecules are released from the binding site and allowed to escape into the exoplasmic leaflet or directly into the extracellular medium.[10]
The common feature of all ABC transporters is that they consist of two distinct domains, the transmembrane domain (TMD) and the nucleotide-binding domain (NBD). The TMD, also known as membrane-spanning domain (MSD) or integral membrane (IM) domain, consists of alpha helices, embedded in the membrane bilayer. It recognizes a variety of substrates and undergoes conformational changes to transport the substrate across the membrane. The sequence and architecture of TMDs is variable, reflecting the chemical diversity of substrates that can be translocated. The NBD or ATP-binding cassette (ABC) domain, on the other hand, is located in the cytoplasm and has a highly conserved sequence. The NBD is the site for ATP binding.[23] In most exporters, the N-terminal transmembrane domain and the C-terminal ABC domains are fused as a single polypeptide chain, arranged as TMD-NBD-TMD-NBD. An example is the E. coli hemolysin exporter HlyB. Importers have an inverted organization, that is, NBD-TMD-NBD-TMD, where the ABC domain is N-terminal whereas the TMD is C-terminal, such as in the E. coli MacB protein responsible for macrolide resistance.[4][5]
The structural architecture of ABC transporters consists minimally of two TMDs and two NBDs. Four individual polypeptide chains including two TMD and two NBD subunits, may combine to form a full transporter such as in the E. coli BtuCD[24][25] importer involved in the uptake of vitamin B12. Most exporters, such as in the multidrug exporter Sav1866[26] from Staphylococcus aureus, are made up of a homodimer consisting of two half transporters or monomers of a TMD fused to a nucleotide-binding domain (NBD). A full transporter is often required to gain functionality. Some ABC transporters have additional elements that contribute to the regulatory function of this class of proteins. In particular, importers have a high-affinity binding protein (BP) that specifically associates with the substrate in the periplasm for delivery to the appropriate ABC transporter. Exporters do not have the binding protein but have an intracellular domain (ICD) that joins the membrane-spanning helices and the ABC domain. The ICD is believed to be responsible for communication between the TMD and NBD.[23]
Transmembrane domain (TMD)
[edit]Most transporters have transmembrane domains that consist of a total of 12 α-helices with 6 α-helices per monomer. Since TMDs are structurally diverse, some transporters have varying number of helices (between six and eleven). The TM domains are categorized into three distinct sets of folds: type I ABC importer, type II ABC importer and ABC exporter folds. The classification of importer folds is based on detailed characterization of the sequences.[23]
The type I ABC importer fold was originally observed in the ModB TM subunit of the molybdate transporter.[27] This diagnostic fold can also be found in the MalF and MalG TM subunits of MalFGK2[28] and the Met transporter MetI.[29] In the MetI transporter, a minimal set of 5 transmembrane helices constitute this fold while an additional helix is present for both ModB and MalG. The common organization of the fold is the "up-down" topology of the TM2-5 helices that lines the translocation pathway and the TM1 helix wrapped around the outer, membrane-facing surface and contacts the other TM helices.
The type II ABC importer fold is observed in the twenty TM helix-domain of BtuCD[24] and in Hi1471,[30] a homologous transporter from Haemophilus influenzae. In BtuCD, the packing of the helices is complex. The noticeable pattern is that the TM2 helix is positioned through the center of the subunit where it is surrounded in close proximity by the other helices. Meanwhile, the TM5 and TM10 helices are positioned in the TMD interface. The membrane spanning region of ABC exporters is organized into two "wings" that are composed of helices TM1 and TM2 from one subunit and TM3-6 of the other, in a domain-swapped arrangement. A prominent pattern is that helices TM1-3 are related to TM4-6 by an approximate twofold rotation around an axis in the plane of the membrane.[23]
The exporter fold is originally observed in the Sav1866 structure. It contains 12 TM helices, 6 per monomer.[23]
Nucleotide-binding domain (NBD)
[edit]
The ABC domain consists of two domains, the catalytic core domain similar to RecA-like motor ATPases and a smaller, structurally diverse α-helical subdomain that is unique to ABC transporters. The larger domain typically consists of two β-sheets and six α helices, where the catalytic Walker A motif (GXXGXGKS/T where X is any amino acid) or P-loop and Walker B motif (ΦΦΦΦD, of which Φ is a hydrophobic residue) is situated. The helical domain consists of three or four helices and the ABC signature motif, also known as LSGGQ motif, linker peptide or C motif. The ABC domain also has a glutamine residue residing in a flexible loop called Q loop, lid or γ-phosphate switch, that connects the TMD and ABC. The Q loop is presumed to be involved in the interaction of the NBD and TMD, particularly in the coupling of nucleotide hydrolysis to the conformational changes of the TMD during substrate translocation. The H motif or switch region contains a highly conserved histidine residue that is also important in the interaction of the ABC domain with ATP. The name ATP-binding cassette is derived from the diagnostic arrangement of the folds or motifs of this class of proteins upon formation of the ATP sandwich and ATP hydrolysis.[4][15][23]
ATP binding and hydrolysis
[edit]Dimer formation of the two ABC domains of transporters requires ATP binding.[31] It is generally observed that the ATP bound state is associated with the most extensive interface between ABC domains, whereas the structures of nucleotide-free transporters exhibit conformations with greater separations between the ABC domains.[23] Structures of the ATP-bound state of isolated NBDs have been reported for importers including HisP,[32] GlcV,[33] MJ1267,[34] E. coli MalK (E.c.MalK),[35] T. litoralis MalK (TlMalK),[36] and exporters such as TAP,[37] HlyB,[38] MJ0796,[39][40] Sav1866,[26] and MsbA.[41] In these transporters, ATP is bound to the ABC domain. Two molecules of ATP are positioned at the interface of the dimer, sandwiched between the Walker A motif of one subunit and the LSGGQ motif of the other.[23] This was first observed in Rad50[42] and reported in structures of MJ0796, the NBD subunit of the LolD transporter from Methanococcus jannaschii[40] and E.c.MalK of a maltose transporter.[35] These structures were also consistent with results from biochemical studies revealing that ATP is in close contact with residues in the P-loop and LSGGQ motif during catalysis.[43]
Nucleotide binding is required to ensure the electrostatic and/or structural integrity of the active site and contribute to the formation of an active NBD dimer.[44] Binding of ATP is stabilized by the following interactions: (1) ring-stacking interaction of a conserved aromatic residue preceding the Walker A motif and the adenosine ring of ATP,[45][46] (2) hydrogen-bonds between a conserved lysine residue in the Walker A motif and the oxygen atoms of the β- and γ-phosphates of ATP and coordination of these phosphates and some residues in the Walker A motif with Mg2+ ion,[33][37] and (3) γ-phosphate coordination with side chain of serine and backbone amide groups of glycine residues in the LSGGQ motif.[47] In addition, a residue that suggests the tight coupling of ATP binding and dimerization, is the conserved histidine in the H-loop. This histidine contacts residues across the dimer interface in the Walker A motif and the D loop, a conserved sequence following the Walker B motif.[35][40][42][48]
The enzymatic hydrolysis of ATP requires proper binding of the phosphates and positioning of the γ-phosphate to the attacking water.[23] In the nucleotide binding site, the oxygen atoms of the β- and γ-phosphates of ATP are stabilized by residues in the Walker A motif[49][50] and coordinate with Mg2+.[23] This Mg2+ ion also coordinates with the terminal aspartate residue in the Walker B motif through the attacking H2O.[33][34][39] A general base, which may be the glutamate residue adjacent to the Walker B motif,[31][40][46] glutamine in the Q-loop,[30][36][40] or a histidine in the switch region that forms a hydrogen bond with the γ-phosphate of ATP, is found to catalyze the rate of ATP hydrolysis by promoting the attacking H2O.[35][36][40][48] The precise molecular mechanism of ATP hydrolysis is still controversial.[4]
Mechanism of transport
[edit]ABC transporters are active transporters, that is, they use energy in the form of adenosine triphosphate (ATP) to translocate substrates across cell membranes. These proteins harness the energy of ATP binding and/or hydrolysis to drive conformational changes in the transmembrane domain (TMD) and consequently transport molecules.[51] ABC importers and exporters have a common mechanism for transporting substrates. They are similar in their structures. The model that describes the conformational changes associated with the binding of the substrate is the alternating-access model. In this model, the substrate binding site alternates between outward- and inward-facing conformations. The relative binding affinities of the two conformations for the substrate largely determines the net direction of transport. For importers, since translocation is directed from the periplasm to the cytoplasm, the outward-facing conformation has higher binding affinity for the substrate. In contrast, the substrate binding affinity in exporters is greater in the inward-facing conformation.[23] A model that describes the conformational changes in the nucleotide-binding domain (NBD) as a result of ATP binding and hydrolysis is the ATP-switch model. This model presents two principal conformations of the NBDs: formation of a closed dimer upon binding two ATP molecules and dissociation to an open dimer facilitated by ATP hydrolysis and release of inorganic phosphate (Pi) and adenosine diphosphate (ADP). Switching between the open and closed dimer conformations induces conformational changes in the TMD resulting in substrate translocation.[52]
The general mechanism for the transport cycle of ABC transporters has not been fully elucidated, but substantial structural and biochemical data has accumulated to support a model in which ATP binding and hydrolysis is coupled to conformational changes in the transporter. The resting state of all ABC transporters has the NBDs in an open dimer configuration, with low affinity for ATP. This open conformation possesses a chamber accessible to the interior of the transporter. The transport cycle is initiated by binding of substrate to the high-affinity site on the TMDs, which induces conformational changes in the NBDs and enhances the binding of ATP. Two molecules of ATP bind, cooperatively, to form the closed dimer configuration. The closed NBD dimer induces a conformational change in the TMDs such that the TMD opens, forming a chamber with an opening opposite to that of the initial state. The affinity of the substrate to the TMD is reduced, thereby releasing the substrate. Hydrolysis of ATP follows and then sequential release of Pi and then ADP restores the transporter to its basal configuration. Although a common mechanism has been suggested, the order of substrate binding, nucleotide binding and hydrolysis, and conformational changes, as well as interactions between the domains is still debated.[4][15][18][23][41][44][51][52][53][54][55]
Several groups studying ABC transporters have differing assumptions on the driving force of transporter function. It is generally assumed that ATP hydrolysis provides the principal energy input or "power stroke" for transport and that the NBDs operate alternately and are possibly involved in different steps in the transport cycle.[56] However, recent structural and biochemical data shows that ATP binding, rather than ATP hydrolysis, provides the "power stroke".[57] It may also be that since ATP binding triggers NBD dimerization, the formation of the dimer may represent the "power stroke." In addition, some transporters have NBDs that do not have similar abilities in binding and hydrolyzing ATP and that the interface of the NBD dimer consists of two ATP binding pockets suggests a concurrent function of the two NBDs in the transport cycle.[52]
Some evidence to show that ATP binding is indeed the power stroke of the transport cycle was reported.[52] It has been shown that ATP binding induces changes in the substrate-binding properties of the TMDs. The affinity of ABC transporters for substrates has been difficult to measure directly, and indirect measurements, for instance through stimulation of ATPase activity, often reflects other rate-limiting steps. Recently, direct measurement of vinblastine binding to permease-glycoprotein (P-glycoprotein) in the presence of nonhydrolyzable ATP analogs, e.g. 5'-adenylyl-β-γ-imidodiphosphate (AMP-PNP), showed that ATP binding, in the absence of hydrolysis, is sufficient to reduce substrate-binding affinity.[58] Also, ATP binding induces substantial conformational changes in the TMDs. Spectroscopic, protease accessibility and crosslinking studies have shown that ATP binding to the NBDs induces conformational changes in multidrug resistance-associated protein-1 (MRP1),[59] HisPMQ,[60] LmrA,[61] and Pgp.[62] Two dimensional crystal structures of AMP-PNP-bound Pgp showed that the major conformational change during the transport cycle occurs upon ATP binding and that subsequent ATP hydrolysis introduces more limited changes.[63] Rotation and tilting of transmembrane α-helices may both contribute to these conformational changes. Other studies have focused on confirming that ATP binding induces NBD closed dimer formation. Biochemical studies of intact transport complexes suggest that the conformational changes in the NBDs are relatively small. In the absence of ATP, the NBDs may be relatively flexible, but they do not involve a major reorientation of the NBDs with respect to the other domains. ATP binding induces a rigid body rotation of the two ABC subdomains with respect to each other, which allows the proper alignment of the nucleotide in the active site and interaction with the designated motifs. There is strong biochemical evidence that binding of two ATP molecules can be cooperative, that is, ATP must bind to the two active site pockets before the NBDs can dimerize and form the closed, catalytically active conformation.[52]
ABC importers
[edit]Most ABC transporters that mediate the uptake of nutrients and other molecules in bacteria rely on a high-affinity solute binding protein (BP). BPs are soluble proteins located in the periplasmic space between the inner and outer membranes of gram-negative bacteria. Gram-positive microorganisms lack a periplasm such that their binding protein is often a lipoprotein bound to the external face of the cell membrane. Some gram-positive bacteria have BPs fused to the transmembrane domain of the transporter itself.[4] The first successful x-ray crystal structure of an intact ABC importer is the molybdenum transporter (ModBC-A) from Archaeoglobus fulgidus.[27] Atomic-resolution structures of three other bacterial importers, E. coli BtuCD,[24] E. coli maltose transporter (MalFGK2-E),[28] and the putative metal-chelate transporter of Haemophilus influenzae, HI1470/1,[30] have also been determined. The structures provided detailed pictures of the interaction of the transmembrane and ABC domains as well as revealed two different conformations with an opening in two opposite directions. Another common feature of importers is that each NBD is bound to one TMD primarily through a short cytoplasmic helix of the TMD, the "coupling helix". This portion of the EAA loop docks in a surface cleft formed between the RecA-like and helical ABC subdomains and lies approximately parallel to the membrane bilayer.[54]
Large ABC importers
[edit]The BtuCD and HI1470/1 are classified as large (Type II) ABC importers. The transmembrane subunit of the vitamin B12 importer, BtuCD, contains 10 TM helices and the functional unit consists of two copies each of the nucleotide binding domain (NBD) and transmembrane domain (TMD). The TMD and NBD interact with one another via the cytoplasmic loop between two TM helices and the Q loop in the ABC. In the absence of nucleotide, the two ABC domains are folded and the dimer interface is open. A comparison of the structures with (BtuCDF) and without (BtuCD) binding protein reveals that BtuCD has an opening that faces the periplasm whereas in BtuCDF, the outward-facing conformation is closed to both sides of the membrane. The structures of BtuCD and the BtuCD homolog, HI1470/1, represent two different conformational states of an ABC transporter. The predicted translocation pathway in BtuCD is open to the periplasm and closed at the cytoplasmic side of the membrane while that of HI1470/1 faces the opposite direction and open only to the cytoplasm. The difference in the structures is a 9° twist of one TM subunit relative to the other.[4][23][54]
Small ABC importers
[edit]Structures of the ModBC-A and MalFGK2-E, which are in complex with their binding protein, correspond to small (Type I) ABC importers. The TMDs of ModBC-A and MalFGK2-E have only six helices per subunit. The homodimer of ModBC-A is in a conformation in which the TM subunits (ModB) orient in an inverted V-shape with a cavity accessible to the cytoplasm. The ABC subunits (ModC), on the other hand, are arranged in an open, nucleotide-free conformation, in which the P-loop of one subunit faces but is detached from the LSGGQ motif of the other. The binding protein ModA is in a closed conformation with substrate bound in a cleft between its two lobes and attached to the extracellular loops of ModB, wherein the substrate is sitting directly above the closed entrance of the transporter. The MalFGK2-E structure resembles the catalytic transition state for ATP hydrolysis. It is in a closed conformation where it contains two ATP molecules, sandwiched between the Walker A and B motifs of one subunit and the LSGGQ motif of the other subunit. The maltose binding protein (MBP or MalE) is docked on the periplasmic side of the TM subunits (MalF and MalG) and a large, occluded cavity can be found at the interface of MalF and MalG. The arrangement of the TM helices is in a conformation that is closed toward the cytoplasm but with an opening that faces outward. The structure suggests a possibility that MBP may stimulate the ATPase activity of the transporter upon binding.[4][23][54]
Mechanism of transport for importers
[edit]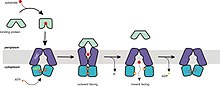
The mechanism of transport for importers supports the alternating-access model. The resting state of importers is inward-facing, where the nucleotide binding domain (NBD) dimer interface is held open by the TMDs and facing outward but occluded from the cytoplasm. Upon docking of the closed, substrate-loaded binding protein towards the periplasmic side of the transmembrane domains, ATP binds and the NBD dimer closes. This switches the resting state of transporter into an outward-facing conformation, in which the TMDs have reoriented to receive substrate from the binding protein. After hydrolysis of ATP, the NBD dimer opens and substrate is released into the cytoplasm. Release of ADP and Pi reverts the transporter into its resting state. The only inconsistency of this mechanism to the ATP-switch model is that the conformation in its resting, nucleotide-free state is different from the expected outward-facing conformation. Although that is the case, the key point is that the NBD does not dimerize unless ATP and binding protein is bound to the transporter.[4][15][23][52][54]
ABC exporters
[edit]Prokaryotic ABC exporters are abundant and have close homologues in eukaryotes. This class of transporters is studied based on the type of substrate that is transported. One class is involved in the protein (e.g. toxins, hydrolytic enzymes, S-layer proteins, lantibiotics, bacteriocins, and competence factors) export and the other in drug efflux. ABC transporters have gained extensive attention because they contribute to the resistance of cells to antibiotics and anticancer agents by pumping drugs out of the cells.[1][64][4] A common mechanism is the overexpression of ABC exporters like P-glycoprotein (P-gp/ABCB1), multidrug resistance-associated protein 1 (MRP1/ABCC1), and breast cancer resistance protein (BCRP/ABCG2) in cancer cells that limit the exposure to anticancer drugs.[65]
In gram-negative organisms, ABC transporters mediate secretion of protein substrates across inner and outer membranes simultaneously without passing through the periplasm. This type of secretion is referred to as type I secretion, which involves three components that function in concert: an ABC exporter, a membrane fusion protein (MFP), and an outer membrane factor (OMF). An example is the secretion of hemolysin (HlyA) from E. coli where the inner membrane ABC transporter HlyB interacts with an inner membrane fusion protein HlyD and an outer membrane facilitator TolC. TolC allows hemolysin to be transported across the two membranes, bypassing the periplasm.[1][64][15]
Bacterial drug resistance has become an increasingly major health problem. One of the mechanisms for drug resistance is associated with an increase in antibiotic efflux from the bacterial cell. Drug resistance associated with drug efflux, mediated by P-glycoprotein, was originally reported in mammalian cells. In bacteria, Levy and colleagues presented the first evidence that antibiotic resistance was caused by active efflux of a drug.[66] P-glycoprotein is the best-studied efflux pump and as such has offered important insights into the mechanism of bacterial pumps.[4] Although some exporters transport a specific type of substrate, most transporters extrude a diverse class of drugs with varying structure.[18] These transporters are commonly called multi-drug resistant (MDR) ABC transporters and sometimes referred to as "hydrophobic vacuum cleaners".[55]
Human ABCB1/MDR1 P-glycoprotein
[edit]P-glycoprotein (3.A.1.201.1) is a well-studied protein associated with multi-drug resistance. It belongs to the human ABCB (MDR/TAP) family and is also known as ABCB1 or MDR1 Pgp. MDR1 consists of a functional monomer with two transmembrane domains (TMD) and two nucleotide-binding domains (NBD). This protein can transport mainly cationic or electrically neutral substrates as well as a broad spectrum of amphiphilic substrates. The structure of the full-size ABCB1 monomer was obtained in the presence and absence of nucleotide using electron cryo crystallography. Without the nucleotide, the TMDs are approximately parallel and form a barrel surrounding a central pore, with the opening facing towards the extracellular side of the membrane and closed at the intracellular face. In the presence of the nonhydrolyzable ATP analog, AMP-PNP, the TMDs have a substantial reorganization with three clearly segregated domains. A central pore, which is enclosed between the TMDs, is slightly open towards the intracellular face with a gap between two domains allowing access of substrate from the lipid phase. Substantial repacking and possible rotation of the TM helices upon nucleotide binding suggests a helix rotation model for the transport mechanism.[18]
Plant transporters
[edit]The genome of the model plant Arabidopsis thaliana is capable of encoding 120 ABC proteins compared to 50-70 ABC proteins that are encoded by the human genome and fruit flies (Drosophila melanogaster). Plant ABC proteins are categorized in 13 subfamilies on the basis of size (full, half or quarter), orientation, and overall amino acid sequence similarity.[67] Multidrug resistant (MDR) homologs, also known as P-glycoproteins, represent the largest subfamily in plants with 22 members and the second largest overall ABC subfamily. The B subfamily of plant ABC transporters (ABCBs) are characterized by their localization to the plasma membrane.[68] Plant ABCB transporters are characterized by heterologously expressing them in Escherichia coli, Saccharomyces cerevisiae, Schizosaccharomyces pombe (fission yeast), and HeLa cells to determine substrate specificity. Plant ABCB transporters have shown to transport the phytohormone indole-3-acetic acid ( IAA),[69] also known as auxin, the essential regulator for plant growth and development.[70][71] The directional polar transport of auxin mediates plant environmental responses through processes such as phototropism and gravitropism.[72] Two of the best studied auxin transporters, ABCB1 and ABCB19, have been characterized to be primary auxin exporters[70] Other ABCB transporters such as ABCB4 participate in both the export and import of auxin[70] At low intracellular auxin concentrations ABCB4 imports auxin until it reaches a certain threshold which then reverses function to only export auxin.[70][73]
Sav1866
[edit]The first high-resolution structure reported for an ABC exporter was that of Sav1866 (3.A.1.106.2) from Staphylococcus aureus.[18][74] Sav1866 is a homolog of multidrug ABC transporters. It shows significant sequence similarity to human ABC transporters of subfamily B that includes MDR1 and TAP1/TAP2. The ATPase activity of Sav1866 is known to be stimulated by cancer drugs such as doxorubicin, vinblastine and others,[75] which suggests similar substrate specificity to P-glycoprotein and therefore a possible common mechanism of substrate translocation. Sav1866 is a homodimer of half transporters, and each subunit contains an N-terminal TMD with six helices and a C-terminal NBD. The NBDs are similar in structure to those of other ABC transporters, in which the two ATP binding sites are formed at the dimer interface between the Walker A motif of one NBD and the LSGGQ motif of the other. The ADP-bound structure of Sav1866 shows the NBDs in a closed dimer and the TM helices split into two "wings" oriented towards the periplasm, forming the outward-facing conformation. Each wing consists of helices TM1-2 from one subunit and TM3-6 from the other subunit. It contains long intracellular loops (ICLs or ICD) connecting the TMDs that extend beyond the lipid bilayer into the cytoplasm and interacts with the 8=D. Whereas the importers contain a short coupling helix that contact a single NBD, Sav1866 has two intracellular coupling helices, one (ICL1) contacting the NBDs of both subunits and the other (ICL2) interacting with only the opposite NBD subunit.[23][26][54]
MsbA
[edit]MsbA (3.A.1.106.1) is a multi-drug resistant (MDR) ABC transporter and possibly a lipid flippase. It is an ATPase that transports lipid A, the hydrophobic moiety of lipopolysaccharide (LPS), a glucosamine-based saccharolipid that makes up the outer monolayer of the outer membranes of most gram-negative bacteria. Lipid A is an endotoxin and so loss of MsbA from the cell membrane or mutations that disrupt transport results in the accumulation of lipid A in the inner cell membrane resulting to cell death. It is a close bacterial homolog of P-glycoprotein (Pgp) by protein sequence homology and has overlapping substrate specificities with the MDR-ABC transporter LmrA from Lactococcus lactis.[76] MsbA from E. coli is 36% identical to the NH2-terminal half of human MDR1, suggesting a common mechanism for transport of amphiphatic and hydrophobic substrates. The MsbA gene encodes a half transporter that contains a transmembrane domain (TMD) fused with a nucleotide-binding domain (NBD). It is assembled as a homodimer with a total molecular mass of 129.2 kD. MsbA contains 6 TMDs on the periplasmic side, an NBD located on the cytoplasmic side of the cell membrane, and an intracellular domain (ICD), bridging the TMD and NBD. This conserved helix extending from the TMD segments into or near the active site of the NBD is largely responsible for crosstalk between TMD and NBD. In particular, ICD1 serves as a conserved pivot about which the NBD can rotate, therefore allowing the NBD to disassociate and dimerize during ATP binding and hydrolysis.[4][15][18][23][44][54][55][77]

Previously published (and now retracted) X-ray structures of MsbA were inconsistent with the bacterial homolog Sav1866.[78][79] The structures were reexamined and found to have an error in the assignment of the hand resulting to incorrect models of MsbA. Recently, the errors have been rectified and new structures have been reported.[41] The resting state of E. coli MsbA exhibits an inverted "V" shape with a chamber accessible to the interior of the transporter suggesting an open, inward-facing conformation. The dimer contacts are concentrated between the extracellular loops and while the NBDs are ≈50Å apart, the subunits are facing each other. The distance between the residues in the site of the dimer interface have been verified by cross-linking experiments[80] and EPR spectroscopy studies.[81] The relatively large chamber allows it to accommodate large head groups such as that present in lipid A. Significant conformational changes are required to move the large sugar head groups across the membrane. The difference between the two nucleotide-free (apo) structures is the ≈30° pivot of TM4/TM5 helices relative to the TM3/TM6 helices. In the closed apo state (from V. cholerae MsbA), the NBDs are aligned and although closer, have not formed an ATP sandwich, and the P loops of opposing monomers are positioned next to one another. In comparison to the open conformation, the dimer interface of the TMDs in the closed, inward-facing conformation has extensive contacts. For both apo conformations of MsbA, the chamber opening is facing inward. The structure of MsbA-AMP-PNP (5'-adenylyl-β-γ-imidodiphosphate), obtained from S. typhimurium, is similar to Sav1866. The NBDs in this nucleotide-bound, outward-facing conformation, come together to form a canonical ATP dimer sandwich, that is, the nucleotide is situated in between the P-loop and LSGGQ motif. The conformational transition from MsbA-closed-apo to MsbA-AMP-PNP involves two steps, which are more likely concerted: a ≈10° pivot of TM4/TM5 helices towards TM3/TM6, bringing the NBDs closer but not into alignment followed by tilting of TM4/TM5 helices ≈20° out of plane. The twisting motion results in the separation of TM3/TM6 helices away from TM1/TM2 leading to a change from an inward- to an outward- facing conformation. Thus, changes in both the orientation and spacing of the NBDs dramatically rearrange the packing of transmembrane helices and effectively switch access to the chamber from the inner to the outer leaflet of the membrane.[41] The structures determined for MsbA is basis for the tilting model of transport.[18] The structures described also highlight the dynamic nature of ABC exporters as also suggested by fluorescence and EPR studies.[54][81][82] Recent work has resulted in the discovery of MsbA inhibitors.[83][84]
Mechanism of transport for exporters
[edit]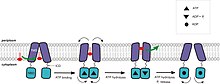
Экспортеры ABC имеют транспортный механизм, который соответствует как модели попеременного доступа, так и модели ATP-переключения. В апо-состояниях экспортеров конформация обращена внутрь, а TMD и NBD расположены относительно далеко друг от друга, что позволяет разместить амфифильные или гидрофобные субстраты. В частности, для MsbA размер камеры достаточно велик, чтобы вместить группы сахаров из липополисахаридов (ЛПС). Как предположили несколько групп, связывание субстрата инициирует транспортный цикл. «Силовой удар», то есть связывание АТФ, которое вызывает димеризацию NBD и образование сэндвича АТФ, вызывает конформационные изменения в ДВНЧС. В MsbA группы сахарных головок изолируются внутри камеры во время «рабочего хода». Полость выстлана заряженными и полярными остатками, которые, вероятно, сольватируются, создавая энергетически невыгодную среду для гидрофобных субстратов и энергетически выгодную для полярных фрагментов амфифильных соединений или сахарных групп ЛПС. Поскольку липид не может быть стабильным в течение длительного времени в среде камеры, липид А и другие гидрофобные молекулы могут «перевернуться» в энергетически более выгодное положение внутри листка внешней мембраны. «Переворот» также может быть вызван сдвигом твердого тела TMD, в то время как гидрофобные хвосты LPS протаскиваются через липидный бислой. Переупаковка спиралей переводит конформацию во внешнее состояние. Гидролиз АТФ может расширить периплазматическое отверстие и подтолкнуть субстрат к внешнему листку липидного бислоя. Гидролиз второй молекулы АТФ и высвобождение P i отделяет НБД с последующим восстановлением состояния покоя, открывая камеру в сторону цитоплазмы для следующего цикла. [41] [44] [52] [55] [78] [79] [81] [85]
Роль в множественной лекарственной устойчивости
[ редактировать ]Известно, что транспортеры ABC играют решающую роль в развитии множественной лекарственной устойчивости (МЛУ). При МЛУ у пациентов, принимающих лекарства, в конечном итоге развивается устойчивость не только к препарату, который они принимают, но и к нескольким различным типам лекарств. Это вызвано несколькими факторами, одним из которых является усиленное выведение препарата из клетки транспортерами ABC. Например, белок ABCB1 ( P-гликопротеин ) участвует в выкачивании из клетки препаратов, подавляющих опухоль. Pgp, также называемый MDR1, ABCB1, является прототипом транспортеров ABC, а также наиболее широко изученным геном. Известно, что Pgp переносит органические катионные или нейтральные соединения. Было также продемонстрировано, что несколько членов семейства ABCC, также известных как MRP, придают МЛУ органическим анионным соединениям. Наиболее изученным членом семейства ABCG является ABCG2, также известный как BCRP (белок устойчивости к раку молочной железы), который обеспечивает устойчивость к большинству ингибиторов топоизомеразы I или II, таких как топотекан, иринотекан и доксорубицин.
Неясно, как именно эти белки могут перемещать такое большое количество лекарств, однако одна модель (модель гидрофобного пылесоса) утверждает, что в P-гликопротеине лекарства без разбора связываются с липидной фазой на основании их гидрофобности.
Открытие первого эукариотического белка-переносчика ABC произошло в результате исследований опухолевых клеток и культивируемых клеток, которые проявляли устойчивость к нескольким лекарствам с несвязанной химической структурой. Было показано, что эти клетки экспрессируют повышенные уровни транспортного белка множественной лекарственной устойчивости (MDR), который первоначально назывался P-гликопротеином (P-gp), но его также называют белком 1 множественной лекарственной устойчивости (MDR1) или ABCB1. Этот белок использует гидролиз АТФ , как и другие транспортеры ABC, для экспорта большого количества лекарств из цитозоля во внеклеточную среду. В клетках с множественной лекарственной устойчивостью ген MDR1 часто амплифицируется. Это приводит к значительному перепроизводству белка MDR1. Субстраты ABCB1 млекопитающих представляют собой в основном плоские жирорастворимые молекулы с одним или несколькими положительными зарядами. Все эти субстраты конкурируют друг с другом за транспорт, что позволяет предположить, что они связываются с одними и теми же или перекрывающимися сайтами белка. Многие из лекарств, транспортируемых ABCB1, представляют собой небольшие неполярные лекарства, которые диффундируют через из внеклеточной среды в цитозоль, где они блокируют различные клеточные функции. Такие лекарства, как колхицин и винбластин , которые блокируют сборку микротрубочек, свободно проникают через мембрану в цитозоль, но экспорт этих препаратов с помощью ABCB1 снижает их концентрацию в клетке. Следовательно, для уничтожения клеток, экспрессирующих ABCB1, требуется более высокая концентрация лекарств, чем для тех, которые не экспрессируют этот ген. [10]
Другими переносчиками ABC, которые способствуют множественной лекарственной устойчивости, являются ABCC1 (MRP1) и ABCG2 (белок устойчивости к раку молочной железы). [86]
Для решения проблем, связанных с множественной лекарственной устойчивостью MDR1, можно использовать разные типы лекарств или необходимо ингибировать сами ABC-транспортеры. Чтобы другие типы лекарств подействовали, они должны обойти механизм резистентности, которым является транспортер ABC. Для этого можно использовать другие противораковые препараты, такие как алкилирующие препараты ( циклофосфамид ), антиметаболиты ( 5-фторурацил ) и модифицированные антрациклином препараты ( аннамицин и доксорубицин -пептид). Эти лекарства не будут функционировать в качестве субстрата транспортеров ABC и, следовательно, не будут транспортироваться. Другой вариант — одновременное использование комбинации препаратов, ингибирующих ABC, и противораковых препаратов. Это могло бы обратить вспять резистентность к противораковым препаратам, чтобы они могли функционировать по назначению. Субстраты, которые обращают устойчивость к противораковым препаратам, называются химиосенсибилизаторами. [8]
Обращение множественной лекарственной устойчивости
[ редактировать ]Лекарственная устойчивость — распространенная клиническая проблема, возникающая у пациентов с инфекционными заболеваниями и у больных раком. Прокариотические и эукариотические микроорганизмы, а также неопластические клетки часто оказываются устойчивыми к лекарствам. МЛУ часто связана со сверхэкспрессией транспортеров ABC. Ингибирование транспортеров ABC низкомолекулярными соединениями широко исследовалось у онкологических больных; однако клинические результаты оказались разочаровывающими. Недавно различные стратегии RNAi были применены для обращения вспять MDR в различных моделях опухолей, и эта технология эффективна в обращении вспять MDR, опосредованной ABC-транспортером, в раковых клетках и, следовательно, является многообещающей стратегией для преодоления MDR с помощью генной терапии. Технологию РНКи также можно рассматривать для преодоления МЛУ при инфекционных заболеваниях, вызванных микробными патогенами. [87]
Физиологическая роль
[ редактировать ]Помимо обеспечения МЛУ в опухолевых клетках, транспортеры ABC также экспрессируются в мембранах здоровых клеток, где они облегчают транспорт различных эндогенных веществ, а также веществ, чужеродных для организма. Например, транспортеры ABC, такие как Pgp, MRP и BCRP, ограничивают всасывание многих лекарств из кишечника и перекачивают лекарства из клеток печени в желчь. [88] как средство выведения инородных веществ из организма. Большое количество наркотиков либо транспортируются самими транспортерами ABC, либо влияют на транспортировку других наркотиков. Последний сценарий может привести к лекарственному взаимодействию . [89] иногда это приводит к изменению действия лекарств. [90]
Методы характеристики взаимодействия транспортеров ABC
[ редактировать ]Существует ряд типов анализов, которые позволяют обнаружить взаимодействие транспортера ABC с эндогенными и ксенобиотическими соединениями. [91] Сложность анализа варьируется от относительно простых мембранных анализов. [92] например, анализ везикулярного транспорта, анализ АТФазы и более сложные клеточные анализы, вплоть до сложных in vivo. Джеффри П., Саммерфилд С.Г. (2007). «Проблемы скрининга гематоэнцефалического барьера (ГЭБ)». Ксенобиотика . 37 (10–11): 1135–51. дои : 10.1080/00498250701570285 . ПМИД 17968740 . S2CID 25944548 . методологии обнаружения. [93]
Мембранные анализы
[ редактировать ]Анализ везикулярного транспорта обнаруживает транслокацию молекул транспортерами ABC. [94] Мембраны, приготовленные в подходящих условиях, содержат ориентированные изнутри наружу везикулы, причем сайт связывания АТФ и сайт связывания субстрата транспортера обращены к буферу снаружи. Субстраты транспортера поглощаются пузырьками АТФ-зависимым образом. Для отделения везикул от инкубационного раствора используют быструю фильтрацию с использованием фильтров из стекловолокна или нитроцеллюлозных мембран, а тестируемое соединение, захваченное внутри везикул, удерживается на фильтре. Количество транспортированных немеченых молекул определяют методами ВЭЖХ, ЖХ/МС, ЖХ/МС/МС. Альтернативно, соединения имеют радиоактивную метку, флуоресцируют или имеют флуоресцентную метку, так что радиоактивность или флуоресценцию, сохраняющуюся на фильтре, можно определить количественно.
В исследованиях везикулярного транспорта используются различные типы мембран из разных источников (например, клетки насекомых, трансфицированные или отобранные клеточные линии млекопитающих). Мембраны коммерчески доступны или могут быть получены из различных клеток или даже тканей, например мембран канальцев печени. Преимущество этого типа анализа заключается в измерении фактического распределения субстрата через клеточную мембрану. Его недостатком является то, что соединения со средней и высокой пассивной проницаемостью не удерживаются внутри везикул, что затрудняет проведение прямых измерений транспорта с этим классом соединений.
Анализ везикулярного транспорта можно проводить в «непрямом» режиме, когда взаимодействующие тестируемые лекарственные средства модулируют скорость транспорта репортерного соединения. Этот тип анализа особенно подходит для обнаружения возможных взаимодействий лекарственного средства и эндогенного субстрата. Он не чувствителен к пассивной проницаемости соединений и поэтому обнаруживает все взаимодействующие соединения. Тем не менее, он не дает информации о том, является ли тестируемое соединение ингибитором переносчика или субстратом переносчика, конкурентно ингибирующим его функцию. Типичным примером непрямого анализа везикулярного транспорта является обнаружение ингибирования транспорта таурохолата с помощью ABCB11 ( BSEP ).
Цельноклеточные анализы
[ редактировать ]Клетки, экспрессирующие транспортер оттока, активно выкачивают субстраты из клетки, что приводит к более низкой скорости накопления субстрата, более низкой внутриклеточной концентрации в равновесном состоянии или более высокой скорости элиминации субстрата из клеток, нагруженных субстратом. Переносимые радиоактивные субстраты или меченые флуоресцентные красители можно измерить напрямую, или косвенно можно определить модуляцию накопления субстрата-зонда (например, флуоресцентных красителей, таких как родамин 123 или кальцеин) в присутствии тестируемого лекарственного средства. [89]
Кальцеин-АМ. Высокопроницаемое производное кальцеина легко проникает в интактные клетки, где эндогенные эстеразы быстро гидролизуют его до флуоресцентного кальцеина. В отличие от кальцеина-АМ, кальцеин имеет низкую проницаемость и поэтому задерживается в клетке и накапливается. Поскольку кальцеин-AM является отличным субстратом отходящих транспортеров MDR1 и MRP1, клетки, экспрессирующие транспортеры MDR1 и/или MRP1, выкачивают кальцеин-AM из клетки до того, как эстеразы смогут его гидролизовать. Это приводит к снижению скорости клеточного накопления кальцеина. Чем выше активность MDR в клеточной мембране, тем меньше кальцеина накапливается в цитоплазме. В клетках, экспрессирующих MDR, добавление в избытке ингибитора MDR или субстрата MDR резко увеличивает скорость накопления кальцеина. Активность мультилекарственного переносчика отражается в разнице между количеством красителя, накопленного в присутствии и в отсутствие ингибитора. Используя селективные ингибиторы, можно легко различить транспортную активность MDR1 и MRP1. Этот анализ можно использовать для проверки лекарств на взаимодействие с транспортерами, а также для количественной оценки активности клеток с МЛУ. Анализ кальцеина является запатентованным анализом компании SOLVO Biotechnology.
Подсемейства
[ редактировать ]Подсемейства млекопитающих
[ редактировать ]У человека имеется 49 известных транспортеров ABC, которые Организация генома человека классифицирует на семь семейств.
| Семья | Члены | Функция | Примеры |
|---|---|---|---|
| АБКА | Это семейство содержит одни из крупнейших переносчиков (длиной более 2100 аминокислот). Пять из них расположены кластером в хромосоме 17q24. | Отвечает, среди прочего, за транспортировку холестерина и липидов. | АБСА12 АБСА1 |
| АВСВ | Состоит из 4 полных и 7 половинных транспортеров. | Некоторые из них расположены, например, в гематоэнцефалическом барьере, печени, митохондриях, транспортируют пептиды и желчь. | АВСВ5 |
| АВСС | Состоит из 12 полноценных транспортеров. | Используется в транспорте ионов, рецепторах клеточной поверхности, секреции токсинов. Включает белок CFTR, которого вызывает муковисцидоз . дефицит | АВСС6 |
| АВСD | Состоит из 4 полутранспортеров. | Все они используются в пероксисомах . | ABCD1 |
| ABCE/ABCF | Состоит из 1 белка ABCE и 3 белков ABCF. | На самом деле это не транспортеры, а просто АТФ-связывающие домены, происходящие из семейства ABC, но без трансмембранных доменов. Эти белки в основном регулируют синтез или экспрессию белков. | ABCE1 , ABCF1 , ABCF2 |
| АБЦГ | Состоит из 6 «реверсивных» полутранспортеров, с НБФ на НХ 3. + конец и TM на конце COO-. | Транспортирует липиды, различные лекарственные субстраты, желчь, холестерин и другие стероиды. | АВСG2 АВСG1 |
Полный список человеческих переносчиков ABC можно найти на сайте . [95]
АБКА
[ редактировать ]Подсемейство ABCA состоит из 12 полноценных транспортеров, разделенных на две подгруппы. Первая подгруппа состоит из семи генов, которые соответствуют шести различным хромосомам . Это ABCA1 , ABCA2 , ABCA3 и ABCA4 , ABCA7 , ABCA12 и ABCA13 . Другая подгруппа состоит из ABCA5 и ABCA6 , а также ABCA8 , ABCA9 и ABCA10 . А8-10.Вся подгруппа 2 организована в кластер хромосом от головы до хвоста на хромосоме 17q 24. Гены этой второй подгруппы отличаются от ABCA1-подобных генов наличием 37-38 экзонов в отличие от 50 экзонов в ABCA1. Подгруппа ABCA1 участвует в развитии генетических заболеваний. При рецессивной болезни Танжера белок ABCA1 мутирует. Кроме того, ABCA4 соответствует участку хромосомы 1p21, который содержит ген болезни Штаргардта. Обнаружено, что этот ген высоко экспрессируется в палочковидных фоторецепторах и мутирует при болезни Штаргардта, рецессивном пигментном ретините и большинстве случаев рецессивной колбочко-палочковидной дистрофии. [9]
АВСВ
[ редактировать ]Подсемейство ABCB состоит из четырех полных транспортеров и двух полутранспортеров. Это единственное человеческое подсемейство, имеющее как половинные, так и полные типы транспортеров. ABCB1 был обнаружен как белок, сверхэкспрессируемый в некоторых опухолевых клетках, устойчивых к лекарствам. Он экспрессируется в первую очередь в гематоэнцефалическом барьере и печени и, как полагают, участвует в защите клеток от токсинов. Клетки, которые сверхэкспрессируют этот белок, обладают множественной лекарственной устойчивостью . [9]
АВСС
[ редактировать ]Подсемейство ABCC содержит тринадцать членов, и девять из этих переносчиков называются белками множественной лекарственной устойчивости (MRP). Белки MRP встречаются повсюду в природе и выполняют множество важных функций. [96] Известно, что они участвуют в транспорте ионов, секреции токсинов и передаче сигналов. [9] Из девяти белков MRP четыре из них, MRP4, 5, 8, 9 (ABCC4, 5, 11 и 12), имеют типичную структуру ABC с четырьмя доменами, состоящими из двух перекрывающих мембрану доменов, за каждым протягивающим доменом следует нуклеотидсвязывающий домен. Их называют короткими MRP. Остальные 5 MRP (MRP1, 2, 6, 7) (ABCC1, 2, 3, 6 и 10) известны как длинные MRP и имеют дополнительный пятый домен на своем N-конце . [96]
CFTR , транспортер, участвующий в заболевании муковисцидозом , также считается частью этого подсемейства. Муковисцидоз возникает при мутации и потере функции CFTR. [9]
Рецепторы сульфонилмочевины (SUR) , участвующие в секреции инсулина, функциях нейронов и мышцах, также являются частью этого семейства белков. Мутации в белках SUR являются потенциальной причиной неонатального сахарного диабета . SUR также является местом связывания таких препаратов, как сульфонилмочевины и активаторов открытия калиевых каналов, таких как диазоксид .
АВСD
[ редактировать ]Подсемейство ABCD состоит из четырех генов, которые кодируют полупереносчики, экспрессируемые исключительно в пероксисоме . ABCD1 отвечает за Х-сцепленную форму адренолейкодистрофии (АЛД), которая представляет собой заболевание, характеризующееся нейродегенерацией и недостаточностью надпочечников, которое обычно начинается в позднем детстве. В клетках больных АЛД наблюдается накопление неразветвленных насыщенных жирных кислот, однако точная роль ABCD1 в этом процессе до сих пор не определена. Кроме того, функция других генов ABCD еще не определена, но считается, что они оказывают сходные функции в метаболизме жирных кислот . [9]
ABCE и ABCF
[ редактировать ]Обе эти подгруппы состоят из генов, которые имеют АТФ-связывающие домены, тесно связанные с другими транспортерами ABC, но эти гены не кодируют трансмембранные домены. ABCE состоит только из одного члена, OABP или ABCE1 , который, как известно, распознает определенные олигодендроциты, образующиеся в ответ на определенные вирусные инфекции. Каждый член подгруппы ABCF состоит из пары доменов связывания АТФ. [9]
АБЦГ
[ редактировать ]Шесть полупереносчиков с сайтами связывания АТФ на N-конце и трансмембранными доменами на C-конце составляют подсемейство ABCG. Эта ориентация противоположна всем другим генам ABC. В геноме человека всего 5 генов ABCG, но в геноме дрозофилы их 15, а в дрожжах — 10. Ген ABCG2 был обнаружен в клеточных линиях, отобранных по высокому уровню устойчивости к митоксантрону и отсутствию экспрессии ABCB1 или ABCC1 . ABCG2 может экспортировать противораковые препараты антрациклина , а также топотекан , митоксантрон или доксорубицин в качестве субстратов. хромосомные транслокации вызывают амплификацию или перестройку ABCG2, обнаруженную в устойчивых клеточных линиях. Было обнаружено, что [9]
Межвидовые подсемейства
[ редактировать ]В этом разделе отсутствует информация о сопоставлении Pfam/InterPro (сложно сделать, они нужны и для улучшения данных). ( декабрь 2020 г. ) |
В TCDB построена следующая система классификации трансмембранных переносчиков растворенных веществ. [97]
Три семьи экспортеров ABC определяются своим эволюционным происхождением. [6] Экспортеры ABC1 эволюционировали путем внутригенного трипликации предшественника 2 TMS (TMS = трансмембранный сегмент. Белок «2 TMS» имеет 2 трансмембранных сегмента), чтобы дать 6 белков TMS. Экспортеры ABC2 возникли в результате внутригенного дупликации предшественника 3 TMS, а экспортеры ABC3 произошли от предшественника 4 TMS, который дублировался либо экстрагенно, чтобы дать два белка 4 TMS, оба необходимые для транспортной функции, либо внутригенно, чтобы дать 8 или 10 белков TMS. По-видимому, 10 белков TMS имеют два дополнительных TMS между двумя 4 повторными единицами TMS. [98] Большинство систем поглощения (все, кроме 3.A.1.21) относятся к типу ABC2, разделенному на тип I и тип II по способу обращения с нуклеотидами. Особое подсемейство импортеров ABC2, называемое ECF, использует отдельную подъединицу для распознавания субстрата. [99]
ABC1 ( ИнтерПро : IPR036640 ):
- 3.A.1.106 Семейство экспортеров липидов (LipidE)
- 3.A.1.108 Семейство экспортеров β-глюканов (GlucanE)
- 3.A.1.109 Семейство экспортеров белка-1 (Prot1E)
- 3.A.1.110 Семейство экспортеров белка-2 (Prot2E)
- 3.A.1.111 Семейство экспортеров пептида-1 (Pep1E)
- 3.A.1.112 Семейство экспортеров пептида-2 (Pep2E)
- 3.A.1.113 Семейство экспортеров пептида-3 (Pep3E)
- 3.A.1.117 Семья экспортеров наркотиков-2 (DrugE2)
- 3.A.1.118 Семейство экспортеров Microcin J25 (McjD)
- 3.A.1.119 Семейство экспортеров лекарств/сидерофоров-3 (DrugE3)
- 3.A.1.123 Семейство экспортеров пептида-4 (Pep4E)
- 3.A.1.127 Семейство экспортеров пептидов AmfS (AmfS-E)
- 3.A.1.129 Семейство экспортеров цистеина CydDC (CydDC-E)
- 3.A.1.135 Семья экспортеров наркотиков-4 (DrugE4)
- 3.A.1.139 Семейство экспортеров UDP-глюкозы (U-GlcE) (семейство UPF0014)
- 3.A.1.201 Семейство экспортеров множественной лекарственной устойчивости (MDR) (ABCB)
- 3.A.1.202 Семейство экспортеров трансмембранной проводимости муковисцидоза (CFTR) (ABCC)
- 3.A.1.203 Семейство пероксисомальных транспортеров жирных ацил-КоА (P-FAT) (ABCD)
- 3.A.1.206 Семейство экспортеров половых феромонов (STE) a-фактора (ABCB)
- 3.A.1.208 Семейство переносчиков лекарственных конъюгатов (DCT) (ABCC) (Dębska et al., 2011)
- 3.A.1.209 Семейство переносчиков пептидов MHC (TAP) (ABCB)
- 3.A.1.210 Семейство транспортеров тяжелых металлов (HMT) (ABCB)
- 3.A.1.212 Семейство экспортеров митохондриальных пептидов (MPE) (ABCB)
- сидерофор-Fe3+ 3.A.1.21 Семейство переносчиков поглощения (SIUT)
ABC2 ( InterPro : IPR000412 [частично]):
- 3.A.1.101 Семейство экспортеров капсульных полисахаридов (CPSE)
- 3.A.1.102 Семейство экспортеров липоолигосахаридов (LOSE)
- 3.A.1.103 Семейство экспортеров липополисахаридов (LPSE)
- 3.A.1.104 Семья экспортеров тейхоевой кислоты (TAE)
- 3.A.1.105 Семья экспортеров наркотиков-1 (DrugE1)
- 3.A.1.107 Семейство предполагаемых экспортеров гема (HemeE)
- 3.A.1.115 Семейство экспортеров Na+ (NatE)
- 3.A.1.116 Семейство экспортеров Microcin B17 (McbE)
- 3.A.1.124 Семейство трехкомпонентных экспортеров пептида-5 (Pep5E)
- 3.A.1.126 Семейство экспортеров β-экзотоксина I (βETE)
- 3.A.1.128 Семейство экспортеров пептидов SkfA (SkfA-E)
- 3.A.1.130 Семейство экспортеров нескольких лекарств/гемолизина (MHE)
- 3.A.1.131 Семейство резистентности к бацитрацину (Bcr)
- 3.A.1.132 Семейство транспортеров ABC Gliding Motility (Gld)
- 3.A.1.133 Семейство экспортеров пептида-6 (Pep6E)
- 3.A.1.138 Неизвестное семейство типа ABC-2 (ABC2-1).
- 3.A.1.141 Семейство экспортеров этилвиологена (EVE) (семейство DUF990; InterPro : IPR010390 )
- 3.A.1.142 Семейство гликолипид-флиппаз (GLFlippase)
- 3.A.1.143 Система секреции экзопротеинов (EcsAB(C))
- 3.A.1.144: Функционально нехарактеризованное семейство ABC2-1 (ABC2-1)
- 3.A.1.145: Слитое с пептидазой функционально неохарактеризованное семейство ABC2-2 (ABC2-2)
- 3.A.1.146: Семейство экспортеров актинородина (ACT) и ундецилпродигиозина (RED) (ARE).
- 3.A.1.147: Функционально нехарактеризованное семейство ABC2-2 (ABC2-2)
- 3.A.1.148: Функционально нехарактеризованное семейство ABC2-3 (ABC2-3)
- 3.A.1.149: Функционально нехарактеризованное семейство ABC2-4 (ABC2-4)
- 3.A.1.150: Функционально нехарактеризованное семейство ABC2-5 (ABC2-5)
- 3.A.1.151: Функционально нехарактеризованное семейство ABC2-6 (ABC2-6)
- 3.A.1.152: Семейство экспортных липополисахаридов (LptBFG) ( InterPro : IPR005495 )
- 3.A.1.204 Семейство транспортеров предшественников глазного пигмента (EPP) (ABCG)
- 3.A.1.205 Семейство плейотропной лекарственной устойчивости (PDR) (ABCG)
- 3.A.1.211 Семейство холестерин/фосфолипид/ретиналь (CPR) флиппаз (ABCA)
- 9.B.74 Семейство белков фаговой инфекции (PIP)
- все системы поглощения (3.A.1.1 – 3.A.1.34, кроме 3.A.1.21)
- 3.A.1.1 Транспортер поглощения углеводов-1 (CUT1)
- 3.A.1.2 Транспортер поглощения углеводов-2 (CUT2)
- 3.A.1.3 Транспортер поглощения полярных аминокислот (PAAT)
- 3.A.1.4 Гидрофобный переносчик поглощения аминокислот (HAAT)
- 3.A.1.5 Транспортер поглощения пептидов/опинов/никеля (PepT)
- 3.A.1.6 Сульфат/вольфраматный переносчик поглощения (SulT)
- 3.A.1.7 Транспортер поглощения фосфата (PhoT)
- 3.A.1.8 Транспортер поглощения молибдата (MolT)
- 3.A.1.9 Транспортер поглощения фосфоната (PhnT)
- 3.A.1.10 Транспортер поглощения трехвалентного железа (FeT)
- 3.A.1.11 Транспортер поглощения полиаминов/опинов/фосфонатов (POPT)
- 3.A.1.12 Транспортер поглощения четвертичного амина (QAT)
- 3.A.1.13 Транспортер поглощения витамина B12 ( B12T)
- 3.A.1.14 Транспортер поглощения хелата железа (FeCT)
- 3.A.1.15 Хелатный транспортер марганца/цинка/железа (MZT)
- 3.A.1.16 Транспортер поглощения нитратов/нитритов/цианатов (NitT)
- 3.A.1.17 Транспортер поглощения таурина (TauT)
- 3.A.1.19 Транспортер захвата тиамина (ThiT)
- 3.A.1.20 Транспортер железа Brachyspira (BIT)
- 3.A.1.21 Транспортер поглощения сидерофора-Fe3+ (SIUT)
- 3.A.1.24 Семейство транспортеров захвата метионина (MUT) (аналогично 3.A.1.3 и 3.A.1.12)
- 3.A.1.27 Семейство γ-гексахлорциклогексана (ГХГ) (аналогично 3.A.1.24 и 3.A.1.12)
- 3.A.1.34 Семейство триптофана (TrpXYZ)
- Системы поглощения ECF
- 3.A.1.18 Семейство транспортеров поглощения кобальта (CoT)
- 3.A.1.22 Семейство транспортеров поглощения никеля (NiT)
- 3.A.1.23 Семейство переносчиков поглощения никеля/кобальта (NiCoT)
- 3.A.1.25 Семейство транспортеров поглощения биотина (BioMNY)
- 3.A.1.26 Семейство предполагаемых транспортеров захвата тиамина (ThiW)
- 3.A.1.28 Семейство кевозина (кеуозин)
- 3.A.1.29 Семейство предшественников метионина (Met-P)
- 3.A.1.30 Семейство предшественников тиамина (Thi-P)
- 3.A.1.31 Семейство Unknown-ABC1 (U-ABC1)
- 3.A.1.32 Семейство предшественников кобаламина (B12-P)
- 3.A.1.33 Семейство метилтиоаденозина (МТА)
ABC3 ( ИнтерПро : IPR003838 ):
- 3.A.1.114 Семейство вероятных экспортеров гликолипидов (DevE)
- 3.A.1.122 Семейство экспортеров макролидов (MacB)
- 3.A.1.125 Семейство липопротеинтранслоказ (LPT)
- 3.A.1.134 Семейство экспортеров пептида-7 (Pep7E)
- 3.A.1.136 Неохарактеризованное семейство типа ABC-3 (U-ABC3-1)
- 3.A.1.137 Неохарактеризованное семейство типа ABC-3 (U-ABC3-2)
- 3.A.1.140 Семейство септаций FtsX/FtsE (FtsX/FtsE)
- 3.A.1.207 Семейство эукариот ABC3 (E-ABC3)
Изображения
[ редактировать ]В последние годы создано множество структур водорастворимых доменов белков ABC. [2]
См. также
[ редактировать ]- АТФ-связывающий домен транспортеров ABC
- Трансмембранный домен транспортеров ABC
- Элизабет П. Карпентер , британский структурный биолог, первой описавшая структуру человеческого ABC-транспортера ABC10.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Фатх, MJ; Колтер, Р. (декабрь 1993 г.). «АВС-транспортеры: бактериальные экспортеры» . Микробиологические обзоры . 57 (4): 995–1017. дои : 10.1128/MMBR.57.4.995-1017.1993 . ISSN 0146-0749 . ПМК 372944 . ПМИД 8302219 .
- ^ Перейти обратно: а б Джонс П.М., Джордж А.М. (март 2004 г.). «Структура и механизм транспортера ABC: перспективы последних исследований» . Клеточные и молекулярные науки о жизни . 61 (6): 682–99. дои : 10.1007/s00018-003-3336-9 . ПМЦ 11138499 . ПМИД 15052411 . S2CID 21422822 .
- ^ Понте-Сукре А, изд. (2009). ABC-транспортеры в микроорганизмах . Кайстер Академик. ISBN 978-1-904455-49-3 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Дэвидсон А.Л., Дасса Э., Орел С., Чен Дж. (июнь 2008 г.). «Структура, функции и эволюция бактериальных АТФ-связывающих кассетных систем» . Обзоры микробиологии и молекулярной биологии . 72 (2): 317–64, оглавление. дои : 10.1128/MMBR.00031-07 . ПМЦ 2415747 . ПМИД 18535149 .
- ^ Перейти обратно: а б с д Гоффо А., де Хертог Б., Барет П.В. (2013). «АВС Транспортерс». В Лейн У.Дж., Леннарц, доктор медицинских наук (ред.). Энциклопедия биологической химии (второе изд.). Лондон: Академическая пресса. стр. 7–11. дои : 10.1016/B978-0-12-378630-2.00224-3 . ISBN 978-0-12-378631-9 .
- ^ Перейти обратно: а б Ван Б., Дукаревич М., Сунь Э.И., Йен М.Р., Сайер М.Х. (сентябрь 2009 г.). «Мембранные портеры АТФ-связывающих кассетных транспортных систем полифилетичны» . Журнал мембранной биологии . 231 (1): 1–10. дои : 10.1007/s00232-009-9200-6 . ПМК 2760711 . ПМИД 19806386 .
- ^ тер Бик Дж., Гуськов А., Slotboom DJ (апрель 2014 г.). «Структурное разнообразие транспортеров ABC» . Журнал общей физиологии . 143 (4): 419–35. дои : 10.1085/jgp.201411164 . ПМЦ 3971661 . ПМИД 24638992 .
- ^ Перейти обратно: а б с Чой Ч. (октябрь 2005 г.). «Транспортеры ABC как механизмы множественной лекарственной устойчивости и разработка химиосенсибилизаторов для их обращения» . Международная организация раковых клеток . 5:30 . дои : 10.1186/1475-2867-5-30 . ПМЦ 1277830 . ПМИД 16202168 .
- ^ Перейти обратно: а б с д и ж г час я Дин М., Хамон Ю., Чимини Дж. (июль 2001 г.). «Суперсемейство транспортеров АТФ-связывающей кассеты человека (ABC)» . Журнал исследований липидов . 42 (7): 1007–17. дои : 10.1016/S0022-2275(20)31588-1 . ПМИД 11441126 .
- ^ Перейти обратно: а б с д Скотт М.П., Лодиш Х.Ф., Берк А., Кайзер, К., Кригер М., Бретчер А., Плох Х., Амон А. (2012). Молекулярно-клеточная биология . Сан-Франциско: WH Freeman. ISBN 978-1-4292-3413-9 .
- ^ Хендерсон Д.П., Пейн С.М. (ноябрь 1994 г.). «Системы транспорта железа Vibrio cholerae: роль транспорта железа по гему и сидерофору в вирулентности и идентификация гена, связанного с множественными системами транспорта железа» . Инфекция и иммунитет . 62 (11): 5120–5. дои : 10.1128/IAI.62.11.5120-5125.1994 . ПМК 303233 . ПМИД 7927795 .
- ^ Кангелози Г.А., Анкенбауэр Р.Г., Нестер Э.В. (сентябрь 1990 г.). «Сахар индуцирует гены вирулентности Agrobacterium через периплазматический связывающий белок и трансмембранный сигнальный белок» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (17): 6708–12. Бибкод : 1990PNAS...87.6708C . дои : 10.1073/pnas.87.17.6708 . ПМК 54606 . ПМИД 2118656 .
- ^ Кемнер Дж. М., Лян X, Нестер Э. В. (апрель 1997 г.). «Ген вирулентности Agrobacterium tumefaciens chvE является частью предполагаемого оперона транспорта сахара ABC-типа» . Журнал бактериологии . 179 (7): 2452–8. дои : 10.1128/jb.179.7.2452-2458.1997 . ПМК 178989 . ПМИД 9079938 .
- ^ Пулман Б., Спитцер Дж.Дж., Вуд Дж.М. (ноябрь 2004 г.). «Бактериальное осмосенсинг: роль структуры мембраны и электростатики в липид-белковых и белок-белковых взаимодействиях» (PDF) . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1666 (1–2): 88–104. дои : 10.1016/j.bbamem.2004.06.013 . ПМИД 15519310 . S2CID 21763870 .
- ^ Перейти обратно: а б с д и ж Дэвидсон А.Л., Чен Дж (2004). «АТФ-связывающие кассетные транспортеры у бактерий». Ежегодный обзор биохимии . 73 : 241–68. doi : 10.1146/annurev.biochem.73.011303.073626 . ПМИД 15189142 .
- ^ Чжоу З, Уайт К.А., Полисси А., Георгопулос С., Раец Ч.Р. (май 1998 г.). «Функция Escherichia coli MsbA, важного транспортера семейства ABC, в биосинтезе липида А и фосфолипидов» . Журнал биологической химии . 273 (20): 12466–75. дои : 10.1074/jbc.273.20.12466 . hdl : 2434/611267 . ПМИД 9575204 .
- ^ Пул Р.К., Гибсон Ф., Ву Джи (апрель 1994 г.). «Продукт гена cydD, компонент гетеродимерного транспортера ABC, необходим для сборки периплазматического цитохрома с и цитохрома bd в Escherichia coli» . Письма FEMS по микробиологии . 117 (2): 217–23. дои : 10.1111/j.1574-6968.1994.tb06768.x . ПМИД 8181727 .
- ^ Перейти обратно: а б с д и ж г час Пол А., Дево П.Ф., Херрманн А. (март 2005 г.). «Функция прокариотических и эукариотических белков ABC в транспорте липидов». Biochimica et Biophysical Acta (BBA) - Молекулярная и клеточная биология липидов . 1733 (1): 29–52. дои : 10.1016/j.bbalip.2004.12.007 . ПМИД 15749056 .
- ^ Рэндольф Дж.Дж. (2001). «Миграция дендритных клеток в лимфатические узлы: цитокины, хемокины и липидные медиаторы». Семинары по иммунологии . 13 (5): 267–74. дои : 10.1006/smim.2001.0322 . ПМИД 11502161 .
- ^ Гедеон С., Бехраван Дж., Корен Г., Пикетт-Миллер М. (2006). «Транспорт глибурида плацентарными транспортерами ABC: последствия воздействия лекарств на плод». Плацента . 27 (11–12): 1096–102. дои : 10.1016/j.placenta.2005.11.012 . ПМИД 16460798 .
- ^ Перейти обратно: а б Скотт, Хейли; Мартинелли, Лилиан М.; Гринспен, Дэвид; Блуаз, Энрико; Коннор, Кристин Л. (24 марта 2022 г.). «Преждевременные роды связаны с повышенной плацентарной экспрессией переносчиков MDR независимо от ИМТ до беременности» . Журнал клинической эндокринологии и метаболизма . 107 (4): 1140–1158. дои : 10.1210/clinem/dgab813 . ISSN 1945-7197 . ПМИД 34748636 . S2CID 243863723 .
- ^ Шуман Х.А. (1982). «Активный транспорт мальтозы в Escherichia coli K12. Роль периплазматического мальтозосвязывающего белка и свидетельство существования сайта узнавания субстрата в цитоплазматической мембране» . Ж. Биол. Хим . 257 (10): 5455–61. дои : 10.1016/S0021-9258(19)83799-7 . ПМИД 7040366 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д Рис, округ Колумбия, Джонсон Э, Левинсон О (март 2009 г.). «Транспортеры ABC: сила перемен» . Nature Reviews Молекулярно-клеточная биология . 10 (3): 218–27. дои : 10.1038/nrm2646 . ПМЦ 2830722 . ПМИД 19234479 .
- ^ Перейти обратно: а б с Лочер К.П., Ли А.Т., Рис, округ Колумбия (май 2002 г.). «Структура BtuCD E. coli: основа архитектуры и механизма транспортера ABC» (PDF) . Наука . 296 (5570): 1091–8. Бибкод : 2002Sci...296.1091L . дои : 10.1126/science.1071142 . ПМИД 12004122 . S2CID 906489 .
- ^ Хворуп Р.Н., Гетц Б.А., Нидерер М., Холленштейн К., Перозо Э., Лохер К.П. (сентябрь 2007 г.). «Асимметрия в структуре белкового комплекса, связывающего транспортер ABC BtuCD-BtuF». Наука . 317 (5843): 1387–90. Бибкод : 2007Sci...317.1387H . дои : 10.1126/science.1145950 . ПМИД 17673622 . S2CID 37232959 .
- ^ Перейти обратно: а б с Доусон Р.Дж., Лочер К.П. (сентябрь 2006 г.). «Структура бактериального мультилекарственного переносчика ABC». Природа . 443 (7108): 180–5. Бибкод : 2006Natur.443..180D . дои : 10.1038/nature05155 . ПМИД 16943773 . S2CID 27132450 .
- ^ Перейти обратно: а б с Холленштайн К., Фрай Д.К., Лохер К.П. (март 2007 г.). «Структура транспортера ABC в комплексе со связывающим его белком». Природа . 446 (7132): 213–6. Бибкод : 2007Natur.446..213H . дои : 10.1038/nature05626 . ПМИД 17322901 . S2CID 4417002 .
- ^ Перейти обратно: а б Олдхэм М.Л., Харе Д., Киочо Ф.А., Дэвидсон А.Л., Чен Дж. (ноябрь 2007 г.). «Кристаллическая структура каталитического промежуточного продукта переносчика мальтозы». Природа . 450 (7169): 515–21. Бибкод : 2007Natur.450..515O . дои : 10.1038/nature06264 . ПМИД 18033289 . S2CID 4384771 .
- ^ Кадаба Н.С., Кайзер Дж.Т., Джонсон Э., Ли А., Рис, округ Колумбия (июль 2008 г.). «Высокоаффинный переносчик метионина ABC E. coli: структура и аллостерическая регуляция» . Наука . 321 (5886): 250–3. Бибкод : 2008Sci...321..250K . дои : 10.1126/science.1157987 . ПМК 2527972 . ПМИД 18621668 .
- ^ Перейти обратно: а б с д Пинкетт Х.В., Ли А.Т., Лам П., Лочер К.П., Рис, округ Колумбия (январь 2007 г.). «Обращенная внутрь конформация предполагаемого транспортера ABC металл-хелатного типа» (PDF) . Наука . 315 (5810): 373–7. дои : 10.1126/science.1133488 . ПМИД 17158291 . S2CID 10531462 .
- ^ Перейти обратно: а б Муди Дж.Э., Миллен Л., Биннс Д., Хант Дж.Ф., Томас П.Дж. (июнь 2002 г.). «Кооперативная, АТФ-зависимая ассоциация нуклеотидсвязывающих кассет во время каталитического цикла переносчиков АТФ-связывающих кассет» . Журнал биологической химии . 277 (24): 21111–4. дои : 10.1074/jbc.C200228200 . ПМК 3516282 . ПМИД 11964392 .
- ^ Хунг Л.В., Ван IX, Никайдо К., Лю П.К., Эймс Г.Ф., Ким Ш.Х. (декабрь 1998 г.). «Кристаллическая структура АТФ-связывающей субъединицы транспортера ABC». Природа . 396 (6712): 703–7. Бибкод : 1998Natur.396..703H . дои : 10.1038/25393 . ПМИД 9872322 . S2CID 204996524 .
- ^ Перейти обратно: а б с Вердон Г., Альберс С.В., Дейкстра Б.В., Дриссен А.Дж., Тунниссен А.М. (июль 2003 г.). «Кристаллические структуры субъединицы АТФазы транспортера глюкозы ABC из Sulfolobus solfataricus: безнуклеотидные и нуклеотидсвязанные конформации». Журнал молекулярной биологии . 330 (2): 343–58. дои : 10.1016/S0022-2836(03)00575-8 . ПМИД 12823973 .
- ^ Перейти обратно: а б Карпович Н., Марцинкевич О., Миллен Л., Юань Ю.Р., Дай П.Л., Маквей К., Томас П.Дж., Хант Дж.Ф. (июль 2001 г.). «Кристаллические структуры АТФ-связывающей кассеты MJ1267 обнаруживают эффект индуцированной подгонки в активном сайте АТФазы транспортера ABC» . Структура . 9 (7): 571–86. дои : 10.1016/S0969-2126(01)00617-7 . ПМИД 11470432 .
- ^ Перейти обратно: а б с д Чен Дж., Лу Дж., Лин Дж., Дэвидсон А.Л., Киочо Ф.А. (сентябрь 2003 г.). «Движение димера АТФ-связывающей кассеты, подобное пинцету, в транспортном цикле ABC» . Молекулярная клетка . 12 (3): 651–61. doi : 10.1016/j.molcel.2003.08.004 . ПМИД 14527411 .
- ^ Перейти обратно: а б с Дидерихс К., Диц Дж., Греллер Г., Мюллер К., Брид Дж., Шнелл К., Фонрайн К., Боос В., Вельте В. (ноябрь 2000 г.). «Кристаллическая структура MalK, субъединицы АТФазы трегалозно-мальтозного переносчика ABC археи Thermococcuslitoralis» . Журнал ЭМБО . 19 (22): 5951–61. дои : 10.1093/emboj/19.22.5951 . ПМК 305842 . ПМИД 11080142 .
- ^ Перейти обратно: а б Годе Р., Уайли, округ Колумбия (сентябрь 2001 г.). «Структура домена ABC-АТФазы человеческого TAP1, транспортера, связанного с процессингом антигена» . Журнал ЭМБО . 20 (17): 4964–72. дои : 10.1093/emboj/20.17.4964 . ПМК 125601 . ПМИД 11532960 .
- ^ Шмитт Л., Бенабдельхак Х., Блайт М.А., Холланд И.Б., Стаббс М.Т. (июль 2003 г.). «Кристаллическая структура нуклеотидсвязывающего домена ABC-транспортера гемолизина B: идентификация вариабельной области внутри спиральных доменов ABC». Журнал молекулярной биологии . 330 (2): 333–42. дои : 10.1016/S0022-2836(03)00592-8 . ПМИД 12823972 .
- ^ Перейти обратно: а б Юань Ю.Р., Блекер С., Марцинкевич О., Миллен Л., Томас П.Дж., Хант Дж.Ф. (август 2001 г.). «Кристаллическая структура АТФ-связывающей кассеты MJ0796. Последствия для структурных последствий гидролиза АТФ в активном центре транспортера ABC» . Журнал биологической химии . 276 (34): 32313–21. дои : 10.1074/jbc.M100758200 . ПМИД 11402022 .
- ^ Перейти обратно: а б с д и ж Смит П.С., Карпович Н., Миллен Л., Муди Дж.Э., Розен Дж., Томас П.Дж., Хант Дж.Ф. (июль 2002 г.). «Связывание АТФ с моторным доменом транспортера ABC приводит к образованию нуклеотидного сэндвич-димера» . Молекулярная клетка . 10 (1): 139–49. дои : 10.1016/S1097-2765(02)00576-2 . ПМЦ 3516284 . ПМИД 12150914 .
- ^ Перейти обратно: а б с д и Уорд А., Рейес С.Л., Ю Дж., Рот С.Б., Чанг Г. (ноябрь 2007 г.). «Гибкость транспортера ABC MsbA: попеременный доступ с изюминкой» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (48): 19005–10. Бибкод : 2007PNAS..10419005W . дои : 10.1073/pnas.0709388104 . ПМК 2141898 . ПМИД 18024585 .
- ^ Перейти обратно: а б Хопфнер К.П., Керхер А., Шин Д.С., Крейг Л., Артур Л.М., Карни Дж.П., Тайнер Дж.А. (июнь 2000 г.). «Структурная биология АТФазы Rad50: АТФ-управляемый конформационный контроль при восстановлении двухцепочечных разрывов ДНК и суперсемейство ABC-АТФазы» . Клетка . 101 (7): 789–800. дои : 10.1016/S0092-8674(00)80890-9 . ПМИД 10892749 . S2CID 18850076 .
- ^ Фетч Э.Э., Дэвидсон А.Л. (июль 2002 г.). «Катализируемое ванадатом фоторасщепление фирменного мотива транспортера АТФ-связывающей кассеты (ABC)» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (15): 9685–90. дои : 10.1073/pnas.152204499 . ПМК 124977 . ПМИД 12093921 .
- ^ Перейти обратно: а б с д Рейес CL, Уорд А, Ю Дж, Чанг Г (февраль 2006 г.). «Структуры MsbA: понимание оттока нескольких лекарств, опосредованного транспортером ABC». Письма ФЭБС . 580 (4): 1042–8. Бибкод : 2006FEBSL.580.1042R . дои : 10.1016/j.febslet.2005.11.033 . ПМИД 16337944 . S2CID 34114828 .
- ^ Амбудкар С.В., Ким И.В., Ся Д., Сауна З.Э. (февраль 2006 г.). «А-петля, новый консервативный субдомен ароматической кислоты, расположенный выше мотива Уокера А в транспортерах ABC, имеет решающее значение для связывания АТФ» . Письма ФЭБС . 580 (4): 1049–55. Бибкод : 2006FEBSL.580.1049A . дои : 10.1016/j.febslet.2005.12.051 . ПМИД 16412422 . S2CID 20550226 .
- ^ Перейти обратно: а б Журжон С, Орель С, Стейнфелс Е, Бланше С, Делеаж Ж, Ди Пьетро А, Жо ЖМ (сентябрь 2001 г.). «Общий механизм гидролиза АТФ в суперсемействах транспортеров ABC и геликаз». Тенденции биохимических наук . 26 (9): 539–44. дои : 10.1016/S0968-0004(01)01907-7 . ПМИД 11551790 .
- ^ Йе Дж., Осборн А.Р., Гролл М., Рапопорт Т.А. (ноябрь 2004 г.). «RecA-подобные моторные АТФазы - уроки структур» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1659 (1): 1–18. дои : 10.1016/j.bbabio.2004.06.003 . ПМИД 15511523 .
- ^ Перейти обратно: а б Зайцева Дж., Дженевейн С., Джампертц Т., Холланд И.Б., Шмитт Л. (июнь 2005 г.). «H662 является основой гидролиза АТФ в нуклеотидсвязывающем домене транспортера ABC HlyB» . Журнал ЭМБО . 24 (11): 1901–10. дои : 10.1038/sj.emboj.7600657 . ПМЦ 1142601 . ПМИД 15889153 .
- ^ Мэгли К.А., Адмирал С.Дж., Хершлаг Д. (август 1996 г.). «Ras-катализируемый гидролиз GTP: новый взгляд на модельные исследования» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (16): 8160–6. Бибкод : 1996PNAS...93.8160M . дои : 10.1073/pnas.93.16.8160 . ПМК 38640 . ПМИД 8710841 .
- ^ Мэтт А., Тари Л.В., Дельбаре Л.Т. (апрель 1998 г.). «Как киназы переносят фосфорильные группы?» . Структура . 6 (4): 413–9. дои : 10.1016/S0969-2126(98)00043-4 . ПМИД 9562560 .
- ^ Перейти обратно: а б Холленштейн К., Доусон Р.Дж., Лочер К.П. (август 2007 г.). «Структура и механизм белков-транспортеров ABC». Современное мнение в области структурной биологии . 17 (4): 412–8. дои : 10.1016/j.sbi.2007.07.003 . ПМИД 17723295 .
- ^ Перейти обратно: а б с д и ж г Хиггинс К.Ф., Линтон К.Дж. (октябрь 2004 г.). «Модель ATP-переключателя для транспортеров ABC». Структурная и молекулярная биология природы . 11 (10): 918–26. дои : 10.1038/nsmb836 . ПМИД 15452563 . S2CID 23058653 .
- ^ Лочер КП (август 2004 г.). «Строение и механизм транспортеров ABC». Современное мнение в области структурной биологии . 14 (4): 426–31. дои : 10.1016/j.sbi.2004.06.005 . ПМИД 15313236 .
- ^ Перейти обратно: а б с д и ж г час Олдхэм М.Л., Дэвидсон А.Л., Чен Дж. (декабрь 2008 г.). «Структурное понимание механизма транспортера ABC» . Современное мнение в области структурной биологии . 18 (6): 726–33. дои : 10.1016/j.sbi.2008.09.007 . ПМЦ 2643341 . ПМИД 18948194 .
- ^ Перейти обратно: а б с д Чанг Джи (ноябрь 2003 г.). «АВС-транспортеры множественной лекарственной устойчивости» . Письма ФЭБС . 555 (1): 102–5. Бибкод : 2003FEBSL.555..102C . дои : 10.1016/S0014-5793(03)01085-8 . ПМИД 14630327 . S2CID 24228062 .
- ^ Старший А.Е., депутат Кнессета аль-Шави, Урбач, Иллинойс (декабрь 1995 г.). «Каталитический цикл Р-гликопротеина». Письма ФЭБС . 377 (3): 285–9. Бибкод : 1995FEBSL.377..285S . дои : 10.1016/0014-5793(95)01345-8 . ПМИД 8549739 . S2CID 20395778 .
- ^ Симпсон, Брент В.; Пахил, Каранбир С.; Оуэнс, Тристан В.; Лундштедт, Эмили А.; Дэвис, Ребекка М.; Кан, Дэниел; Руис, Нативидад (20 августа 2019 г.). «Сочетание мутаций, которые ингибируют два различных этапа цикла гидролиза АТФ, восстанавливает функцию дикого типа в переносчике липополисахаридов и показывает, что связывание АТФ запускает транспорт» . мБио . 10 (4): e01931–19, /mbio/10/4/mBio.01931–19.atom. дои : 10.1128/mBio.01931-19 . ПМК 6703430 . ПМИД 31431556 .
- ^ Мартин С., Хиггинс С.Ф., Каллаган Р. (декабрь 2001 г.). «Сайт связывания винбластина принимает конформации с высоким и низким сродством во время транспортного цикла P-гликопротеина». Биохимия . 40 (51): 15733–42. дои : 10.1021/bi011211z . ПМИД 11747450 .
- ^ Манчу Л., Чанг Х.Б., Буйсе Ф., Хоу Ю.С., Густот А., Риордан-младший, Рейсшерт Дж.М. (январь 2003 г.). «Промежуточные структурные состояния, участвующие в транспорте лекарств, опосредованном MRP1. Роль глутатиона» . Журнал биологической химии . 278 (5): 3347–56. дои : 10.1074/jbc.M207963200 . ПМИД 12424247 .
- ^ Креймер Д.И., Чай К.П., Ферро-Луцци Эймс Дж. (ноябрь 2000 г.). «Неэквивалентность нуклеотидсвязывающих субъединиц транспортера ABC, гистидинпермеазы и конформационные изменения в мембранном комплексе». Биохимия . 39 (46): 14183–95. дои : 10.1021/bi001066 . ПМИД 11087367 .
- ^ Вигано С., Марголлес А., ван Вин Х.В., Конингс В.Н., Рейсшерт Дж.М. (апрель 2000 г.). «Изменения вторичной и третичной структуры восстановленного LmrA, индуцированные связыванием или гидролизом нуклеотидов. Инфракрасная спектроскопия полного отражения с Фурье-преобразованием и анализ тушения флуоресценции триптофана» (PDF) . Журнал биологической химии . 275 (15): 10962–7. дои : 10.1074/jbc.275.15.10962 . ПМИД 10753896 . S2CID 33274934 .
- ^ Сонво Н., Вигано С., Шапиро А.Б., Линг В., Рейсшерт Дж.М. (июнь 1999 г.). «Опосредованные лигандами изменения третичной структуры восстановленного P-гликопротеина. Анализ тушения флуоресценции триптофана» . Журнал биологической химии . 274 (25): 17649–54. дои : 10.1074/jbc.274.25.17649 . ПМИД 10364203 .
- ^ Розенберг М.Ф., Веларде Г., Форд Р.К., Мартин С., Берридж Г., Керр И.Д., Каллаган Р., Шмидлин А., Вудинг С., Линтон К.Дж., Хиггинс К.Ф. (октябрь 2001 г.). «Переупаковка трансмембранных доменов Р-гликопротеина в ходе транспортного АТФазного цикла» . Журнал ЭМБО . 20 (20): 5615–25. дои : 10.1093/emboj/20.20.5615 . ПМК 125677 . ПМИД 11598005 .
- ^ Перейти обратно: а б Гилсон, Л.; Маханти, Гонконг; Колтер, Р. (декабрь 1990 г.). «Генетический анализ MDR-подобной экспортной системы: секреция колицина V» . Журнал ЭМБО . 9 (12): 3875–3884. дои : 10.1002/j.1460-2075.1990.tb07606.x . ISSN 0261-4189 . ПМК 552155 . ПМИД 2249654 .
- ^ Чхве, Ён Хи; Ю, Ай-Мин (2014). «Переносчики ABC при множественной лекарственной устойчивости и фармакокинетике, а также стратегии разработки лекарств» . Текущий фармацевтический дизайн . 20 (5): 793–807. дои : 10.2174/138161282005140214165212 . ISSN 1381-6128 . ПМК 6341993 . ПМИД 23688078 .
- ^ Макмерри Л., Петруччи Р.Э., Леви С.Б. (июль 1980 г.). «Активный выброс тетрациклина, кодируемый четырьмя генетически различными детерминантами устойчивости к тетрациклину у Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (7): 3974–7. Бибкод : 1980PNAS...77.3974M . дои : 10.1073/pnas.77.7.3974 . ПМК 349750 . ПМИД 7001450 .
- ^ Ри П.А. (2007). «Растительные АТФ-связывающие кассетные транспортеры». Ежегодный обзор биологии растений . 58 : 347–75. doi : 10.1146/annurev.arplant.57.032905.105406 . ПМИД 17263663 .
- ^ Байи А., Ян Х., Мартинойя Э., Гейслер М., Мерфи А.С. (2011). «Уроки завода: изучение функциональности ABCB посредством структурного моделирования» . Границы в науке о растениях . 2 : 108. doi : 10.3389/fpls.2011.00108 . ПМЦ 3355715 . ПМИД 22639627 .
- ^ Гейслер М., Мерфи А.С. (февраль 2006 г.). «Азбука транспорта ауксина: роль p-гликопротеинов в развитии растений». Письма ФЭБС . 580 (4): 1094–102. Бибкод : 2006FEBSL.580.1094G . дои : 10.1016/j.febslet.2005.11.054 . ПМИД 16359667 . S2CID 23368914 .
- ^ Перейти обратно: а б с д Ян Х, Мерфи А.С. (июль 2009 г.). «Функциональное выражение и характеристика переносчиков ауксина Arabidopsis ABCB, AUX 1 и PIN в Schizosaccharomyces pombe» . Заводской журнал . 59 (1): 179–91. дои : 10.1111/j.1365-313X.2009.03856.x . ПМИД 19309458 .
- ^ Блейксли Дж. Дж., Пер В.А., Мерфи А.С. (октябрь 2005 г.). «Ауксин транспорт». Современное мнение в области биологии растений . 8 (5): 494–500. Бибкод : 2005COPB....8..494B . дои : 10.1016/j.pbi.2005.07.014 . ПМИД 16054428 .
- ^ Кречмар Т., Бурла Б., Ли Ю., Мартинойя Э., Надь Р. (сентябрь 2011 г.). «Функции транспортеров ABC в растениях» (PDF) . Очерки по биохимии . 50 (1): 145–60. дои : 10.1042/bse0500145 . ПМИД 21967056 .
- ^ Кубеш М., Ян Х., Рихтер Г.Л., Ченг Ю., Млодзинска Е., Ван Х., Блейксли Дж.Дж., Карраро Н., Петрашек Дж., Зажималова Е., Хойерова К., Пер В.А., Мерфи А.С. (февраль 2012 г.). «Зависимый от концентрации транспортер притока/оттока арабидопсиса ABCB4 регулирует клеточные уровни ауксина в корневом эпидермисе». Заводской журнал . 69 (4): 640–54. дои : 10.1111/j.1365-313X.2011.04818.x . ПМИД 21992190 .
- ^ Доусон Р.Дж., Лочер К.П. (март 2007 г.). «Структура мультилекарственного переносчика ABC Sav1866 из Staphylococcus aureus в комплексе с AMP-PNP» . Письма ФЭБС . 581 (5): 935–8. Бибкод : 2007FEBSL.581..935D . дои : 10.1016/j.febslet.2007.01.073 . ПМИД 17303126 . S2CID 19960736 .
- ^ Веламаканни С., Яо Ю., Гутманн Д.А., ван Вин Х.В. (сентябрь 2008 г.). «Множественный транспорт ABC-транспортером Sav1866 от Staphylococcus aureus». Биохимия . 47 (35): 9300–8. дои : 10.1021/bi8006737 . ПМИД 18690712 .
- ^ Рейтер Г., Джанвилисри Т., Вентер Х., Шахи С., Балакришнан Л., ван Вин Х.В. (сентябрь 2003 г.). «АТФ-связывающая кассета мультилекарственного переносчика LmrA и липидный переносчик MsbA имеют перекрывающуюся субстратную специфичность» . Журнал биологической химии . 278 (37): 35193–8. дои : 10.1074/jbc.M306226200 . ПМИД 12842882 .
- ^ Раец Ч.Р., Рейнольдс К.М., Трент М.С., Бишоп Р.Э. (2007). «Системы модификации липида А у грамотрицательных бактерий» . Ежегодный обзор биохимии . 76 : 295–329. doi : 10.1146/annurev.biochem.76.010307.145803 . ПМК 2569861 . ПМИД 17362200 .
- ^ Перейти обратно: а б Чанг Дж., Рот CB (сентябрь 2001 г.). «Структура MsbA из E. coli: гомолог переносчиков АТФ-связывающей кассеты (ABC) с множественной лекарственной устойчивостью». Наука . 293 (5536): 1793–800. Бибкод : 2001Sci...293.1793C . дои : 10.1126/science.293.5536.1793 . ПМИД 11546864 . (Отозвано, см. дои : 10.1126/science.314.5807.1875b , ПМИД 17185584 ) (Отозвано, см. дои : 10.1126/science.314.5807.1875b )
- ^ Перейти обратно: а б Рейес CL, Чанг Дж. (май 2005 г.). «Структура транспортера ABC MsbA в комплексе с ADP.ванадатом и липополисахаридом». Наука . 308 (5724): 1028–31. Бибкод : 2005Sci...308.1028R . дои : 10.1126/science.1107733 . ПМИД 15890884 . S2CID 37250061 . (Отозвано, см. дои : 10.1126/science.314.5807.1875b , ПМИД 17185584 ) (Отозвано, см. дои : 10.1126/science.314.5807.1875b )
- ^ Бучаклян А.Х., Функ А.Л., Клуг К.С. (июль 2004 г.). «Конформация гомодимера MsbA в состоянии покоя, изученная с помощью сайт-направленного спинового мечения». Биохимия . 43 (26): 8600–6. дои : 10.1021/bi0497751 . ПМИД 15222771 .
- ^ Перейти обратно: а б с Донг Дж., Ян Дж., МакХаураб Х.С. (май 2005 г.). «Структурные основы преобразования энергии в транспортном цикле МсбА». Наука . 308 (5724): 1023–8. Бибкод : 2005Sci...308.1023D . дои : 10.1126/science.1106592 . ПМИД 15890883 . S2CID 1308350 .
- ^ Борбат П.П., Сурендран К., Бортолус М., Зу П., Фрид Дж.Х., Мчаураб Х.С. (октябрь 2007 г.). «Конформационное движение транспортера ABC MsbA, индуцированное гидролизом АТФ» . ПЛОС Биология . 5 (10): е271. doi : 10.1371/journal.pbio.0050271 . ПМК 2001213 . ПМИД 17927448 .
- ^ Чжан, Ге; Байдин, Вадим; Пахил, Каранбир С.; Мойсон, Эйлин; Томасек, Дэвид; Рамадосс, Нитья С.; Чаттерджи, Арнаб К.; Макнамара, Кейс В.; Янг, Трэвис С.; Шульц, Питер Г.; Мередит, Тимоти С.; Кане, Дэниел (7 мая 2018 г.). «Клеточный скрининг для обнаружения ингибиторов биогенеза липополисахаридов» . Труды Национальной академии наук . 115 (26): 6834–6839. Бибкод : 2018PNAS..115.6834Z . дои : 10.1073/pnas.1804670115 . ПМК 6042065 . ПМИД 29735709 .
- ^ Хо, Хоангдун; Миу, Ань; Александр, Мэри Кейт; Гарсия, Натали К.; О, Анжела; Зильберлейб, Инна; Райхельт, Майк; Остин, Кэри Д.; Тэм, Кристина; Шрайвер, Стефани; Ху, Хуэйонг; Лабади, Шарада С.; Лян, Цзюнь; Ван, Лан; Ван, Цзянь; Лу, Ян; Пурки, Ханс Э.; Куинн, Джон; Франке, Ивонн; Кларк, Кевин; Березини, Морин Х.; Тан, Ман-Ва; Селлерс, Бенджамин Д.; Маурер, Тилль; Келер, Майкл Ф.Т.; Векслер, Аарон Т.; Кифер, Джеймс Р.; Верма, Вишал; Сюй, Имин; Нисияма, Мирей; Паянде, Цзян; Кот, Кристофер М. (май 2018 г.). «Структурная основа двухрежимного ингибирования транспортера ABC MsbA». Природа . 557 (7704): 196–201. Бибкод : 2018Natur.557..196H . дои : 10.1038/s41586-018-0083-5 . ПМИД 29720648 . S2CID 13660653 .
- ^ Гутманн Д.А., Уорд А., Урбач И.Л., Чанг Г., ван Вин Х.В. (январь 2010 г.). «Понимание полиспецифичности мультилекарственных переносчиков ABC: устранение пробелов в ABCB1» . Тенденции биохимических наук . 35 (1): 36–42. дои : 10.1016/j.tibs.2009.07.009 . ПМК 4608440 . ПМИД 19819701 .
- ^ Леонард Г.Д., Фойо Т., Бейтс С.Е. (2003). «Роль ABC-транспортеров в клинической практике» . Онколог . 8 (5): 411–24. doi : 10.1634/теонколог.8-5-411 . ПМИД 14530494 . S2CID 2780630 .
- ^ Лаге Л (2009). «Транспортеры ABC как мишень для устранения множественной лекарственной устойчивости, опосредованного интерференцией РНК». ABC-транспортеры в микроорганизмах . Кайстер Академик. ISBN 978-1-904455-49-3 .
- ^ Аннаерт П.П., Тернклифф Р.З., Бут К.Л., Таккер Д.Р., Брауэр К.Л. (октябрь 2001 г.). «P-гликопротеин-опосредованная экскреция желчи in vitro в гепатоцитах крысы, культивированных в сэндвич-культуре». Препарат Метаб Диспос . 29 (10): 1277–83. ПМИД 11560870 .
- ^ Перейти обратно: а б Аннаерт П.П., Брауэр К.Л. (март 2005 г.). «Оценка взаимодействия лекарств при гепатобилиарном транспорте с использованием родамина 123 в гепатоцитах крысы, культивированных в сэндвич-культивировании». Препарат Метаб Диспос . 33 (3): 388–94. дои : 10.1124/dmd.104.001669 . ПМИД 15608134 . S2CID 7063502 .
- ^ Матссон, Пяр (2007). «АТФ-связывающие транспортеры оттока кассет и пассивная проницаемость мембран при абсорбции и распределении лекарств» . Дива .
- ^ Главинас Х., Крайчи П., Черепеш Дж., Саркади Б. (январь 2004 г.). «Роль транспортеров ABC в лекарственной устойчивости, метаболизме и токсичности». Текущая доставка лекарств . 1 (1): 27–42. дои : 10.2174/1567201043480036 . ПМИД 16305368 .
- ^ Главанас Х., Мен Д., Яни М., Остерхейс Б., Эреди-Сабо К., Крайчи П. (июнь 2008 г.). «Использование препаратов мембранных везикул для изучения взаимодействия лекарственного средства и переносчика ABC». Экспертное заключение по метаболизму и токсикологии лекарственных средств . 4 (6): 721–32. дои : 10.1517/17425255.4.6.721 . ПМИД 18611113 . S2CID 86198612 .
- ^ Весь этот том посвящен различным используемым методам: Никайдо Х., Холл Дж. (1998). Транспортеры ABC: биохимические, клеточные и молекулярные аспекты . Методы энзимологии. Том. 292. стр. 3–853. дои : 10.1016/s0076-6879(00)x0188-7 . ISBN 9780121821937 . ПМИД 9711542 .
- ^ Хорио М., Готтесман М.М., Пастан I (май 1988 г.). «АТФ-зависимый транспорт винбластина в везикулах из клеток человека с множественной лекарственной устойчивостью» . Труды Национальной академии наук Соединенных Штатов Америки . 85 (10): 3580–4. Бибкод : 1988PNAS...85.3580H . дои : 10.1073/pnas.85.10.3580 . ПМК 280257 . ПМИД 3368466 .
- ^ Василиу, В; Василиу, К; Неберт, Д.В. (апрель 2009 г.). «Семейство транспортеров АТФ-связывающих кассет (ABC) человека» . Геномика человека . 3 (3): 281–90. дои : 10.1186/1479-7364-3-3-281 . ПМК 2752038 . ПМИД 19403462 .
- ^ Перейти обратно: а б Чен З.С., Тивари А.К. (сентябрь 2011 г.). «Белки множественной лекарственной устойчивости (MRP/ABCC) при химиотерапии рака и генетических заболеваниях» . Журнал ФЭБС . 278 (18): 3226–45. дои : 10.1111/j.1742-4658.2011.08235.x . ПМК 3168698 . ПМИД 21740521 .
- ^ Сайер М.Х. (июнь 2000 г.). «Функционально-филогенетическая система классификации трансмембранных переносчиков растворенных веществ» . Обзоры микробиологии и молекулярной биологии . 64 (2): 354–411. дои : 10.1128/MMBR.64.2.354-411.2000 . ПМК 98997 . ПМИД 10839820 . ; Группа биоинформатики Saier Lab. «3.A.1 Суперсемейство АТФ-связывающих кассет (ABC)» . База данных классификации транспортеров (TCDB) . Калифорнийский университет в Сан-Диего.
- ^ Ходжа М., Ма К., Сайер М.Х. (март 2005 г.). «Топологический анализ интегральных мембранных компонентов прокариотических систем оттока ABC» . Исследования в области микробиологии . 156 (2): 270–7. дои : 10.1016/j.resmic.2004.07.010 . ПМИД 15748994 .
- ^ Чжэн, WH; Вестермарк, Å; Шлыков М.А.; Редди, В.; Вс, Э.И.; Сайер М.Х.-младший (6 мая 2013 г.). «Эволюционные взаимоотношения переносчиков поглощения АТФ-связывающей кассеты (ABC)» . БМК Микробиология . 13:98 . дои : 10.1186/1471-2180-13-98 . ПМЦ 3654945 . ПМИД 23647830 .
Дальнейшее чтение
[ редактировать ]- Сентпетери З, Керн А, Лилиом К, Саркади Б, Варади А, Бакос Э (октябрь 2004 г.). «Роль консервативных глицинов сигнатурных мотивов АТФ-связывающей кассеты MRP1 в связи между сайтом связывания субстрата и каталитическими центрами» . Журнал биологической химии . 279 (40): 41670–8. дои : 10.1074/jbc.M406484200 . ПМИД 15252017 .
- Фицджеральд М.Л., Окухира К., Шорт Г.Ф., Мэннинг Дж.Дж., Белл С.А., Фриман М.В. (ноябрь 2004 г.). «АТФ-связывающий кассетный транспортер А1 содержит новый С-концевой мотив VFVNFA, который необходим для оттока холестерина и активности связывания ApoA-I» . Журнал биологической химии . 279 (46): 48477–85. дои : 10.1074/jbc.M409848200 . ПМИД 15347662 .
- Линтон К.Дж. (2011). ABC-транспортеры физиологии и болезней человека: генетика и биохимия АТФ-связывающей кассеты . Всемирная научная. ISBN 978-981-4280-06-8 . Архивировано из оригинала 11 мая 2012 г. Проверено 16 мая 2012 г.
Внешние ссылки
[ редактировать ]- Классификация транспортеров ABC в TCDB
- ABCdb База данных архейных и бактериальных систем ABC, ABCdb
- АТФ-связывание + кассета + транспортеры в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
| Базы данных органов управления : Национальные |
|---|