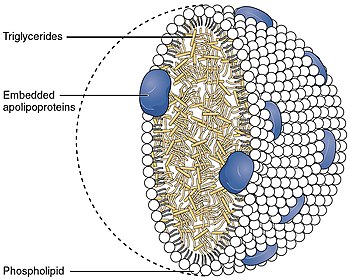
Липид

Липиды представляют собой обширную группу органических соединений, в которую входят жиры , воски , стерины , жирорастворимые витамины (такие как витамины А , D , Е и К ), моноглицериды , диглицериды , фосфолипиды и другие. Функции липидов включают хранение энергии, передачу сигналов и действие в качестве структурных компонентов клеточных мембран . [3] [4] Липиды находят применение в косметической и пищевой промышленности , а также в нанотехнологиях . [5]
Липиды можно в широком смысле определить как гидрофобные или амфифильные небольшие молекулы; Амфифильная природа некоторых липидов позволяет им образовывать такие структуры, как везикулы , многослойные/ однослойные липосомы или мембраны в водной среде. Биологические липиды полностью или частично происходят из двух различных типов биохимических субъединиц или «строительных блоков»: кетоацильной и изопреновой групп. [3] Используя этот подход, липиды можно разделить на восемь категорий: жирные ацилы , глицеролипиды , глицерофосфолипиды , сфинголипиды , сахаролипиды и поликетиды (полученные в результате конденсации кетоацильных субъединиц); и стероловые липиды и пренольные липиды (полученные в результате конденсации изопреновых субъединиц). [3]
Хотя термин «липид» иногда используется как синоним жиров, жиры представляют собой подгруппу липидов, называемую триглицеридами . Липиды также включают такие молекулы, как жирные кислоты и их производные (включая три-, ди-, моноглицериды и фосфолипиды), а также другие стеролсодержащие , метаболиты такие как холестерин . [6] Хотя люди и другие млекопитающие используют различные пути биосинтеза как для расщепления, так и для синтеза липидов, некоторые незаменимые липиды не могут быть получены таким путем и должны быть получены с пищей.
История [ править ]
В 1815 году Анри Браконно разделил липиды ( graisses ) на две категории: suifs (твердые жиры или жир) и huiles (жидкие масла). [7] В 1823 году Мишель Эжен Шеврель разработал более подробную классификацию, включающую масла, жиры, жир, воски, смолы, бальзамы и летучие масла (или эфирные масла). [8] [9] [10]
О первом синтетическом триглицериде сообщил Теофиль-Жюль Пелуз году, когда он получил трибутирин путем обработки масляной кислоты глицерином в 1844 в присутствии концентрированной серной кислоты . [11] Несколько лет спустя Марселлен Бертло , один из учеников Пелуза, синтезировал тристеарин и трипальмитин путем реакции аналогичных жирных кислот с глицерином в присутствии газообразного хлористого водорода при высокой температуре. [12]
В 1827 году Уильям Праут признал жир («маслянистые» пищевые вещества), а также белки («белковые») и углеводы («сахарин») важным питательным веществом для людей и животных. [13] [14]
В течение столетия химики рассматривали «жиры» как простые липиды, состоящие из жирных кислот и глицерина (глицеридов), но позже были описаны новые формы. Теодор Гобли (1847) обнаружил в мозге млекопитающих и куриных яйцах фосфолипиды, названные им « лецитинами ». Тудичум обнаружил в мозге человека некоторые фосфолипиды ( цефалин ), гликолипиды ( цереброзид ) и сфинголипиды ( сфингомиелин ). [9]
Термины «липоид», «липин», «липид» и «липид» использовались в разных значениях от автора к автору. [15] В 1912 году Розенблюм и Гис предложили замену «липоида» на «липин». [16] В 1920 году Блур ввел новую классификацию «липоидов»: простые липоиды (жиры и воски), сложные липоиды (фосфолипоиды и гликолипоиды) и производные липоиды (жирные кислоты, спирты , стерины). [17] [18]
Слово липид , этимологически происходящее от греческого λίπος, Lipos «жир», было введено в 1923 году французским фармакологом Габриэлем Бертраном . [19] Бертран включил в понятие не только традиционные жиры (глицериды), но и «липоиды», имеющие сложное строение. [9] Слово «липид» было единогласно одобрено международной комиссией Société de Chimie Biologique на пленарном заседании 3 июля 1923 года. слово «липид» Позже было переведено на английский язык как «липид» из-за его произношения ('lɪpɪd). Во французском языке суффикс -ide от древнегреческого -ίδης (означающий «сын» или «потомок») всегда произносится (ɪd).
В 1947 году Т. П. Хилдитч определил «простые липиды» как жиры и воски (настоящие воски, стерины, спирты). [20] [ нужна страница ]
Категории [ править ]
классифицировал липиды на восемь категорий. Lipid MAPS Консорциум [3] следующее:
Жирные ацилы [ править ]


Жирные ацилы, общий термин для описания жирных кислот, их конъюгатов и производных, представляют собой разнообразную группу молекул, синтезируемых путем удлинения цепи праймера ацетил-КоА с группами малонил-КоА или метилмалонил-КоА в процессе, называемом синтезом жирных кислот . [21] [22] Они состоят из углеводородной цепи , оканчивающейся группой карбоновой кислоты ; такое расположение придает молекуле полярный гидрофильный конец конец и неполярный гидрофобный , нерастворимый в воде. Структура жирных кислот является одной из наиболее фундаментальных категорий биологических липидов и обычно используется в качестве строительного блока более структурно сложных липидов. Углеродная цепь, обычно имеющая длину от четырех до 24 атомов углерода, [23] может быть насыщенным или ненасыщенным и может быть присоединен к функциональным группам, содержащим кислород , галогены , азот и серу . Если жирная кислота содержит двойную связь, существует возможность цис- или транс - геометрической изомерии молекулы , что существенно влияет на конфигурацию . Цис -двойные связи заставляют цепь жирной кислоты изгибаться, и этот эффект усугубляется увеличением количества двойных связей в цепи. Три двойные связи в 18-углеродной линоленовой кислоте , наиболее распространенной жирно-ацильной цепи мембран тилакоидов растений , придают этим мембранам высокую текучесть, несмотря на низкие температуры окружающей среды. [24] а также заставляет линоленовую кислоту давать доминирующие острые пики в спектрах 13-С ЯМР высокого разрешения хлоропластов. Это, в свою очередь, играет важную роль в структуре и функции клеточных мембран. [25] : 193–5 Большинство встречающихся в природе жирных кислот имеют цис -конфигурацию, хотя транс- форма существует в некоторых природных и частично гидрогенизированных жирах и маслах. [26]
Примеры биологически важных жирных кислот включают эйкозаноиды , полученные главным образом из арахидоновой кислоты и эйкозапентаеновой кислоты , которые включают простагландины , лейкотриены и тромбоксаны . Докозагексаеновая кислота также важна в биологических системах, особенно в отношении зрения. [27] [28] Другими основными классами липидов в категории жирных кислот являются жирные эфиры и жирные амиды. Сложные эфиры жирных кислот включают важные биохимические промежуточные соединения, такие как сложные эфиры воска тиоэфиров жирных кислот , производные кофермента А , производные тиоэфиров жирных кислот ACP и карнитины жирных кислот. Жирные амиды включают N-ацилэтаноламины , такие как каннабиноидный нейромедиатор анандамид . [29]
Глицеролипиды [ править ]

Глицеролипиды состоят из моно-, ди- и тризамещенных глицеринов . [30] наиболее известными из них являются триэфиры жирных кислот глицерина, называемые триглицеридами . Слово «триацилглицерин» иногда используется как синоним слова «триглицерид». В этих соединениях каждая из трех гидроксильных групп глицерина этерифицирована, как правило, разными жирными кислотами. Поскольку они функционируют как хранилище энергии, эти липиды составляют основную часть запасаемого жира в тканях животных. Гидролиз сложноэфирных связей триглицеридов и высвобождение глицерина и жирных кислот из жировой ткани являются начальными этапами метаболизма жиров. [31] : 630–1
Дополнительные подклассы глицеролипидов представлены гликозилглицеринами, для которых характерно наличие одного или нескольких остатков сахара , присоединенных к глицерину посредством гликозидной связи . Примерами структур этой категории являются дигалактозилдиацилглицерины, обнаруженные в растительных мембранах. [32] млекопитающих и семинолипид из сперматозоидов . [33]
Глицерофосфолипиды [ править ]
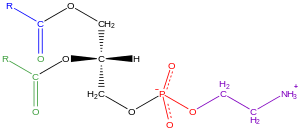
Глицерофосфолипиды, обычно называемые фосфолипидами (хотя сфингомиелины также классифицируются как фосфолипиды), широко распространены в природе и являются ключевыми компонентами липидного бислоя клеток. [34] а также участвует в метаболизме и передаче сигналов клеткам . [35] Нервная ткань (включая мозг) содержит относительно большое количество глицерофосфолипидов, и изменения в их составе связаны с различными неврологическими расстройствами. [36] Глицерофосфолипиды можно подразделить на отдельные классы в зависимости от природы полярной головной группы в положении sn -3 глицеринового остова у эукариот и эубактерий или в положении sn -1 в случае архебактерий . [37]
Примерами глицерофосфолипидов, обнаруженных в биологических мембранах, являются фосфатидилхолин (также известный как PC, GPCho или лецитин ), фосфатидилэтаноламин (PE или GPEtn) и фосфатидилсерин (PS или GPSer). Помимо того, что некоторые глицерофосфолипиды в эукариотических клетках служат основным компонентом клеточных мембран и сайтами связывания внутри- и межклеточных белков, такие как фосфатидилинозитолы и фосфатидные кислоты, они являются либо предшественниками, либо самими мембранными вторичными мессенджерами . [31] : 844 Обычно одна или обе эти гидроксильные группы ацилируются длинноцепочечными жирными кислотами, но у архебактерий существуют также алкилсвязанные и 1Z-алкенилсвязанные ( плазмалоген ) глицерофосфолипиды, а также варианты диалкилэфиров. [38]
Сфинголипиды [ править ]
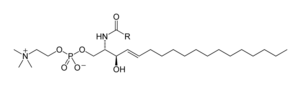
Сфинголипиды представляют собой сложное семейство соединений. [39] которые имеют общую структурную особенность: основную цепь сфингоидного основания , которая синтезируется de novo из аминокислоты серина и длинноцепочечного жирного ацил-КоА, а затем превращается в церамиды , фосфосфинголипиды, гликосфинголипиды и другие соединения. Основное основание сфингоида млекопитающих обычно называют сфингозином . Церамиды (N-ацил-сфингоидные основания) представляют собой основной подкласс производных сфингоидных оснований с жирной кислотой, связанной с амидом . Жирные кислоты обычно являются насыщенными или мононенасыщенными с длиной цепи от 16 до 26 атомов углерода. [25] : 421–2
Основными фосфосфинголипидами млекопитающих являются сфингомиелины (церамиды фосфохолины), [40] тогда как насекомые содержат в основном церамиды фосфоэтаноламины [41] а грибы имеют фитоцерамидные фосфоинозитолы и маннозосодержащие головные группы. [42] Гликосфинголипиды представляют собой разнообразное семейство молекул, состоящих из одного или нескольких остатков сахара, связанных гликозидной связью со сфингоидным основанием. Примерами являются простые и сложные гликосфинголипиды, такие как цереброзиды и ганглиозиды .
Стеролы [ править ]

Стеролы, такие как холестерин и его производные, являются важным компонентом мембранных липидов. [43] наряду с глицерофосфолипидами и сфингомиелинами. Другими примерами стеринов являются желчные кислоты и их конъюгаты. [44] которые у млекопитающих являются окисленными производными холестерина и синтезируются в печени. Растительными эквивалентами являются фитостерины , такие как β-ситостерин , стигмастерин и брассикастерин ; последнее соединение также используется в качестве биомаркера роста водорослей . [45] Преобладающим стеролом в грибов клеточных мембранах является эргостерин . [46]
Стерины – это стероиды , у которых один из атомов водорода замещен гидроксильной группой в положении 3 углеродной цепи. Они имеют общую со стероидами структуру ядра из четырех колец. Стероиды выполняют различные биологические роли в качестве гормонов и сигнальных молекул . Восемнадцатиуглеродные стероиды (C18) включают семейство эстрогенов , тогда как стероиды C19 включают андрогены, такие как тестостерон и андростерон . Подкласс C21 включает прогестагены , а также глюкокортикоиды и минералокортикоиды . [2] : 749 Секостероиды витамина , содержащие различные формы D , характеризуются расщеплением кольца B основной структуры. [47]
Пренолы [ править ]

Липиды пренола синтезируются из пятиуглеродных предшественников изопентенилдифосфата и диметилаллилдифосфата , которые производятся главным образом по пути мевалоновой кислоты (MVA). [48] Простые изопреноиды (линейные спирты, дифосфаты и т. д.) образуются путем последовательного присоединения звеньев С5 и классифицируются по числу этих терпеновых звеньев. Структуры, содержащие более 40 атомов углерода, известны как политерпены. Каротиноиды — это важные простые изопреноиды, которые действуют как и предшественники витамина А. антиоксиданты [49] Другим биологически важным классом молекул являются хиноны и гидрохиноны , которые содержат изопреноидный хвост, прикрепленный к хиноноидному ядру неизопреноидного происхождения. [50] Витамин Е и витамин К , а также убихиноны являются примерами этого класса. Прокариоты синтезируют полипренолы (называемые бактопренолами ), у которых концевая изопреноидная единица, присоединенная к кислороду, остается ненасыщенной, тогда как у животных полипренолов ( долихолов ) терминальная изопреноидная единица восстанавливается. [51]
Сахаролипиды [ править ]

Сахаролипиды представляют собой соединения, в которых жирные кислоты связаны с основной цепью сахара, образуя структуры, совместимые с бислоями мембран. В сахаролипидах моносахарид заменяет глицериновую основу, присутствующую в глицеролипидах и глицерофосфолипидах. Наиболее известными сахаролипидами являются ацилированные глюкозамина предшественники липида А, входящего в состав липополисахаридов грамотрицательных бактерий . Типичные молекулы липида А представляют собой дисахариды глюкозамина, которые содержат до семи жирно-ацильных цепей. Минимальным липополисахаридом, необходимым для роста E. coli, является Kdo 2 -Липид А, гексаацилированный дисахарид глюкозамина, гликозилированный двумя остатками 3-дезокси-D-маннооктулозоновой кислоты (Kdo). [52]
Поликетиды [ править ]
Поликетиды синтезируются путем полимеризации ацетильных и пропионильных субъединиц с помощью классических ферментов, а также итерационных и мультимодульных ферментов, которые имеют общие механические характеристики с синтазами жирных кислот . Они включают множество вторичных метаболитов и натуральных продуктов животного, растительного, бактериального, грибкового и морского происхождения и имеют большое структурное разнообразие. [53] [54] Многие поликетиды представляют собой циклические молекулы, чьи основные цепи часто модифицируются гликозилированием , метилированием , гидроксилированием , окислением или другими процессами. Многие широко используемые противомикробные , противопаразитарные и противораковые средства представляют собой поликетиды или производные поликетидов, такие как эритромицины , тетрациклины , авермектины и противоопухолевые эпотилоны . [55]
Биологические функции [ править ]
Компонент биологических мембран [ править ]
Эукариотические клетки имеют разделенные на мембраны органеллы , которые выполняют различные биологические функции. Глицерофосфолипиды , являются основным структурным компонентом биологических мембран , как клеточной плазматической мембраны так и внутриклеточных мембран органелл; в клетках животных плазматическая мембрана физически отделяет внутриклеточные компоненты от внеклеточной среды. [ нужна ссылка ] Глицерофосфолипиды представляют собой амфипатические молекулы (содержащие как гидрофобные, так и гидрофильные области), которые содержат глицериновое ядро, связанное с двумя «хвостами» жирных кислот сложноэфирными связями и с одной «головной» группой сложноэфирной связью фосфата . [ нужна ссылка ] другие неглицеридные липидные компоненты, такие как сфингомиелин и стерины (в основном холестерин в мембранах клеток животных). Хотя глицерофосфолипиды являются основным компонентом биологических мембран, в биологических мембранах также обнаруживаются [56] [2] : 329–331 В растениях и водорослях галактозилдиацилглицерины [57] и сульфохиновозилдиацилглицерин, [32] лишенные фосфатной группы, являются важными компонентами мембран хлоропластов и родственных им органелл и входят в число наиболее распространенных липидов в фотосинтезирующих тканях, в том числе высших растений, водорослей и некоторых бактерий. [58]
Мембраны тилакоидов растений содержат самый большой липидный компонент небислойного моногалактозилдиглицерида (МГДГ) и небольшое количество фосфолипидов; Несмотря на этот уникальный липидный состав, было показано, что мембраны тилакоидов хлоропластов содержат динамический липидный бислойный матрикс, как показали исследования с помощью магнитного резонанса и электронного микроскопа. [59]
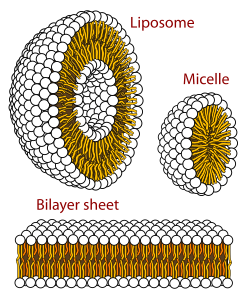
Биологическая мембрана представляет собой форму ламеллярного липидного бислоя . Образование липидных бислоев является энергетически предпочтительным процессом, когда описанные выше глицерофосфолипиды находятся в водной среде. [2] : 333–4 Это известно как гидрофобный эффект . В водной системе полярные головки липидов ориентируются по направлению к полярной водной среде, тогда как гидрофобные хвосты сводят к минимуму их контакт с водой и имеют тенденцию группироваться вместе, образуя везикулу ; в зависимости от концентрации липида это биофизическое взаимодействие может привести к образованию мицелл , липосом или липидных бислоев . Другие агрегации также наблюдаются и являются частью полиморфизма амфифильного (липидного) поведения. Фазовое поведение является областью изучения биофизики . [60] [61] Мицеллы и бислои образуются в полярной среде в результате процесса, известного как гидрофобный эффект. [62] При растворении липофильного или амфифильного вещества в полярном окружении полярные молекулы (т. е. вода в водном растворе) становятся более упорядоченными вокруг растворенного липофильного вещества, поскольку полярные молекулы не могут образовывать водородные связи с липофильными участками амфифила. Таким образом, в водной среде молекулы воды образуют упорядоченную « клатратную » клетку вокруг растворенной липофильной молекулы. [63]
Образование липидов в мембранах протоклеток представляет собой ключевой этап в моделях абиогенеза , происхождения жизни. [64]
Хранение энергии [ править ]
Триглицериды, хранящиеся в жировой ткани, являются основной формой хранения энергии как у животных, так и у растений. Они являются основным источником энергии при аэробном дыхании. При полном окислении жирных кислот выделяется около 38 кДж/г (9 ккал/г ) по сравнению с всего лишь 17 кДж/г (4 ккал/г) при окислительном расщеплении углеводов и белков . Адипоциты , или жировые клетки, предназначены для непрерывного синтеза и расщепления триглицеридов у животных, причем разрушение контролируется главным образом активацией гормоночувствительного фермента липазы . [65] Перелетные птицы, которым приходится летать на большие расстояния без еды, используют триглицериды в качестве топлива для своих полетов. [2] : 619
Сигнализация [ править ]
Появились данные, показывающие, что передача сигналов липидов является жизненно важной частью передачи сигналов в клетках . [66] [67] [68] [69] Передача сигналов липидов может происходить посредством активации связанных с G-белком или ядерных рецепторов , а представители нескольких различных категорий липидов были идентифицированы как сигнальные молекулы и клеточные мессенджеры . [70] К ним относятся сфингозин-1-фосфат , сфинголипид, полученный из церамида, который является мощной молекулой-мессенджером, участвующим в регуляции мобилизации кальция. [71] рост клеток и апоптоз; [72] диацилглицерин и фосфатидилинозитолфосфаты (PIP), участвующие в кальций-опосредованной активации протеинкиназы C ; [73] простагландины , которые представляют собой один из типов эйкозаноидов , полученных из жирных кислот, участвующих в воспалении и иммунитете ; [74] стероидные гормоны, такие как эстроген , тестостерон и кортизол , которые модулируют множество функций, таких как репродуктивная функция, обмен веществ и кровяное давление; и оксистерины, такие как 25-гидроксихолестерин, которые являются X-рецепторов печени агонистами . [75] Известно, что фосфатидилсериновые липиды участвуют в передаче сигналов фагоцитоза апоптотических клеток или фрагментов клеток. Они достигают этого, подвергаясь воздействию внеклеточной поверхности клеточной мембраны после инактивации флипаз , которые размещают их исключительно на цитозольной стороне, и активации скрамблаз, которые меняют ориентацию фосфолипидов. После того, как это происходит, другие клетки распознают фосфатидилсерины и фагоцитируют клетки или фрагменты клеток, подвергающие их воздействию. [76]
Другие функции [ править ]
«Жирорастворимые» витамины ( A , D , E и K ), которые представляют собой липиды на основе изопрена , являются важными питательными веществами, хранящимися в печени и жировых тканях, с разнообразным спектром функций. Ацилкарнитины участвуют в транспортировке и метаболизме жирных кислот в митохондрии и из них, где они подвергаются бета-окислению . [77] Полипренолы и их фосфорилированные производные также играют важную транспортную роль, в данном случае транспорта олигосахаридов через мембраны. Полипренолфосфатные сахара и полипренолдифосфатные сахара участвуют в реакциях внецитоплазматического гликозилирования, во внеклеточном биосинтезе полисахаридов (например, полимеризация пептидогликана у бактерий) и в N- гликозилировании эукариотических белков . [78] [79] Кардиолипины представляют собой подкласс глицерофосфолипидов, содержащий четыре ацильные цепи и три глицериновые группы, которые особенно распространены во внутренней мембране митохондрий. [80] [81] Считается, что они активируют ферменты, участвующие в окислительном фосфорилировании . [82] Липиды также составляют основу стероидных гормонов. [83]
Метаболизм [ править ]
Основными диетическими липидами для человека и других животных являются триглицериды животного и растительного происхождения, стерины и мембранные фосфолипиды. В процессе липидного обмена синтезируются и разрушаются липидные запасы и образуются структурные и функциональные липиды, характерные для отдельных тканей.
Биосинтез [ править ]
У животных при избытке пищевых углеводов избыток углеводов превращается в триглицериды. Это включает синтез жирных кислот из ацетил-КоА и этерификацию жирных кислот с образованием триглицеридов — процесс, называемый липогенезом . [2] : 634 Жирные кислоты производятся синтазами жирных кислот , которые полимеризуются, а затем восстанавливают звенья ацетил-КоА. Ацильные цепи в жирных кислотах удлиняются за счет цикла реакций, в которых ацетильная группа присоединяется, восстанавливается до спирта, дегидратируется до алкеновой группы, а затем снова восстанавливается до алкановой группы. Ферменты биосинтеза жирных кислот делятся на две группы: у животных и грибов все эти реакции синтаз жирных кислот осуществляются одним многофункциональным белком, [84] растений в то время как в пластидах и бактериях каждый этап этого пути выполняют отдельные ферменты. [85] [86] Жирные кислоты могут впоследствии превращаться в триглицериды, которые упаковываются в липопротеины и секретируются печенью.
Синтез ненасыщенных жирных кислот включает реакцию десатурации , при которой в жирную ацильную цепь вводится двойная связь. Например, у людей десатурация стеариновой кислоты стеароил -КоА-десатуразой-1 приводит к образованию олеиновой кислоты . Дважды ненасыщенная жирная кислота, линолевая кислота , а также трижды ненасыщенная α-линоленовая кислота не могут синтезироваться в тканях млекопитающих и, следовательно, являются незаменимыми жирными кислотами и должны поступать с пищей. [2] : 643
Синтез триглицеридов происходит в эндоплазматическом ретикулуме метаболическими путями, при которых ацильные группы жирных ацил-КоА передаются на гидроксильные группы глицерин-3-фосфата и диацилглицерина. [2] : 733–9
Терпены и изопреноиды , включая каротиноиды , производятся путем сборки и модификации изопреновых единиц, полученных из реакционноспособных предшественников изопентенилпирофосфата и диметилаллилпирофосфата . [48] Эти предшественники могут быть получены разными способами. У животных и архей мевалонатный путь производит эти соединения из ацетил-КоА. [87] в то время как у растений и бактерий немевалонатный путь использует в качестве субстратов пируват и глицеральдегид-3-фосфат . [48] [88] Одной из важных реакций, в которых используются активированные доноры изопрена, является биосинтез стероидов . Здесь единицы изопрена соединяются вместе, образуя сквален , а затем сворачиваются и формируются в набор колец, образуя ланостерин . [89] Ланостерин затем может быть преобразован в другие стероиды, такие как холестерин и эргостерин. [89] [90]
Деградация [ править ]
Бета-окисление — это метаболический процесс, в ходе которого жирные кислоты расщепляются в митохондриях или пероксисомах с образованием ацетил-КоА . По большей части жирные кислоты окисляются по механизму, который аналогичен, но не идентичен обращению процесса синтеза жирных кислот. То есть двухуглеродные фрагменты последовательно удаляются с карбоксильного конца кислоты после стадий дегидрирования , гидратации и окисления с образованием бета-кетокислоты , которая расщепляется тиолизом . Ацетил-КоА затем в конечном итоге превращается в аденозинтрифосфат (АТФ), CO 2 и H 2 O с использованием цикла лимонной кислоты и цепи переноса электронов . Следовательно, цикл лимонной кислоты может начинаться с ацетил-КоА, когда жир расщепляется для получения энергии, если глюкозы мало или она отсутствует. Энергетический выход полного окисления пальмитата жирной кислоты составляет 106 АТФ. [2] : 625–6 Ненасыщенные жирные кислоты и жирные кислоты с нечетной цепью требуют дополнительных ферментативных этапов для разложения.
Питание и здоровье [ править ]
Большая часть жиров, содержащихся в пище, находится в форме триглицеридов, холестерина и фосфолипидов. Некоторое количество пищевых жиров необходимо для облегчения усвоения жирорастворимых витаминов ( A , D , E и K ) и каротиноидов . [91] : 903 У людей и других млекопитающих есть диетическая потребность в определенных незаменимых жирных кислотах, таких как линолевая кислота ( жирная кислота омега-6 ) и альфа-линоленовая кислота (жирная кислота омега-3), поскольку они не могут быть синтезированы из простых предшественников в рационе. . [2] : 643 Обе эти жирные кислоты представляют собой 18-углеродные полиненасыщенные жирные кислоты, различающиеся числом и положением двойных связей. Большинство растительных масел богаты линолевой кислотой ( сафлоровое , подсолнечное и кукурузное масла). Альфа-линоленовая кислота содержится в зеленых листьях растений, а также в некоторых семенах, орехах и бобовых (в частности , льне , рапсе , грецких орехах и сое ). [92] Рыбий жир особенно богат длинноцепочечными жирными кислотами омега-3, эйкозапентаеновой кислотой и докозагексаеновой кислотой . [91] : 388 Многие исследования показали положительную пользу для здоровья, связанную с потреблением жирных кислот омега-3, в отношении развития детей, рака, сердечно-сосудистых заболеваний и различных психических заболеваний (таких как депрессия, синдром дефицита внимания и гиперактивности и деменция). [93] [94]
Напротив, в настоящее время хорошо известно, что потребление трансжиров , например тех, которые присутствуют в частично гидрогенизированных растительных маслах , является фактором риска сердечно-сосудистых заболеваний . Полезные для человека жиры могут быть превращены в трансжиры при неправильных методах приготовления, что приводит к перевариванию липидов. [95] [96] [97]
Несколько исследований показали, что общее потребление жиров с пищей связано с повышенным риском ожирения. [98] [99] и диабет; [100] Другие, в том числе исследование по изменению диеты в рамках Инициативы по здоровью женщин, восьмилетнее исследование с участием 49 000 женщин, исследование здоровья медсестер и последующее исследование медицинских работников, не выявили таких связей. [101] [102] Ни одно из этих исследований не выявило какой-либо связи между процентом калорий, получаемых из жиров, и риском развития рака, болезней сердца или увеличения веса. Источник питания, [103] Веб-сайт, поддерживаемый отделом питания Школы общественного здравоохранения Т.Ч. Чана при Гарвардском университете , обобщает текущие данные о влиянии пищевых жиров: «Детальные исследования — большая часть которых проведена в Гарварде — показывают, что общее количество жиров в организме диета на самом деле не связана с весом или болезнью». [104]
См. также [ править ]
- Твердые липидные наночастицы – новая система доставки лекарств
- Простой липид
- Тест эмульсии
- Липидный микродомен
- Мембранный липид - молекулы липидов на клеточной мембране.
- Липидомика - крупномасштабное исследование липидного обмена организма с использованием высокотехнологичных методов химического анализа.
- Липидом – совокупность липидов в клетках.
- Белково-липидное взаимодействие
- Фенольные липиды - класс органических соединений, класс натуральных продуктов, состоящих из длинных алифатических цепей и фенольных колец, которые встречаются в растениях, грибах и бактериях.
Ссылки [ править ]
- ^ Мейтленд-младший (1998). Органическая химия . WW Norton & Co Inc (Нью-Йорк). п. 139. ИСБН 978-0-393-97378-5 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж Страйер Л., Берг Дж.М., Тимочко Дж.Л. (2007). Биохимия (6-е изд.). Сан-Франциско: WH Freeman. ISBN 978-0-7167-8724-2 .
- ^ Jump up to: Перейти обратно: а б с д Фэйи Э., Субраманиам С., Мерфи Р.К., Нисидзима М., Раец Ч.Р., Симидзу Т., Спенер Ф., ван Меер Дж., Уэйкелам М.Дж., Деннис Э.А. (апрель 2009 г.). «Обновление комплексной системы классификации липидов LIPID MAPS» . Журнал исследований липидов . 50 (С1): С9–14. doi : 10.1194/jlr.R800095-JLR200 . ПМЦ 2674711 . ПМИД 19098281 .
- ^ Субраманиам С., Фэхи Э., Гупта С., Суд М., Бирнс Р.В., Коттер Д., Динасарапу А.Р., Маурья М.Р. (октябрь 2011 г.). «Биоинформатика и системная биология липидома» . Химические обзоры . 111 (10): 6452–6490. дои : 10.1021/cr200295k . ПМЦ 3383319 . ПМИД 21939287 .
- ^ Машаги С., Джадиди Т., Кендеринк Г. , Машаги А. (февраль 2013 г.). «Липидная нанотехнология» . Международный журнал молекулярных наук . 14 (2): 4242–4282. дои : 10.3390/ijms14024242 . ПМЦ 3588097 . ПМИД 23429269 .

- ^ Мишель А., Хопкинс Дж., Маклафлин К.В., Джонсон С., Уорнер М.К., ЛаХарт Д., Райт Дж.Д. (1993). Биология человека и здоровье . Энглвуд Клиффс, Нью-Джерси: Прентис Холл. ISBN 978-0-13-981176-0 .
- ^ Браконнот Х. (31 марта 1815 г.). «О природе жирных веществ» . Анналы химии . 2 (XCIII): 225–277.
- ^ Шеврель М.Е. (1823 г.). Исследования жировых веществ животного происхождения . Париж: Левро.
- ^ Jump up to: Перейти обратно: а б с Лерэ С. (2012). Введение в липомику . Бока-Ратон: CRC Press. ISBN 978-1466551466 .
- ^ Лерэ С (2015). «Введение, история и эволюция». Липиды. Питание и здоровье . Бока-Ратон: CRC Press. ISBN 978-1482242317 .
- ^ Пелуз Т.Дж., Желис А. (1844 г.). «Воспоминания о масляной кислоте». Анналы химии и физики . 10 :434.
- ^ Еженедельные отчеты сессий Академии наук, Париж, 1853 г., 36, 27; Летопись химии и физики 1854, 41, 216.
- ^ Лере К. «Хронологическая история липидного центра» . Киберлипидный центр . Архивировано из оригинала 13 октября 2017 года . Проверено 1 декабря 2017 г.
- ^ Прут В. (1827 г.). «О конечном составе простых пищевых веществ, с некоторыми предварительными замечаниями по анализу организованных тел вообще» . Фил. Пер. : 355–388.
- ^ Каллинг CF (1974). «Липиды. (Жиры, Липоиды. Липины).». Справочник по гистопатологическим методам (3-е изд.). Лондон: Баттервортс. стр. 351–376. ISBN 978-1483164793 .
- ^ Розенблум Дж., Гис В.Дж. (1911). «Предложение учителям биохимии. I. Предложенная химическая классификация липинов с примечанием о тесной связи между холестеринами и солями желчных кислот» . Биохим. Бык . 1 : 51–56.
- ^ Блур В.Р. (1920). «Очерк классификации липидов» . Учеб. Соц. Эксп. Биол. Мед . 17 (6): 138–140. дои : 10.3181/00379727-17-75 . S2CID 75844378 .
- ^ Кристи WW, Хан Икс (2010). Анализ липидов: выделение, разделение, идентификация и липидомный анализ . Бриджуотер, Англия: The Oily Press. ISBN 978-0857097866 .
- ^ Бертран Г (1923). «Проект реформирования номенклатуры биологической химии» . Бюллетень Общества биологической химии . 5 : 96–109.
- ^ Хилдич, Томас Перси (1956). Химический состав натуральных жиров . Уайли.
- ^ Вэнс Дж. Э., Вэнс Д. Е. (2002). Биохимия липидов, липопротеинов и мембран . Амстердам: Эльзевир. ISBN 978-0-444-51139-3 .
- ^ Браун Х.А., изд. (2007). Липодомика и биоактивные липиды: анализ липидов на основе масс-спектрометрии . Методы энзимологии. Том. 423. Бостон: Академик Пресс. ISBN 978-0-12-373895-0 .
- ^ Хант С.М., Грофф Дж.Л., Гроппер С.А. (1995). Продвинутое питание и обмен веществ человека . Бельмонт, Калифорния: Западный паб. Компания р. 98 . ISBN 978-0-314-04467-9 .
- ^ Яшрой Р.Ц. (1987). " 13 Исследование ЯМР липидов липидных жирных ацильных цепей мембран хлоропластов» . Индийский журнал биохимии и биофизики . 24 (6): 177–178. doi : 10.1016/0165-022X(91)90019-S . PMID 3428918 .
- ^ Jump up to: Перейти обратно: а б Девлин Т.М. (1997). Учебник биохимии: с клиническими корреляциями (4-е изд.). Чичестер: Джон Уайли и сыновья. ISBN 978-0-471-17053-2 .
- ^ Хантер Дж. Э. (ноябрь 2006 г.). «Диетические трансжирные кислоты: обзор недавних исследований на людях и реакции пищевой промышленности». Липиды . 41 (11): 967–992. дои : 10.1007/s11745-006-5049-y . ПМИД 17263298 . S2CID 1625062 .
- ^ Фурс С (2 декабря 2011 г.). «Длинный липид, длинное название: докозагексаеновая кислота» . Липидные хроники .
- ^ «DHA для оптимального функционирования мозга и зрения» . Институт DHA/EPA Омега-3.
- ^ Фецца Ф, Де Симоне С, Амадио Д, Маккарроне М (2008). «Амидгидролаза жирных кислот: страж эндоканнабиноидной системы». Липиды в здоровье и болезни . Субклеточная биохимия. Том. 49. стр. 101–132. дои : 10.1007/978-1-4020-8831-5_4 . ISBN 978-1-4020-8830-8 . ПМИД 18751909 .
- ^ Коулман Р.А., Ли Д.П. (март 2004 г.). «Ферменты синтеза триацилглицеринов и их регуляция». Прогресс в исследованиях липидов . 43 (2): 134–176. дои : 10.1016/S0163-7827(03)00051-1 . ПМИД 14654091 .
- ^ Jump up to: Перейти обратно: а б ван Холде К.Е., Мэтьюз К.К. (1996). Биохимия (2-е изд.). Менло-Парк, Калифорния: Паб Benjamin/Cummings. компании ISBN 978-0-8053-3931-4 .
- ^ Jump up to: Перейти обратно: а б Хёльцль Г., Дорманн П. (сентябрь 2007 г.). «Структура и функции гликоглицеролипидов у растений и бактерий». Прогресс в исследованиях липидов . 46 (5): 225–243. дои : 10.1016/j.plipres.2007.05.001 . ПМИД 17599463 .
- ^ Хонке К., Чжан Й., Ченг Х., Котани Н., Танигучи Н. (2004). «Биологическая роль сульфогликолипидов и патофизиология их дефицита». Гликоконъюгатный журнал . 21 (1–2): 59–62. дои : 10.1023/B:GLYC.0000043749.06556.3d . ПМИД 15467400 . S2CID 2678053 .
- ^ «Строение мембраны» . Липидные хроники . 5 ноября 2011 года . Проверено 31 декабря 2011 г.
- ^ Берридж М.Дж., Ирвин РФ (сентябрь 1989 г.). «Инозитолфосфаты и передача сигналов в клетках». Природа . 341 (6239): 197–205. Бибкод : 1989Natur.341..197B . дои : 10.1038/341197a0 . ПМИД 2550825 . S2CID 26822092 .
- ^ Фаруки А.А., Хоррокс Л.А., Фаруки Т. (июнь 2000 г.). «Глицерофосфолипиды в головном мозге: их метаболизм, включение в мембраны, функции и участие в неврологических расстройствах». Химия и физика липидов . 106 (1): 1–29. дои : 10.1016/S0009-3084(00)00128-6 . ПМИД 10878232 .
- ^ Иванова П.Т., Милн С.Б., Бирн М.О., Сян Ю., Браун Х.А. (2007). «Идентификация и количественное определение глицерофосфолипидов методом масс-спектрометрии с ионизацией электрораспылением». Липидомика и биоактивные липиды: анализ липидов на основе масс-спектрометрии . Методы энзимологии. Том. 432. стр. 21–57. дои : 10.1016/S0076-6879(07)32002-8 . ISBN 978-0-12-373895-0 . ПМИД 17954212 .
- ^ Палтауф Ф (декабрь 1994 г.). «Эфирные липиды в биомембранах». Химия и физика липидов . 74 (2): 101–139. дои : 10.1016/0009-3084(94)90054-X . ПМИД 7859340 .
- ^ Меррилл А.Х., Сандофф К. (2002). «Глава 14: Сфинголипиды: метаболизм и передача сигналов в клетках» (PDF) . В Вэнсе Дж. Э., Вэнсе Э. Э. (ред.). Биохимия липидов, липопротеинов и мембран (4-е изд.). Амстердам: Эльзевир. стр. 373–407. ISBN 978-0-444-51138-6 .
- ^ Хори Т., Сугита М. (1993). «Сфинголипиды у низших животных». Прогресс в исследованиях липидов . 32 (1): 25–45. дои : 10.1016/0163-7827(93)90003-Ф . ПМИД 8415797 .
- ^ Вигандт Х. (январь 1992 г.). «Гликолипиды насекомых». Biochimica et Biophysical Acta (BBA) - Липиды и липидный обмен . 1123 (2): 117–126. дои : 10.1016/0005-2760(92)90101-Z . ПМИД 1739742 .
- ^ Гуань X, Венк М.Р. (май 2008 г.). «Биохимия инозитоллипидов» . Границы бионауки . 13 (13): 3239–3251. дои : 10.2741/2923 . ПМИД 18508430 .
- ^ Бах Д., Вахтель Э. (март 2003 г.). «Мембраны модели фосфолипида/холестерина: образование кристаллитов холестерина» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1610 (2): 187–197. дои : 10.1016/S0005-2736(03)00017-8 . ПМИД 12648773 .
- ^ Рассел Д.В. (2003). «Ферменты, регуляция и генетика синтеза желчных кислот». Ежегодный обзор биохимии . 72 : 137–174. doi : 10.1146/annurev.biochem.72.121801.161712 . ПМИД 12543708 .
- ^ Виллински Дж.К., Хейс Дж.М., Брасселл С.С., Риггерт В.Л., Данбар Р. (2008). «Осадочные стерины как биогеохимические индикаторы Южного океана». Органическая геохимия . 39 (5): 567–588. Бибкод : 2008OrGeo..39..567V . doi : 10.1016/j.orggeochem.2008.01.009 .
- ^ Дьякон Дж. (2005). Грибковая биология . Кембридж, Массачусетс: Издательство Blackwell. п. 342. ИСБН 978-1-4051-3066-0 .
- ^ Буйон Р., Верстюф А., Матье С., Ван Кромфо С., Масуяма Р., Дехаес П., Кармелиет Г. (декабрь 2006 г.). «Резистентность к витамину D». Лучшие практики и исследования. Клиническая эндокринология и обмен веществ . 20 (4): 627–645. дои : 10.1016/j.beem.2006.09.008 . ПМИД 17161336 .
- ^ Jump up to: Перейти обратно: а б с Кузуяма Т., Сето Х (апрель 2003 г.). «Разнообразие биосинтеза изопреновых единиц». Отчеты о натуральных продуктах . 20 (2): 171–183. дои : 10.1039/b109860h . ПМИД 12735695 .
- ^ Рао А.В., Рао Л.Г. (март 2007 г.). «Каротиноиды и здоровье человека». Фармакологические исследования . 55 (3): 207–216. дои : 10.1016/j.phrs.2007.01.012 . ПМИД 17349800 .
- ^ Брунмарк А, Каденас Э (1989). «Окислительно-восстановительная и аддитивная химия хиноидных соединений и ее биологическое значение». Свободно-радикальная биология и медицина . 7 (4): 435–477. дои : 10.1016/0891-5849(89)90126-3 . ПМИД 2691341 .
- ^ Свежевска Е, Даникевич В (июль 2005 г.). «Полиизопреноиды: строение, биосинтез и функции». Прогресс в исследованиях липидов . 44 (4): 235–258. дои : 10.1016/j.plipres.2005.05.002 . ПМИД 16019076 .
- ^ Jump up to: Перейти обратно: а б Раец Ч.Р., Гарретт Т.А., Рейнольдс К.М., Шоу В.А., Мур Дж.Д., Смит Д.С. и др. (май 2006 г.). «Kdo2-Липид А Escherichia coli, определенный эндотоксин, который активирует макрофаги через TLR-4» . Журнал исследований липидов . 47 (5): 1097–1111. doi : 10.1194/jlr.M600027-JLR200 . hdl : 10919/74310 . ПМИД 16479018 .
- ^ Уолш, Коннектикут (март 2004 г.). «Поликетидные и нерибосомальные пептидные антибиотики: модульность и универсальность». Наука . 303 (5665): 1805–1810. Бибкод : 2004Sci...303.1805W . дои : 10.1126/science.1094318 . ПМИД 15031493 . S2CID 44858908 .
- ^ Кэффри П., Апарисио Дж. Ф., Мальпартида Ф., Зотчев С.Б. (2008). «Биосинтетическая инженерия полиеновых макролидов для создания улучшенных противогрибковых и противопаразитарных средств» . Актуальные темы медицинской химии . 8 (8): 639–653. дои : 10.2174/156802608784221479 . hdl : 10197/8333 . ПМИД 18473889 .
- ^ Минто Р.Э., Блэклок БиДжей (июль 2008 г.). «Биосинтез и функции полиацетиленов и родственных им натуральных продуктов» . Прогресс в исследованиях липидов . 47 (4): 233–306. дои : 10.1016/j.plipres.2008.02.002 . ПМК 2515280 . ПМИД 18387369 .
- ^ Кунс RT, Грин Р.Дж., Фрейзер Р.А. (июль 2021 г.). «Исследование состава головных групп липидов в эпителиальных мембранах: систематический обзор» . Мягкая материя . 17 (28): 6773–6786. Бибкод : 2021SMat...17.6773C . дои : 10.1039/D1SM00703C . ISSN 1744-683X . ПМИД 34212942 . S2CID 235708094 .
- ^ Хайнц Э. (1996). «Растительные гликолипиды: структура, выделение и анализ», стр. 211–332 в журнале Advances in Lipid Methodology , Vol. 3. У. В. Кристи (ред.). Oily Press, Данди. ISBN 978-0-9514171-6-4
- ^ Лю, Цзябао; Гао, Ренджун; Го, Чжэн (2021). «Галактозилдиацилглицерины: от аппарата, связанного с фотосинтезом, к сборке in vitro с определенной структурой ». Журнал сельскохозяйственной и пищевой химии . 69 (32): 8910–8928. doi : 10.1021/acs.jafc.1c00204 . ПМИД 33793221 . S2CID 232761961 .
- ^ Яшрой Р.Ц. (1990). «Магнитно-резонансные исследования динамической организации липидов в мембранах хлоропластов». Журнал биологических наук . 15 (4): 281–288. дои : 10.1007/BF02702669 . S2CID 360223 .
- ^ ван Меер Г., Фелькер Д.Р., Фейгенсон Г.В. (февраль 2008 г.). «Мембранные липиды: где они находятся и как ведут себя» . Nature Reviews Молекулярно-клеточная биология . 9 (2): 112–124. дои : 10.1038/nrm2330 . ПМК 2642958 . ПМИД 18216768 .
- ^ Фейгенсон Г.В. (ноябрь 2006 г.). «Фазовое поведение смесей липидов» . Химическая биология природы . 2 (11): 560–563. дои : 10.1038/nchembio1106-560 . ПМК 2685072 . ПМИД 17051225 .
- ^ Виггинс ПМ (декабрь 1990 г.). «Роль воды в некоторых биологических процессах» . Микробиологические обзоры . 54 (4): 432–449. дои : 10.1128/MMBR.54.4.432-449.1990 . ПМЦ 372788 . ПМИД 2087221 .
- ^ Рашке Т.М., Левитт М. (май 2005 г.). «Неполярные растворенные вещества улучшают структуру воды внутри гидратных оболочек, одновременно уменьшая взаимодействие между ними» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (19): 6777–6782. дои : 10.1073/pnas.0500225102 . ПМЦ 1100774 . ПМИД 15867152 .
- ^ Сегре Д., Бен-Эли Д., Димер Д.В., Lancet Д. (2001). «Липидный мир» (PDF) . Происхождение жизни и эволюция биосферы . 31 (1–2): 119–145. Бибкод : 2001OLEB...31..119S . дои : 10.1023/А:1006746807104 . ПМИД 11296516 . S2CID 10959497 . Архивировано из оригинала (PDF) 11 сентября 2008 года . Проверено 15 марта 2015 г.
- ^ Брасамле Д.Л. (декабрь 2007 г.). «Серия тематических обзоров: биология адипоцитов. Семейство перилипинов структурных белков липидных капель: стабилизация липидных капель и контроль липолиза» . Журнал исследований липидов . 48 (12): 2547–2559. doi : 10.1194/jlr.R700014-JLR200 . ПМИД 17878492 .
- ^ Малинаускас Т., Арическу А.Р., Лу В., Зибольд С., Джонс Э.Ю. (июль 2011 г.). «Модульный механизм ингибирования передачи сигналов Wnt ингибирующим фактором 1 Wnt» . Структурная и молекулярная биология природы . 18 (8): 886–893. дои : 10.1038/nsmb.2081 . ПМК 3430870 . ПМИД 21743455 .
- ^ Малинаускас Т. (март 2008 г.). «Стыковка жирных кислот с доменом WIF человеческого фактора-1, ингибирующего Wnt». Липиды . 43 (3): 227–230. дои : 10.1007/s11745-007-3144-3 . ПМИД 18256869 . S2CID 31357937 .
- ^ Ван Икс (июнь 2004 г.). «Липидная сигнализация». Современное мнение в области биологии растений . 7 (3): 329–336. дои : 10.1016/j.pbi.2004.03.012 . ПМИД 15134755 .
- ^ Динасарапу А.Р., Сондерс Б., Озерлат И., Азам К., Субраманиам С. (июнь 2011 г.). «Страницы молекул сигнального шлюза – взгляд на модель данных» . Биоинформатика . 27 (12): 1736–1738. doi : 10.1093/биоинформатика/btr190 . ПМК 3106186 . ПМИД 21505029 .
- ^ Эйстер К.М. (март 2007 г.). «Мембрана и липиды как неотъемлемые участники передачи сигнала: передача липидного сигнала для нелипидного биохимика». Достижения в области физиологического образования . 31 (1): 5–16. дои : 10.1152/advan.00088.2006 . ПМИД 17327576 . S2CID 9194419 .
- ^ Хиньковска-Гальчева В., ВанВэй С.М., Шэнли Т.П., Кункель Р.Г. (ноябрь 2008 г.). «Роль сфингозин-1-фосфата и церамид-1-фосфата в гомеостазе кальция». Текущее мнение об исследуемых препаратах . 9 (11): 1192–1205. ПМИД 18951299 .
- ^ Саддуги С.А., Сонг П., Огретмен Б. (2008). «Роль биоактивных сфинголипидов в биологии и терапии рака». Липиды в здоровье и болезни . Субклеточная биохимия. Том. 49. стр. 413–440. дои : 10.1007/978-1-4020-8831-5_16 . ISBN 978-1-4020-8830-8 . ПМК 2636716 . ПМИД 18751921 .
- ^ Кляйн С., Мальвия А.Н. (январь 2008 г.). «Механизм ядерной передачи сигналов кальция с помощью инозитол-1,4,5-трифосфата, продуцируемого в ядре, ядерной протеинкиназы C и циклической AMP-зависимой протеинкиназы» . Границы бионауки . 13 (13): 1206–1226. дои : 10.2741/2756 . ПМИД 17981624 .
- ^ Бойс Дж. А. (август 2008 г.). «Эйкозаноиды при астме, аллергическом воспалении и защите организма». Современная молекулярная медицина . 8 (5): 335–349. дои : 10.2174/156652408785160989 . ПМИД 18691060 .
- ^ Белтовский Ю. (2008). «Х-рецепторы печени (LXR) как терапевтические мишени при дислипидемии» . Сердечно-сосудистая терапия . 26 (4): 297–316. дои : 10.1111/j.1755-5922.2008.00062.x . ПМИД 19035881 .
- ^ Бирманн М., Мауэредер С., Браунер Дж.М., Чаурио Р., Янко С., Херрманн М., Муньос Л.Е. (декабрь 2013 г.). «Поверхностный код - биофизические сигналы для апоптотического клиренса клеток». Физическая биология . 10 (6): 065007. Бибкод : 2013PhBio..10f5007B . дои : 10.1088/1478-3975/10/6/065007 . ПМИД 24305041 . S2CID 23782770 .
- ^ Индивери К, Тонацци А, Палмьери Ф (октябрь 1991 г.). «Характеристика однонаправленного транспорта карнитина, катализируемого восстановленным переносчиком карнитина из митохондрий печени крысы». Biochimica et Biophysical Acta (BBA) – Биомембраны . 1069 (1): 110–116. дои : 10.1016/0005-2736(91)90110-т . ПМИД 1932043 .
- ^ Пароди А.Дж., Лелуар Л.Ф. (апрель 1979 г.). «Роль липидных промежуточных продуктов в гликозилировании белков в эукариотической клетке». Biochimica et Biophysical Acta (BBA) — Обзоры биомембран . 559 (1): 1–37. дои : 10.1016/0304-4157(79)90006-6 . ПМИД 375981 .
- ^ Хелениус А., Эби М. (март 2001 г.). «Внутриклеточные функции N-связанных гликанов». Наука . 291 (5512): 2364–2369. Бибкод : 2001Sci...291.2364H . дои : 10.1126/science.291.5512.2364 . ПМИД 11269317 . S2CID 7277949 .
- ^ Новицкий М., Мюллер Ф., Френтцен М. (апрель 2005 г.). «Кардиолипинсинтаза Arabidopsis thaliana» . Письма ФЭБС . 579 (10): 2161–2165. дои : 10.1016/j.febslet.2005.03.007 . ПМИД 15811335 . S2CID 21937549 .
- ^ Гохил В.М., Гринберг М.Л. (февраль 2009 г.). «Биогенез митохондриальной мембраны: фосфолипиды и белки идут рука об руку» . Журнал клеточной биологии . 184 (4): 469–472. дои : 10.1083/jcb.200901127 . ПМЦ 2654137 . ПМИД 19237595 .
- ^ Хох, Флорида (март 1992 г.). «Кардиолипины и функция биомембраны» (PDF) . Biochimica et Biophysical Acta (BBA) — Обзоры биомембран . 1113 (1): 71–133. дои : 10.1016/0304-4157(92)90035-9 . hdl : 2027.42/30145 . ПМИД 1550861 .
- ^ «Стероиды» . Элмхерст. образование . Архивировано из оригинала 23 октября 2011 года . Проверено 10 октября 2013 г.
- ^ Чирала С.С., Вакил С.Дж. (ноябрь 2004 г.). «Структура и функция синтазы жирных кислот животных». Липиды . 39 (11): 1045–1053. дои : 10.1007/s11745-004-1329-9 . ПМИД 15726818 . S2CID 4043407 .
- ^ Уайт С.В., Чжэн Дж., Чжан Ю.М. (2005). «Структурная биология биосинтеза жирных кислот II типа». Ежегодный обзор биохимии . 74 : 791–831. doi : 10.1146/annurev.biochem.74.082803.133524 . ПМИД 15952903 .
- ^ Олрогге Дж.Б., Яворски Дж.Г. (июнь 1997 г.). «Регуляция синтеза жирных кислот». Ежегодный обзор физиологии растений и молекулярной биологии растений . 48 : 109–136. дои : 10.1146/annurev.arplant.48.1.109 . ПМИД 15012259 . S2CID 46348092 .
- ^ Гроховски Л.Л., Сюй Х., Уайт Р.Х. (май 2006 г.). «Methanocaldococcus jannaschii использует модифицированный мевалонатный путь для биосинтеза изопентенилдифосфата» . Журнал бактериологии . 188 (9): 3192–3198. дои : 10.1128/JB.188.9.3192-3198.2006 . ПМЦ 1447442 . ПМИД 16621811 .
- ^ Лихтенталер Гонконг (июнь 1999 г.). «1-дидезокси-D-ксилулозо-5-фосфатный путь биосинтеза изопреноидов в растениях». Ежегодный обзор физиологии растений и молекулярной биологии растений . 50 : 47–65. дои : 10.1146/annurev.arplant.50.1.47 . ПМИД 15012203 .
- ^ Jump up to: Перейти обратно: а б Шрёпфер Г.Дж. (1981). «Биосинтез стерола». Ежегодный обзор биохимии . 50 : 585–621. дои : 10.1146/annurev.bi.50.070181.003101 . ПМИД 7023367 .
- ^ Лиз Н.Д., Скэггс Б., Кирш Д.Р., Бард М. (март 1995 г.). «Клонирование поздних генов пути биосинтеза эргостерина Saccharomyces cerevisiae - обзор». Липиды . 30 (3): 221–226. дои : 10.1007/BF02537824 . ПМИД 7791529 . S2CID 4019443 .
- ^ Jump up to: Перейти обратно: а б Багаван Н.В. (2002). Медицинская биохимия . Сан-Диего: Харкорт/Академическая пресса. ISBN 978-0-12-095440-7 .
- ^ Руссо Г.Л. (март 2009 г.). «Диетические полиненасыщенные жирные кислоты n-6 и n-3: от биохимии к клиническому значению в профилактике сердечно-сосудистых заболеваний». Биохимическая фармакология . 77 (6): 937–946. дои : 10.1016/j.bcp.2008.10.020 . ПМИД 19022225 .
- ^ Ридигер Н.Д., Отман Р.А., Сух М., Могадасян М.Х. (апрель 2009 г.). «Системный обзор роли жирных кислот n-3 в здоровье и болезнях». Журнал Американской диетической ассоциации . 109 (4): 668–679. дои : 10.1016/j.jada.2008.12.022 . ПМИД 19328262 .
- ^ Галли С., Рисе П. (2009). «Потребление рыбы, жирные кислоты омега-3 и сердечно-сосудистые заболевания. Наука и клинические испытания». Питание и здоровье . 20 (1): 11–20. дои : 10.1177/026010600902000102 . ПМИД 19326716 . S2CID 20742062 .
- ^ Миша Р., Мозаффариан Д. (2008). «Трансжирные кислоты: влияние на кардиометаболическое здоровье и значение для политики» . Простагландины, лейкотриены и незаменимые жирные кислоты . 79 (3–5): 147–152. дои : 10.1016/j.plefa.2008.09.008 . ПМЦ 2639783 . ПМИД 18996687 .
- ^ Далайнас I, Иоанну HP (апрель 2008 г.). «Роль трансжирных кислот в атеросклерозе, сердечно-сосудистых заболеваниях и развитии младенцев». Международная ангиология . 27 (2): 146–156. ПМИД 18427401 .
- ^ Мозаффариан Д., Уиллетт У.К. (декабрь 2007 г.). «Трансжирные кислоты и сердечно-сосудистый риск: уникальный кардиометаболический отпечаток?». Текущие отчеты об атеросклерозе . 9 (6): 486–493. дои : 10.1007/s11883-007-0065-9 . ПМИД 18377789 . S2CID 24998042 .
- ^ Аструп А., Дайерберг Дж., Селлек М., Стендер С. (2008), «Переход в питании и его связь с развитием ожирения и связанных с ним хронических заболеваний», Obes Rev , 9 (S1): 48–52, doi : 10.1111/j. 1467-789X.2007.00438.x , PMID 18307699 , S2CID 34030743
- ^ Аструп А (февраль 2005 г.). «Роль пищевых жиров в ожирении». Семинары по сосудистой медицине . 5 (1): 40–47. дои : 10.1055/s-2005-871740 . ПМИД 15968579 . S2CID 260372605 .
- ^ Аструп А (2008). «Диетическое лечение ожирения». Журнал парентерального и энтерального питания . 32 (5): 575–577. дои : 10.1177/0148607108321707 . ПМИД 18753397 .
- ^ Бересфорд С.А., Джонсон К.С., Ритенбо С., Лассер Н.Л., Снетселар Л.Г., Блэк Х.Р. и др. (февраль 2006 г.). «Обезжиренная диета и риск колоректального рака: рандомизированное контролируемое исследование модификации диеты Инициативы по охране здоровья женщин». Журнал Американской медицинской ассоциации . 295 (6): 643–654. дои : 10.1001/jama.295.6.643 . ПМИД 16467233 .
- ^ Ховард Б.В., Мэнсон Дж.Э., Стефаник М.Л., Бересфорд С.А., Фрэнк Г., Джонс Б., Родабо Р.Дж., Снетселаар Л., Томсон С., Тинкер Л., Витолинс М., Прентис Р. (январь 2006 г.). «Обезжиренная диета и изменение веса за 7 лет: исследование модификации диеты Инициативы по здоровью женщин». Журнал Американской медицинской ассоциации . 295 (1): 39–49. дои : 10.1001/jama.295.1.39 . ПМИД 16391215 .
- ^ «Источник питания» . Школа общественного здравоохранения Т.Х.Чана . Гарвардский университет.
- ^ «Жиры и холестерин: долой плохое, добавим хорошее — что следует есть? — Источник питания» . Гарвардская школа общественного здравоохранения.
Библиография [ править ]
- Багаван Н.В. (2002). Медицинская биохимия . Сан-Диего: Харкорт/Академическая пресса. ISBN 978-0-12-095440-7 .
- Девлин Т.М. (1997). Учебник биохимии: с клиническими корреляциями (4-е изд.). Чичестер: Джон Уайли и сыновья. ISBN 978-0-471-17053-2 .
- Страйер Л., Берг Дж.М., Тимочко Дж.Л. (2007). Биохимия (6-е изд.). Сан-Франциско: WH Freeman. ISBN 978-0-7167-8724-2 .
- ван Холде К.Е., Мэтьюз К.К. (1996). Биохимия (2-е изд.). Менло-Парк, Калифорния: Паб Benjamin/Cummings. компании ISBN 978-0-8053-3931-4 .
Внешние ссылки [ править ]
Вводный
- Список веб-сайтов, посвященных липидам
- Nature Lipidomics Gateway – Обзор и резюме недавних исследований липидов
- Липидная библиотека - Общий справочник по химии и биохимии липидов.
- Cyberlipid.org – Ресурсы и история липидов.
- Молекулярное компьютерное моделирование – моделирование липидных мембран
- Липиды, мембраны и торговля везикулами - Виртуальная библиотека биохимии, молекулярной биологии и клеточной биологии
Номенклатура
Базы данных
- LIPID MAPS – Комплексные базы данных липидов и липид-ассоциированных генов/белков.
- LipidBank - японская база данных липидов и связанных с ними свойств, спектральных данных и ссылок.
Общий
- ApolloLipids – предоставляет информацию о профилактике и лечении дислипидемии и сердечно-сосудистых заболеваний, а также программы непрерывного медицинского образования.
- Национальная липидная ассоциация – профессиональная медицинская образовательная организация для медицинских работников, стремящихся предотвратить заболеваемость и смертность, вызванную дислипидемиями и другими нарушениями, связанными с холестерином.
Липиды : метаболизм липопротеиновых частиц . |
|---|