ЛРП5
| ЛРП5 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | LRP5 , BMND1, EVR1, EVR4, HBM, LR3, LRP-5, LRP7, OPPG, OPS, OPTA1, VBCH2, белок 5, родственный рецептору ЛПНП, PCLD4, LRP-7 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 603506 ; МГИ : 1278315 ; Гомологен : 1746 ; Генные карты : LRP5 ; ОМА : LRP5 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Белок 5, родственный рецептору липопротеинов низкой плотности, представляет собой белок , который у человека кодируется LRP5 геном . [5] [6] [7] LRP5 является ключевым компонентом LRP5/ LRP6 / Frizzled группы корецепторов , которая участвует в каноническом пути Wnt . Мутации LRP5 могут привести к значительным изменениям костной массы. Мутация потери функции вызывает синдром псевдоглиомы остеопороза с уменьшением костной массы, тогда как мутация приобретения функции вызывает резкое увеличение костной массы.
Структура
[ редактировать ]низкой плотности липопротеинов LRP5 представляет собой трансмембранный рецептор , имеющий структуру, аналогичную LRP6 . В каждом белке около 85% его длины, состоящей из 1600 аминокислот, находится внеклеточно. Каждый из них имеет четыре мотива β-пропеллера на аминоконце, которые чередуются с четырьмя повторами, подобными эпидермальному фактору роста (EGF). Большинство внеклеточных лигандов связываются с LRP5 и LRP6 на β-пропеллерах. Каждый белок имеет однопроходной сегмент из 22 аминокислот, который пересекает клеточную мембрану, и сегмент из 207 аминокислот, который находится внутри клетки. [8]
Функция
[ редактировать ]LRP5 действует как корецептор с LRP6 и членами семейства белков Frizzled для передачи сигналов белками Wnt через канонический путь Wnt . [8] Этот белок играет ключевую роль в гомеостазе скелета. [7]
Транскрипция
[ редактировать ]LRP5 Промотор содержит сайты связывания для KLF15 и SP1 . [9] Кроме того, 5'-область гена LRP5 содержит четыре сайта связывания RUNX2 . [10] На мышах и людях было показано, что LRP5 ингибирует экспрессию TPH1 , фермента, ограничивающего скорость биосинтеза серотонина, в энтерохромаффинных клетках двенадцатиперстной кишки. [11] [12] [13] [14] [15] [16] и что избыток серотонина в плазме приводит к угнетению костей. С другой стороны, одно исследование на мышах показало прямое влияние Lrp5 на кости. [17]
Взаимодействия
[ редактировать ]Было показано, что LRP5 взаимодействует с AXIN1 . [18] [19]
Канонические сигналы WNT передаются через рецептор Frizzled и корецептор LRP5/ LRP6 для подавления активности GSK3beta ( GSK3B Ser-9 ), не зависящей от фосфорилирования . [20] Снижение канонических сигналов Wnt при истощении LRP5 и LRP6 приводит к деградации p120- катенина . [21]
Клиническое значение
[ редактировать ]был Сигнальный путь Wnt впервые связан с развитием костей, когда было обнаружено, что мутация с потерей функции в LRP5 вызывает синдром остеопороза-псевдоглиомы. [22] Вскоре после этого в двух исследованиях сообщалось, что мутации LRP5 , приводящие к увеличению функции, вызывают увеличение костной массы. [23] [24] Многие заболевания, связанные с плотностью костной ткани, вызваны мутациями гена LRP5. Существуют разногласия по поводу того, растет ли кость посредством Lrp5 через кость или через кишечник. [25] Большинство текущих данных подтверждают концепцию о том, что костная масса контролируется LRP5 через остеоциты. [26] Мыши с теми же мутациями усиления функции Lrp5 также имеют высокую костную массу. [27] Высокая костная масса сохраняется, когда мутация возникает только в конечностях или в клетках остеобластной линии. [17] Костная механотрансдукция происходит через Lrp5. [28] и подавляется, если Lrp5 удаляется только из остеоцитов . [29] Есть многообещающие клинические исследования остеопороза, нацеленные на склеростин , специфичный для остеоцитов белок, который ингибирует передачу сигналов Wnt путем связывания с Lrp5. [26] [30] Альтернативная модель, которая была проверена на мышах и людях, заключается в том, что Lrp5 контролирует образование кости путем ингибирования экспрессии TPH1 , фермента, ограничивающего скорость биосинтеза серотонина , молекулы, которая регулирует образование кости, в энтерохромаффинных клетках двенадцатиперстной кишки. [11] [12] [13] [14] [15] [16] и что избыток серотонина в плазме приводит к угнетению костей. Другое исследование показало, что другой ингибитор Tph1 снижает уровень серотонина в крови и кишечнике, но не влияет на костную массу или маркеры костеобразования. [17]
LRP5 может быть важен для развития сосудистой сети сетчатки и может играть роль в созревании капилляров. [31] Мутации в этом гене также вызывают семейную экссудативную витреоретинопатию . [7]
Внеклеточный лиганд глиального происхождения, Norrin, действует на трансмембранный рецептор Frizzled4 , корецептор Lrp5 и вспомогательный мембранный белок TSPAN12 на поверхности развивающихся эндотелиальных клеток, контролируя программу транскрипции, которая регулирует рост и созревание эндотелия. [32]
Нокаут LRP5 у мышей приводил к повышению уровня холестерина в плазме на диете с высоким содержанием жиров из-за снижения печеночного клиренса остатков хиломикронов . При нормальном питании у мышей с дефицитом LRP5 наблюдалось заметное нарушение толерантности к глюкозе с заметным снижением внутриклеточного АТФ и кальция. 2+ в ответ на глюкозу и нарушение секреции инсулина, индуцированной глюкозой. Производство IP3 в ответ на глюкозу также было снижено в LRP5-островках, что, возможно, вызвано заметным снижением различных транскриптов генов, участвующих в чувствительности к глюкозе в LRP5-островках. Островки с дефицитом LRP5 лишены Wnt-3a -стимулированной секреции инсулина. Эти данные позволяют предположить, что передача сигналов WntLRP5 способствует индуцированной глюкозой секреции инсулина в островках. [33]
В больных остеоартритом, хондроцитах, путь Wnt/бета-катенин активируется со значительным усилением экспрессии мРНК бета-катенина. Экспрессия мРНК и белка LRP5 также значительно усиливается в хряще, пораженном остеоартритом, по сравнению с нормальным хрящом, а экспрессия мРНК LRP5 дополнительно увеличивается под действием витамина D. Блокирование экспрессии LRP5 с использованием миРНК против LRP5 приводило к значительному снижению экспрессии мРНК и белка MMP13 . Катаболическая роль LRP5 , по-видимому, опосредована путем Wnt/бета-катенин при остеоартрите человека. [34]
Полифенол куркумин увеличивает экспрессию мРНК LRP5. [35]
Мутации в LRP5 вызывают поликистоз печени . [36]
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000162337 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000024913 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Эй, П.Дж., Твеллс Р.К., Филлипс М.С., Браун С.Д., Кавагути Ю., Кокс Р., Дуган В., Хаммонд Х., Мецкер М.Л., Тодд Дж.А., Хесс Дж.Ф. (август 1998 г.). «Клонирование нового члена семейства рецепторов липопротеинов низкой плотности». Джин . 216 (1): 103–11. дои : 10.1016/S0378-1119(98)00311-4 . ПМИД 9714764 .
- ^ Чен Д., Латроп В., Донг Ю (февраль 1999 г.). «Молекулярное клонирование кДНК мышиного Lrp7 (Lr3) и хромосомное картирование ортологичных генов у мыши и человека». Геномика . 55 (3): 314–21. дои : 10.1006/geno.1998.5688 . ПМИД 10049586 .
- ^ Jump up to: а б с «Ген Энтрез: белок 5, родственный рецептору липопротеина низкой плотности LRP5» .
- ^ Jump up to: а б Уильямс Б.О., Insogna KL (февраль 2009 г.). «Куда пошел Wnts: взрывное поле передачи сигналов Lrp5 и Lrp6 в кости» . Журнал исследований костей и минералов . 24 (2): 171–8. дои : 10.1359/jbmr.081235 . ПМК 3276354 . ПМИД 19072724 .
- ^ Ли Дж, Ян Ю, Цзян Б, Чжан Икс, Цзоу Ю, Гун Ю (2010). «Sp1 и KLF15 регулируют базальную транскрипцию гена LRP5 человека» . БМК Генетика . 11:12 . дои : 10.1186/1471-2156-11-12 . ПМЦ 2831824 . ПМИД 20141633 .
- ^ Агеда Л., Веласкес-Крус Р., Уррейсти Р., Йосковиц Г., Саррион П., Хурадо С., Герри Р., Гарсиа-Жиральт Н., Ног Х., Меллибовский Л., Диес-Перес А., Мари П.Дж., Балселлс С., Гринберг Д. (май 2011 г.) ). «Функциональная значимость полиморфизма rs312009, связанного с МПК: новое участие RUNX2 в регуляции транскрипции LRP5» . Журнал исследований костей и минералов . 26 (5): 1133–44. дои : 10.1002/jbmr.293 . ПМИД 21542013 . S2CID 20985443 .
- ^ Jump up to: а б Ядав В.К., Рю Дж.Х., Суда Н., Танака К.Ф., Гингрич Дж.А., Шютц Г., Глориё Ф.Х., Чанг С.И., Заджак Дж.Д., Инсогна К.Л., Манн Дж.Дж., Хен Р., Дьюси П., Карсенти Г. (ноябрь 2008 г.). «Lrp5 контролирует образование костной ткани, ингибируя синтез серотонина в двенадцатиперстной кишке» . Клетка . 135 (5): 825–37. дои : 10.1016/j.cell.2008.09.059 . ПМЦ 2614332 . ПМИД 19041748 .
- ^ Jump up to: а б Коде А, Мосиалоу И, Силва БК, Рашед МТ, Чжоу Б, Ван Дж, Таунс ТМ, Хен Р, ДеПиньо Р.А., Го ХЕ, Кустени С (октябрь 2012 г.). «FOXO1 управляет функцией серотонина, полученного из кишечника, по подавлению костей» . Журнал клинических исследований . 122 (10): 3490–503. дои : 10.1172/JCI64906 . ПМК 3461930 . ПМИД 22945629 .
- ^ Jump up to: а б Фрост М., Андерсен Т.Е., Ядав В., Бриксен К., Карсенти Г., Кассем М. (март 2010 г.). «Пациенты с фенотипом высокой костной массы из-за мутации Lrp5-T253I имеют низкие уровни серотонина в плазме» . Журнал исследований костей и минералов . 25 (3): 673–5. дои : 10.1002/jbmr.44 . ПМИД 20200960 . S2CID 24280062 .
- ^ Jump up to: а б Розен CJ (февраль 2009 г.). «Прорыв в биологию костей: секреты серотонина». Природная медицина . 15 (2): 145–6. дои : 10.1038/nm0209-145 . ПМИД 19197289 . S2CID 5489589 .
- ^ Jump up to: а б Мёддер У.И., Ахенбах С.Дж., Амин С., Риггс Б.Л., Мелтон Л.Дж., Хосла С. (февраль 2010 г.). «Связь уровня серотонина в сыворотке крови с плотностью костей и структурными параметрами у женщин» . Журнал исследований костей и минералов . 25 (2): 415–22. дои : 10.1359/jbmr.090721 . ПМК 3153390 . ПМИД 19594297 .
- ^ Jump up to: а б Фрост М., Андерсен Т., Госсиел Ф., Хансен С., Боллерслев Дж., ван Хал В., Истелл Р., Кассем М., Бриксен К. (август 2011 г.). «Уровни серотонина, склеростина, маркеров костного обмена, а также плотность и микроархитектура костной ткани у пациентов с фенотипом высокой костной массы из-за мутации Lrp5» . Журнал исследований костей и минералов . 26 (8): 1721–8. дои : 10.1002/jbmr.376 . ПМИД 21351148 . S2CID 28504199 .
- ^ Jump up to: а б с Цуй Ю, Низиолек П.Дж., Макдональд Б.Т., Зилстра Ч.Р., Аленина Н., Робинсон Д.Р., Чжун З., Маттес С., Якобсен К.М., Конлон Р.А., Броммейдж Р., Лю К., Мси Ф., Пауэлл Д.Р., Ян К.М., Замбровиц Б., Герритс Х. , Госсен Дж.А., Хе Х, Бадер М., Уильямс Б.О., Уорман М.Л., Роблинг А.Г. (июнь 2011 г.). «Lrp5 функционирует в кости, регулируя костную массу» . Природная медицина . 17 (6): 684–91. дои : 10.1038/нм.2388 . ПМЦ 3113461 . ПМИД 21602802 .
- ^ Мао Дж., Ван Дж., Лю Б., Пан В., Фарр Г.Х., Флинн С., Юань Х., Такада С., Кимельман Д., Ли Л., Ву Д. (апрель 2001 г.). «Белок-5, родственный рецептору липопротеинов низкой плотности, связывается с аксином и регулирует канонический сигнальный путь Wnt» . Молекулярная клетка . 7 (4): 801–9. дои : 10.1016/S1097-2765(01)00224-6 . ПМИД 11336703 .
- ^ Ким М.Дж., Чиа IV, Константини Ф. (ноябрь 2008 г.). «Целевые сайты СУМОилирования на С-конце защищают аксин от убиквитинирования и придают белку стабильность» . Журнал ФАСЭБ . 22 (11): 3785–94. дои : 10.1096/fj.08-113910 . ПМК 2574027 . ПМИД 18632848 .
- ^ Като М, Като М (сентябрь 2006 г.). «Перекрестное взаимодействие сигнальных путей WNT и FGF на GSK3beta для регуляции сигнальных каскадов бета-катенина и SNAIL» . Биология и терапия рака . 5 (9): 1059–64. дои : 10.4161/cbt.5.9.3151 . ПМИД 16940750 .
- ^ Хонг Дж.И., Пак Дж.И., Чо К., Гу Д., Джи Х., Артанди С.Э., МакКри П.Д. (декабрь 2010 г.). «Общие молекулярные механизмы регулируют множество белков катенина: канонические сигналы и компоненты Wnt модулируют изоформу-1 p120-катенина и дополнительные члены подсемейства p120» . Журнал клеточной науки . 123 (Часть 24): 4351–65. дои : 10.1242/jcs.067199 . ПМЦ 2995616 . ПМИД 21098636 .
- ^ Гонг Ю., Сли Р.Б., Фукаи Н., Равади Г., Роман-Роман С., Регинато А.М. и др. (ноябрь 2001 г.). «Белок 5, связанный с рецептором ЛПНП (LRP5), влияет на наращивание костей и развитие глаз» . Клетка . 107 (4): 513–23. дои : 10.1016/S0092-8674(01)00571-2 . ПМИД 11719191 . S2CID 1631509 .
- ^ Литтл Р.Д., Карулли Дж.П., Дель Мастро Р.Г., Дюпюи Дж. , Осборн М., Фольц К., Мэннинг С.П., Суэйн П.М., Чжао С.К., Юстас Б., Лаппе М.М., Спитцер Л., Цвайер С., Брауншвейгер К., Бенчекрун Ю., Ху Х, Адэр Р, Чи Л, Фитцджеральд М.Г., Тулиг С., Карузо А., Целлас Н., Бава А., Франклин Б., Макгуайр С., Ногс Х, Гонг Дж., Аллен К.М., Анисович А., Моралес А.Дж., Ломедико П.Т., Рекер С.М., Ван Эрдевег П. , Рекер Р.Р., Джонсон М.Л. (январь 2002 г.). «Мутация в гене белка 5, связанного с рецептором ЛПНП, приводит к аутосомно-доминантному признаку высокой костной массы» . Американский журнал генетики человека . 70 (1): 11–9. дои : 10.1086/338450 . ПМК 419982 . ПМИД 11741193 .
- ^ Бойден Л.М., Мао Дж., Бельский Дж., Мицнер Л., Фархи А., Митник М.А., Ву Д., Инсогна К., Лифтон Р.П. (май 2002 г.). «Высокая плотность костной ткани из-за мутации белка 5, связанного с рецептором ЛПНП» . Медицинский журнал Новой Англии . 346 (20): 1513–21. doi : 10.1056/NEJMoa013444 . ПМИД 12015390 .
- ^ Чжан В., Дрейк М.Т. (март 2012 г.). «Потенциальная роль терапии, нацеленной на DKK1, LRP5 и серотонин, в лечении остеопороза». Текущие отчеты об остеопорозе . 10 (1): 93–100. дои : 10.1007/s11914-011-0086-8 . ПМИД 22210558 . S2CID 23718294 .
- ^ Jump up to: а б Барон Р., Кнейссель М. (февраль 2013 г.). «Передача сигналов WNT в костном гомеостазе и заболеваниях: от мутаций человека до лечения». Природная медицина . 19 (2): 179–92. дои : 10.1038/нм.3074 . ПМИД 23389618 . S2CID 19968640 .
- ^ Бабий П., Чжао В., Смолл С., Хароде Ю., Яворски П.Дж., Буксейн М.Л., Редди П.С., Бодин П.В., Робинсон Дж.А., Бхат Б., Марзольф Дж., Моран Р.А., Бекс Ф. (июнь 2003 г.). «Высокая костная масса у мышей, экспрессирующих мутантный ген LRP5». Журнал исследований костей и минералов . 18 (6): 960–74. дои : 10.1359/jbmr.2003.18.6.960 . ПМИД 12817748 . S2CID 36863658 .
- ^ Саваками К., Роблинг А.Г., Ай М., Питнер Н.Д., Лю Д., Уорден С.Дж., Ли Дж., Мэй П., Роу Д.В., Дункан Р.Л., Уорман М.Л., Тернер Ч.Х. (август 2006 г.). «Корецептор Wnt LRP5 необходим для скелетной механотрансдукции, но не для анаболического ответа костей на лечение паратироидным гормоном» . Журнал биологической химии . 281 (33): 23698–711. дои : 10.1074/jbc.M601000200 . ПМИД 16790443 .
- ^ Чжао Л., Шим Дж.В., Додж Т.Р., Роблинг А.Г., Йокота Х. (май 2013 г.). «Инактивация Lrp5 в остеоцитах снижает модуль Юнга и чувствительность к механической нагрузке» . Кость . 54 (1): 35–43. дои : 10.1016/j.bone.2013.01.033 . ПМК 3602226 . ПМИД 23356985 .
- ^ Бургерс Т.А., Уильямс Б.О. (июнь 2013 г.). «Регуляция передачи сигналов Wnt/β-катенина внутри и из остеоцитов» . Кость . 54 (2): 244–9. дои : 10.1016/j.bone.2013.02.022 . ПМЦ 3652284 . ПМИД 23470835 .
- ^ Ся Ч., Лю Х., Чунг Д., Ван М., Ченг С., Ду Икс, Чанг Б., Бейтлер Б., Гонг Икс (июнь 2008 г.). «Модель семейной экссудативной витреоретинопатии, вызванной мутациями LPR5» . Молекулярная генетика человека . 17 (11): 1605–12. дои : 10.1093/hmg/ddn047 . ПМЦ 2902293 . ПМИД 18263894 .
- ^ Йе X, Ван Й, Натанс Дж (сентябрь 2010 г.). «Сигнальный путь Norrin/Frizzled4 в развитии и заболевании сосудов сетчатки» . Тенденции молекулярной медицины . 16 (9): 417–25. doi : 10.1016/j.molmed.2010.07.003 . ПМК 2963063 . ПМИД 20688566 .
- ^ Фуджино Т., Асаба Х., Кан М.Дж., Икеда Ю., Соне Х., Такада С., Ким Д.Х., Иока Р.С., Оно М., Томойори Х., Окубо М., Мурасе Т., Каматаки А., Ямамото Дж., Магори К., Такахаши С., Миямото Ю. , Оиси Х., Нос М., Окадзаки М., Усуи С., Имаидзуми К., Янагисава М., Сакаи Дж., Ямамото Т.Т. (январь 2003 г.). «Белок 5, связанный с рецептором липопротеинов низкой плотности (LRP5), необходим для нормального метаболизма холестерина и секреции инсулина, индуцированной глюкозой» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (1): 229–34. Бибкод : 2003PNAS..100..229F . дои : 10.1073/pnas.0133792100 . ПМК 140935 . ПМИД 12509515 .
- ^ Папатанасиу И., Мализос К.Н., Цезу А. (март 2010 г.). «Экспрессия белка 5, связанного с рецептором липопротеинов низкой плотности (LRP5), в хондроцитах человека, больных остеоартритом» . Журнал ортопедических исследований . 28 (3): 348–53. дои : 10.1002/jor.20993 . ПМИД 19810105 . S2CID 13525881 .
- ^ Ан Дж., Ли Х., Ким С., Ха Т (июнь 2010 г.). «Вызванное куркумином подавление адипогенной дифференцировки сопровождается активацией передачи сигналов Wnt/бета-катенин». Американский журнал физиологии. Клеточная физиология . 298 (6): C1510–6. doi : 10.1152/ajpcell.00369.2009 . ПМИД 20357182 . S2CID 25514556 .
- ^ Кноссен В.Р., Те Морше Р.Х., Хойшен А., Гилиссен С., Криспейн М., Венселаар Х., Мехди С., Бергманн С., Велтман Дж.А., Дрент Дж.П. (апрель 2014 г.). «Секвенирование всего экзома выявляет мутации LRP5 и каноническую передачу сигналов Wnt, связанную с цистогенезом печени» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (14): 5343–8. Бибкод : 2014PNAS..111.5343C . дои : 10.1073/pnas.1309438111 . ПМЦ 3986119 . ПМИД 24706814 .
Дальнейшее чтение
[ редактировать ]- Хэ X, Семенов М, Тамай К, Цзэн X (апрель 2004 г.). «Белки 5 и 6, связанные с рецептором ЛПНП, в передаче сигналов Wnt/бета-катенин: стрелки указывают путь». Разработка . 131 (8): 1663–77. дои : 10.1242/dev.01117 . ПМИД 15084453 . S2CID 2297859 .
- Година С., Лиау Г., Попа И., Стефанссон С., Аргрейвс В.С. (июнь 1995 г.). «Идентификация белка, связанного с рецептором липопротеинов низкой плотности (LRP), как эндоцитарного рецептора тромбоспондина-1» . Журнал клеточной биологии . 129 (5): 1403–10. дои : 10.1083/jcb.129.5.1403 . ПМК 2120467 . ПМИД 7775583 .
- Гонг Ю, Виккула М, Бун Л, Лю Дж, Бейтон П, Рамесар Р, Пелтонен Л, Сомер Х, Хиросе Т, Даллапиккола Б, Де Паепе А, Свобода В, Забель Б, Суперти-Фурга А, Штайнманн Б, Бруннер Х.Г. , Янс А., Болес Р.Г., Адкинс В., ван ден Бугаард М.Дж., Олсен Б.Р., Уорман М.Л. (июль 1996 г.). «Синдром остеопороза-псевдоглиомы, заболевание, влияющее на прочность скелета и зрение, отнесен к области хромосомы 11q12-13» . Американский журнал генетики человека . 59 (1): 146–51. ПМК 1915094 . ПМИД 8659519 .
- Джонсон М.Л., Гонг Дж., Кимберлинг В., Рекер С.М., Киммел Д.Б., Рекер Р.Б. (июнь 1997 г.). «Связь гена, вызывающего высокую костную массу, с хромосомой 11 человека (11q12-13)» . Американский журнал генетики человека . 60 (6): 1326–32. дои : 10.1086/515470 . ПМК 1716125 . ПМИД 9199553 .
- Донг Ю, Латроп В., Уивер Д., Цю Ц, Чини Дж., Бертолини Д., Чен Д. (октябрь 1998 г.). «Молекулярное клонирование и характеристика LR3, нового белка семейства рецепторов ЛПНП с митогенной активностью». Связь с биохимическими и биофизическими исследованиями . 251 (3): 784–90. дои : 10.1006/bbrc.1998.9545 . ПМИД 9790987 .
- де Креккио Г, Симонелли Ф, Нунциата Г, Маццео С, Греко ГМ, Ринальди Э, Вентруто В, Чиккодикола А, Миано МГ, Теста Ф, Курчи А, Д'Урсо М, Ринальди ММ, Кавальере МЛ, Кастеллуччио П (октябрь 1998 г.) ). «Аутосомно-рецессивная семейная экссудативная витреоретинопатия: доказательства генетической гетерогенности». Клиническая генетика . 54 (4): 315–20. дои : 10.1034/j.1399-0004.1998.5440409.x . ПМИД 9831343 . S2CID 37420287 .
- Мао Дж., Ван Дж., Лю Б., Пан В., Фарр Г.Х., Флинн С., Юань Х., Такада С., Кимельман Д., Ли Л., Ву Д. (апрель 2001 г.). «Белок-5, родственный рецептору липопротеинов низкой плотности, связывается с аксином и регулирует канонический сигнальный путь Wnt» . Молекулярная клетка . 7 (4): 801–9. дои : 10.1016/S1097-2765(01)00224-6 . ПМИД 11336703 .
- Твеллс Р.К., Мецкер М.Л., Браун С.Д., Кокс Р., Гэри С., Хаммонд Х., Эй П.Дж., Леви Э., Накагава Ю., Филипс М.С., Тодд Дж.А., Хесс Дж.Ф. (март 2001 г.). «Последовательность и характеристика гена кандидатной области размером 400 т.п.н. для IDDM4 на хромосоме 11q13». Геномика . 72 (3): 231–42. дои : 10.1006/geno.2000.6492 . ПМИД 11401438 .
- Семенов М.В., Тамай К., Бротт Б.К., Кюль М., Сокол С., Хе Х (июнь 2001 г.). «Головной индуктор Dickkopf-1 является лигандом корецептора Wnt LRP6» . Современная биология . 11 (12): 951–61. Бибкод : 2001CBio...11..951S . дои : 10.1016/S0960-9822(01)00290-1 . ПМИД 11448771 . S2CID 15702819 .
- Зорн А.М. (август 2001 г.). «Сигнализация Wnt: антагонистические Диккопфы» . Современная биология . 11 (15): R592–5. Бибкод : 2001CBio...11.R592Z . дои : 10.1016/S0960-9822(01)00360-8 . ПМИД 11516963 . S2CID 14970864 .
- Окубо М., Хориниси А., Ким Д.Х., Ямамото Т.Т., Мурасе Т. (февраль 2002 г.). «Семь новых вариантов последовательности гена белка 5, родственного рецептору липопротеина низкой плотности человека (LRP5)». Человеческая мутация . 19 (2): 186. дои : 10.1002/humu.9012 . ПМИД 11793484 . S2CID 41880501 .
- Ван Хул Э., Грэм Дж., Боллерслев Дж., Ван Везенбек Л., Матисен Д., Андерсен П.Е., Ванхонакер Ф., Ван Хул В. (июнь 2002 г.). «Локализация гена, вызывающего аутосомно-доминантный остеопетроз I типа, на хромосоме 11q12-13». Журнал исследований костей и минералов . 17 (6): 1111–7. дои : 10.1359/jbmr.2002.17.6.1111 . ПМИД 12054167 . S2CID 8308650 .
- Ван Везенбек Л., Клейрен Э., Грэм Дж., Билс Р.К., Бенишу О., Скопеллити Д., Ки Л., Рентон Т., Бартельс К., Гонг Ю., Уорман М.Л., Де Вернежуль М.К., Боллерслев Дж., Ван Халл В. (март 2003 г.). «Шесть новых миссенс-мутаций в гене белка 5, связанного с рецептором ЛПНП (LRP5), в различных условиях с повышенной плотностью костей» . Американский журнал генетики человека . 72 (3): 763–71. дои : 10.1086/368277 . ПМК 1180253 . ПМИД 12579474 .
Внешние ссылки
[ редактировать ]Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .
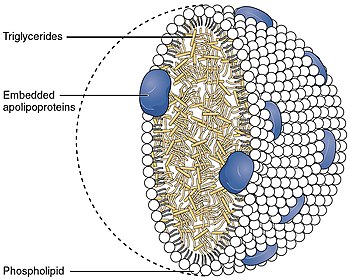
Липиды : метаболизм липопротеиновых частиц . |
|---|