Кортизол
 | |
 | |
| Имена | |
|---|---|
| Название ИЮПАК 11β,17α,21-тригидроксипрегн-4-ен-3,20-дион | |
| Систематическое название ИЮПАК (1 R ,3a S ,3b S ,9a R ,9b S ,11a S )-1,10-Дигидрокси-1-(гидроксиацетил)-9a,11a-диметил-1,2,3,3a,3b,4, 5,8,9,9a,9b,10,11,11a-тетрадекагидро-7H - циклопента[ а ]фенантен-7-он | |
| Идентификаторы | |
3D model ( JSmol ) | |
| ЧЭБИ | |
| ЧЕМБЛ | |
| ХимическийПаук | |
| Лекарственный Банк | |
| Информационная карта ECHA | 100.000.019 |
| КЕГГ | |
ПабХим CID | |
| НЕКОТОРЫЙ | |
Панель управления CompTox ( EPA ) | |
| Характеристики | |
| С 21 Н 30 О 5 | |
| Молярная масса | 362.460 g/mol |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
Кортизол — это стероидный гормон из класса глюкокортикоидов и гормон стресса . При использовании в качестве лекарства он известен как гидрокортизон .
Он вырабатывается у многих животных, главным образом пучковой коры надпочечников , оболочкой . [1] В других тканях он вырабатывается в меньших количествах. [2] В соответствии с дневным циклом уровень кортизола высвобождается и увеличивается в ответ на стресс и низкую концентрацию глюкозы в крови . [1] Он повышает уровень сахара в крови посредством глюконеогенеза , подавляет иммунную систему и способствует метаболизму калорий . [3] Это также уменьшает образование костей. [4] Эти заявленные функции выполняются путем связывания кортизола с глюкокортикоидными или минералокортикоидными рецепторами внутри клетки, которые затем связываются с ДНК, влияя на экспрессию генов. [1] [5]
Влияние на здоровье
[ редактировать ]Метаболический ответ
[ редактировать ]Метаболизм глюкозы
[ редактировать ]Кортизол играет решающую роль в регуляции метаболизма глюкозы и способствует глюконеогенезу ( синтезу глюкозы ) и гликогенезу ( синтезу гликогена ) в печени и гликогенолизу (расщеплению гликогена ) в скелетных мышцах. [1] Он также повышает уровень глюкозы в крови за счет снижения поглощения глюкозы мышечной и жировой тканью, снижения синтеза белка и увеличения расщепления жиров на жирные кислоты (липолиз). Все эти метаболические этапы в конечном итоге приводят к повышению уровня глюкозы в крови, которая питает мозг и другие ткани во время реакции «бей или беги». Кортизол также отвечает за высвобождение аминокислот из мышц, обеспечивая субстрат для глюконеогенеза . [1] Его воздействие сложно и разнообразно. [6]
В целом кортизол стимулирует глюконеогенез (синтез «новой» глюкозы из неуглеводных источников, который происходит главным образом в печени , но также в почках и тонком кишечнике при определенных обстоятельствах ). Конечным эффектом является увеличение концентрации глюкозы в крови, что дополнительно дополняется снижением чувствительности периферических тканей к инсулину , что предотвращает получение этой тканью глюкозы из крови. Кортизол оказывает разрешающее действие на действие гормонов, повышающих выработку глюкозы, таких как глюкагон и адреналин . [7]
Кортизол также играет важную, но опосредованную роль в гликогенолизе печени и мышц (расщеплении гликогена до глюкозо-1-фосфата и глюкозы), который происходит в результате действия глюкагона и адреналина. Кроме того, кортизол способствует активации гликогенфосфорилазы , которая необходима адреналину для воздействия на гликогенолиз. [8] [9]
Парадоксально, но кортизол способствует не только глюконеогенезу (биосинтезу молекул глюкозы) в печени, но и гликогенезу ( полимеризации молекул глюкозы в гликоген ): таким образом, кортизол лучше рассматривать как стимулирующий обмен глюкозы/гликогена в печени. [10] Это контрастирует с действием кортизола на скелетные мышцы, где гликогенолиз (распад гликогена на молекулы глюкозы) стимулируется косвенно через катехоламины . [11] Таким образом, кортизол и катехоламины действуют синергически, способствуя расщеплению мышечного гликогена на глюкозу для использования в мышечной ткани. [12]
Обмен белков и липидов
[ редактировать ]Повышенный уровень кортизола, если его продлить, может привести к протеолизу (расщеплению белков) и атрофии мышц. [13] Целью протеолиза является обеспечение соответствующей ткани сырьем для глюконеогенеза; см. глюкогенные аминокислоты . [7] Влияние кортизола на липидный обмен более сложное, поскольку липогенез наблюдается у пациентов с хроническим повышенным уровнем циркулирующих глюкокортикоидов (т.е. кортизола). [7] хотя резкое увеличение циркулирующего кортизола способствует липолизу . [14] Обычное объяснение этого кажущегося несоответствия состоит в том, что повышенная концентрация глюкозы в крови (под действием кортизола) стимулирует высвобождение инсулина . Инсулин стимулирует липогенез, поэтому это является косвенным следствием повышения концентрации кортизола в крови, но это произойдет только в течение более длительного периода времени.
Иммунный ответ
[ редактировать ]Кортизол предотвращает выброс в организм веществ, вызывающих воспаление . Он используется для лечения состояний, возникающих в результате гиперактивности реакции антител, опосредованной B-клетками. Примеры включают воспалительные и ревматоидные заболевания, а также аллергии . Низкие дозы гидрокортизона для местного применения , продаваемые без рецепта в некоторых странах, используются для лечения кожных проблем, таких как сыпь и экзема .
Кортизол ингибирует выработку интерлейкина 12 (IL-12), интерферона гамма (IFN-гамма), IFN-альфа и фактора некроза опухоли альфа (TNF-альфа) антигенпрезентирующими клетками (APC) и Т-хелперными клетками (клетками Th1). , но повышает регуляцию интерлейкина 4 , интерлейкина 10 и интерлейкина 13 клетками Th2. Это приводит к сдвигу в сторону Th2-иммунного ответа, а не к общей иммуносупрессии. Считается, что активация системы стресса (и, как следствие, увеличение уровня кортизола и сдвига Th2), наблюдаемая во время инфекции, является защитным механизмом, который предотвращает чрезмерную активацию воспалительной реакции. [15]
Кортизол может ослабить активность иммунной системы . Он предотвращает пролиферацию Т-клеток, делая интерлейкина-2 -продуценты Т-клетки невосприимчивыми к интерлейкину-1 и неспособными продуцировать фактор роста Т-клеток IL-2. Кортизол подавляет экспрессию IL-2R рецептора IL2 на поверхности хелперных Т-клеток, что необходимо для индукции «клеточного» иммунного ответа Th1, тем самым способствуя сдвигу в сторону доминирования Th2 и высвобождению перечисленных выше цитокинов, что приводит к в доминировании Th2 и способствует «гуморальному» иммунному ответу антител, опосредованному B-клетками. [16]
Кортизол также оказывает эффект отрицательной обратной связи на IL-1. [17] Принцип действия этой отрицательной обратной связи заключается в том, что иммунный стрессор заставляет периферические иммунные клетки высвобождать IL-1 и другие цитокины, такие как IL-6 и TNF-альфа. Эти цитокины стимулируют гипоталамус, заставляя его высвобождать кортикотропин-рилизинг гормон (CRH). КРГ, в свою очередь, стимулирует выработку адренокортикотропного гормона (АКТГ), среди прочего, в надпочечниках, что (помимо прочего) увеличивает выработку кортизола. Затем кортизол замыкает петлю, поскольку он ингибирует выработку TNF-альфа в иммунных клетках и делает их менее чувствительными к IL-1. [18]
Благодаря этой системе, пока иммунный стрессор невелик, реакция будет регулироваться до нужного уровня. Подобно термостату, управляющему обогревателем, гипоталамус использует кортизол для выключения тепла, как только выработка кортизола соответствует стрессу, вызываемому иммунной системой. Но при тяжелой инфекции или в ситуации, когда иммунная система чрезмерно чувствительна к антигену (например, при аллергических реакциях ) или наблюдается массовый поток антигенов (как это может случиться с эндотоксичными бактериями), правильная заданная точка может никогда не быть достигнута. . Кроме того, из-за подавления иммунитета Th1 кортизолом и другими сигнальными молекулами некоторые типы инфекций (особенно микобактерии туберкулеза ) могут заставить организм застрять в неправильном режиме атаки, используя опосредованный антителами гуморальный ответ, когда клеточный ответ является неправильным. нужный.
Лимфоциты включают В-клеточные лимфоциты, которые являются клетками организма, продуцирующими антитела, и, таким образом, являются основными агентами гуморального иммунитета . Увеличение количества лимфоцитов в лимфатических узлах, костном мозге и коже означает, что в организме усиливается гуморальный иммунный ответ. В-клеточные лимфоциты выделяют антитела в кровоток. Эти антитела снижают инфекцию тремя основными путями: нейтрализацией, опсонизацией и активацией комплемента . Антитела нейтрализуют патогены, связываясь с поверхностными белками, предотвращая связывание патогенов с клетками-хозяевами. При опсонизации антитела связываются с патогеном и создают мишень для фагоцитарных иммунных клеток, которые они могут найти и закрепиться, что позволяет им легче уничтожить патоген. Наконец, антитела также могут активировать молекулы комплемента, которые могут объединяться различными способами, способствуя опсонизации, или даже действовать непосредственно, лизируя бактерии. Существует много различных типов антител, и их производство очень сложное, в нем участвуют несколько типов лимфоцитов, но в целом лимфоциты и другие клетки, регулирующие и продуцирующие антитела, мигрируют в лимфатические узлы, чтобы способствовать высвобождению этих антител в кровоток. [19]
Быстрое введение кортикостерона (эндогенного агониста рецепторов типа I и типа II) или RU28362 (специфического агониста рецепторов типа II) адреналэктомированным животным вызывало изменения в распределении лейкоцитов.
С другой стороны, существуют естественные клетки-киллеры ; эти клетки обладают способностью уничтожать более крупные угрозы, такие как бактерии, паразиты и опухолевые клетки. Отдельное исследование [20] обнаружили, что кортизол эффективно обезоруживает естественные клетки-киллеры, подавляя экспрессию их естественных рецепторов цитотоксичности. Интересно, что пролактин имеет противоположный эффект. Он увеличивает экспрессию рецепторов цитотоксичности на естественных клетках-киллерах, увеличивая их огневую мощь. [ нужна ссылка ]
Кортизол стимулирует многие ферменты меди (часто до 50% от их общего потенциала), включая лизилоксидазу , фермент, который сшивает коллаген и эластин . Особенно ценным для иммунного ответа является стимуляция кортизолом супероксиддисмутазы . [21] поскольку этот медный фермент почти наверняка используется организмом, чтобы позволить супероксидам отравлять бактерии.
Известно, что некоторые вирусы, такие как грипп и SARS-CoV-1 и SARS-CoV-2 , подавляют секрецию гормонов стресса, чтобы избежать иммунного ответа организма, тем самым избегая иммунной защиты организма. Эти вирусы подавляют кортизол, производя белок, который имитирует человеческий гормон АКТГ, но является неполным и не обладает гормональной активностью. АКТГ — это гормон, который стимулирует надпочечники вырабатывать кортизол и другие стероидные гормоны. Однако против этого вирусного белка организм вырабатывает антитела, которые также убивают человеческий гормон АКТГ, что приводит к подавлению функции надпочечников. Такое подавление надпочечников является для вируса способом избежать иммунного обнаружения и устранения. [22] [23] [24] Эта вирусная стратегия может иметь серьезные последствия для хозяина (человека, инфицированного вирусом), поскольку кортизол необходим для регулирования различных физиологических процессов, таких как обмен веществ, кровяное давление, воспаление и иммунный ответ. Недостаток кортизола может привести к состоянию, называемому недостаточностью надпочечников, которое может вызывать такие симптомы, как усталость, потеря веса, низкое кровяное давление, тошнота, рвота и боль в животе. Надпочечниковая недостаточность также может ухудшить способность хозяина справляться со стрессом и инфекциями, поскольку кортизол помогает мобилизовать источники энергии, увеличивать частоту сердечных сокращений и подавлять несущественные метаболические процессы во время стресса. Таким образом, подавляя выработку кортизола, некоторые вирусы могут выйти за пределы иммунной системы и ослабить общее состояние здоровья и устойчивость хозяина. [25] [23] [24]
Другие эффекты
[ редактировать ]Метаболизм
[ редактировать ]Глюкоза
[ редактировать ]Кортизол противодействует инсулину , способствует гипергликемии путем стимуляции глюконеогенеза и подавляет периферическое использование глюкозы ( резистентность к инсулину ). [26] за счет уменьшения транслокации переносчиков глюкозы (особенно GLUT4 ) на клеточную мембрану. [1] [27] Кортизол также увеличивает синтез гликогена (гликогенез) в печени, сохраняя глюкозу в легкодоступной форме. [28]
Кость и коллаген
[ редактировать ]Кортизол снижает костеобразование, [4] благоприятствуя долгосрочному развитию остеопороза (прогрессирующего заболевания костей). Механизм, лежащий в основе этого, двоякий: кортизол стимулирует выработку RANKL остеобластами , которые посредством связывания с RANK -рецепторами стимулируют активность остеокластов – клеток, ответственных за резорбцию кальция из кости, – а также ингибирует выработку остеопротегерина (OPG), который действует как рецептор-ловушка и захватывает некоторое количество RANKL, прежде чем он сможет активировать остеокласты посредством RANK. [7] Другими словами, когда RANKL связывается с OPG, ответа не происходит, в отличие от связывания с RANK, которое приводит к активации остеокластов.
Он выводит калий из клеток в обмен на такое же количество ионов натрия (см. выше). [29] Это может спровоцировать гиперкалиемию метаболического шока после операции. Кортизол также снижает всасывание кальция в кишечнике. [30] Кортизол подавляет синтез коллагена . [31]
Аминокислота
[ редактировать ]Кортизол повышает уровень свободных аминокислот в сыворотке, ингибируя образование коллагена, уменьшая поглощение аминокислот мышцами и ингибируя синтез белка. [32] Кортизол (как и оптикортинол) может обратно ингибировать клетки-предшественники IgA в кишечнике телят. [33] Кортизол также ингибирует IgA в сыворотке, как и IgM ; однако не показано, что он ингибирует IgE . [34]
Электролитный баланс
[ редактировать ]Кортизол увеличивает скорость клубочковой фильтрации, [35] и почечный поток плазмы из почек, что увеличивает экскрецию фосфатов, [36] [37] а также увеличение задержки натрия и воды и выведение калия путем воздействия на минералокортикоидные рецепторы . Он также увеличивает всасывание натрия и воды и выведение калия в кишечнике. [38]
Натрий
[ редактировать ]Кортизол способствует всасыванию натрия через тонкий кишечник млекопитающих. [39] Однако истощение натрия не влияет на уровень кортизола. [40] поэтому кортизол нельзя использовать для регулирования натрия в сыворотке. Первоначальной целью кортизола, возможно, был транспорт натрия. Эта гипотеза подтверждается тем фактом, что пресноводные рыбы используют кортизол для стимуляции поступления натрия внутрь, в то время как морские рыбы имеют систему, основанную на кортизоле, для выведения избытка натрия. [41]
Калий
[ редактировать ]Натриевая нагрузка усиливает интенсивное выведение калия кортизолом. В этом случае кортикостерон сравним с кортизолом. [42] Чтобы калий вышел из клетки, кортизол перемещает в клетку равное количество ионов натрия. [29] Это должно значительно облегчить регулирование pH (в отличие от обычной ситуации дефицита калия, в которой на каждые три иона калия приходят два иона натрия — ближе к эффекту дезоксикортикостерона ).
Желудок и почки
[ редактировать ]Кортизол стимулирует секрецию желудочной кислоты. [43] Единственное прямое воздействие кортизола на выведение ионов водорода почками заключается в стимуляции выведения ионов аммония путем дезактивации фермента почечной глутаминазы. [44]
Память
[ редактировать ]Кортизол работает с адреналином (адреналином), создавая воспоминания о кратковременных эмоциональных событиях; Это предлагаемый механизм хранения воспоминаний-вспышек , который может возникнуть как средство запоминания того, чего следует избегать в будущем. [45] Однако длительное воздействие кортизола повреждает клетки гиппокампа ; [46] этот ущерб приводит к нарушению обучения.
Суточные циклы
[ редактировать ]
суточные циклы уровня кортизола. У человека наблюдаются [8]
Стресс
[ редактировать ]Длительный стресс может привести к повышению уровня циркулирующего кортизола (который считается одним из наиболее важных из нескольких «гормонов стресса»). [47]
Эффекты во время беременности
[ редактировать ]Во время беременности у человека повышенное производство кортизола плодом между 30 и 32 неделями инициирует выработку сурфактанта легких плода , способствующего созреванию легких. У плодов ягнят уровень глюкокортикоидов (главным образом кортизола) повышается примерно после 130-го дня, при этом уровень поверхностно-активного вещества в легких в ответ значительно увеличивается примерно к 135-му дню. [48] и хотя кортизол плода ягненка в основном имеет материнское происхождение в течение первых 122 дней, 88% или более имеет эмбриональное происхождение к 136-му дню беременности. [49] Хотя время повышения концентрации кортизола у плода у овец может несколько различаться, в среднем оно составляет около 11,8 дней до начала родов. [50] У некоторых видов домашнего скота (например, крупного рогатого скота, овец, коз и свиней) выброс фетального кортизола на поздних сроках беременности вызывает начало родов, устраняя прогестероновый блок расширения шейки матки и сокращения миометрия . Механизмы, оказывающие этот эффект на прогестерон, различаются у разных видов. У овец, у которых прогестерон, достаточный для поддержания беременности, вырабатывается плацентой примерно после 70-го дня беременности, [51] [52] Всплеск кортизола плода перед родами индуцирует плацентарное ферментативное превращение прогестерона в эстроген. (Повышенный уровень эстрогена стимулирует секрецию простагландинов и развитие рецепторов окситоцина .)
Воздействие кортизола на плод во время беременности может иметь различные последствия для развития, включая изменения в пренатальных и постнатальных моделях роста. У мартышек , вида приматов Нового Света, беременные самки имеют разные уровни кортизола во время беременности, как внутри самок, так и между ними. Младенцы, рожденные от матерей с высоким уровнем гестационного кортизола в первом триместре беременности, имели более низкие темпы роста индекса массы тела, чем младенцы, рожденные от матерей с низким уровнем гестационного кортизола (примерно на 20% ниже). Однако темпы постнатального роста у этих детей с высоким уровнем кортизола были более быстрыми, чем у детей с низким уровнем кортизола в более поздние постнатальные периоды, и полное догоняющее развитие произошло к 540-дневному возрасту. Эти результаты позволяют предположить, что воздействие кортизола на плод во время беременности оказывает важное потенциальное влияние на программирование плода как на пре-, так и на постнатальный рост приматов. [53]
Синтез и выпуск
[ редактировать ]Кортизол вырабатывается в организме человека пучковой вторым оболочкой надпочечников , из трех слоев коры надпочечников . [1] Эта кора образует внешнюю «кору» каждого надпочечника, расположенную над почками. Выброс кортизола контролируется гипоталамусом головного мозга. Секреция кортикотропин-рилизинг гормона гипоталамусом заставляет клетки соседней передней доли гипофиза секретировать адренокортикотропный гормон (АКТГ) в сосудистую систему, через которую кровь переносит его в кору надпочечников. [1] АКТГ стимулирует синтез кортизола и других глюкокортикоидов, минералокортикоида альдостерона и дегидроэпиандростерона . [1]
Тестирование физических лиц
[ редактировать ]Нормальные значения, указанные в следующих таблицах, относятся к людям (нормальные уровни различаются у разных видов). Измеренные уровни кортизола и, следовательно, референтные диапазоны зависят от типа образца, используемого аналитического метода и таких факторов, как возраст и пол. Поэтому результаты испытаний всегда следует интерпретировать с использованием референтного диапазона лаборатории, выдавшей результат. [54] [55] [56] Уровни кортизола человека можно определить в крови, сыворотке, моче, слюне и поте. [57]
| Время | Нижний предел | Верхний предел | Единица |
|---|---|---|---|
| 09:00 утра | 140 [58] [59] | 700 [58] | нмоль/л |
| 5 [60] | 25 [60] | мкг/дл | |
| Полночь | 80 [58] | 350 [58] | нмоль/л |
| 2.9 [60] | 13 [60] | мкг/дл |
При молекулярной массе 362,460 г/моль коэффициент перевода мкг/дл в нмоль/л составляет примерно 27,6; [61] [62] таким образом, 10 мкг/дл составляет около 276 нмоль/л.
| Нижний предел | Верхний предел | Единица |
|---|---|---|
| 28 [63] или 30 [64] | 280 [63] или 490 [64] | нмоль /24 часа |
| 10 [65] или 11 [66] | 100 [65] или 176 [66] | мкг /24 ч |
Кортизол подчиняется циркадному ритму , и для точного измерения уровня кортизола лучше всего проводить тестирование четыре раза в день через слюну. У человека может быть нормальный общий уровень кортизола, но его уровень ниже нормального в течение определенного периода дня и выше нормального в другой период. Поэтому некоторые ученые ставят под сомнение клиническую полезность измерения кортизола. [67] [68] [69] [70]
Кортизол липофильен и транспортируется в связанном виде с транскортином (также известным как кортикостероидсвязывающий глобулин (КБГ)) и альбумином , тогда как лишь небольшая часть общего сывороточного кортизола несвязана и обладает биологической активностью. [71] Это связывание кортизола с транскортином осуществляется посредством гидрофобных взаимодействий, при которых кортизол связывается в соотношении 1:1. [72] Анализы на кортизол в сыворотке измеряют общий уровень кортизола, и его результаты могут вводить в заблуждение пациентов с измененной концентрацией белка в сыворотке. Тест на кортизол в слюне позволяет избежать этой проблемы, поскольку только свободный кортизол может пройти через гемато-слюнный барьер . [73] [74] [75] [76] Частицы транскортина слишком велики, чтобы пройти через этот барьер. [77] Состоит из слоев эпителиальных клеток слизистой оболочки полости рта и слюнных желез. [78]
Кортизол может попасть в волосы из крови, пота и кожного сала . Сегмент волос на голове длиной 3 сантиметра может соответствовать 3 месяцам роста волос, хотя скорость роста может варьироваться в разных участках кожи головы. Кортизол в волосах является надежным индикатором хронического воздействия кортизола. [79]
Автоматизированным иммуноанализам не хватает специфичности, они демонстрируют значительную перекрестную реактивность из-за взаимодействия со структурными аналогами кортизола, а также демонстрируют различия между анализами. Жидкостная хроматография-тандемная масс-спектрометрия (ЖХ-МС/МС) может улучшить специфичность и чувствительность. [80]
Нарушения выработки кортизола
[ редактировать ]Некоторые медицинские расстройства связаны с аномальной выработкой кортизола, например:
- Первичный гиперкортизолизм ( синдром Кушинга ): чрезмерный уровень кортизола. [81]
- Вторичный гиперкортицизм (опухоль гипофиза, приводящая к болезни Кушинга , [82] [83] синдром псевдо-Кушинга ).
- Первичный гипокортизолизм ( болезнь Аддисона , синдром Нельсона ): недостаточный уровень кортизола.
- Вторичный гипокортизолизм (опухоль гипофиза, синдром Шихана )
Регулирование
[ редактировать ]Первичным контролем кортизола является пептид гипофиза АКТГ, который, вероятно, контролирует кортизол, контролируя движение кальция в клетки-мишени, секретирующие кортизол. [84] АКТГ, в свою очередь, контролируется гипоталамическим пептидом кортикотропин-рилизинг-гормоном (CRH), который находится под нервным контролем. КРГ действует синергично с аргинином, вазопрессином , ангиотензином II и адреналином . [85] (У свиней, которые не производят аргинин-вазопрессин, лизин-вазопрессин действует синергически с КРГ. [86] )
Когда активированные макрофаги начинают секретировать IL-1, который синергически с CRH увеличивает АКТГ, [17] Т-клетки также секретируют фактор, модифицирующий глюкостероидный ответ (GRMF), а также IL-1; оба увеличивают количество кортизола, необходимого для подавления почти всех иммунных клеток. [87] Затем иммунные клетки принимают на себя собственную регуляцию, но с более высоким уровнем кортизола. Однако повышение уровня кортизола у телят с диареей минимально по сравнению со здоровыми телятами и со временем падает. [88] Клетки не теряют своей функции «бей или беги» из-за синергизма интерлейкина-1 с CRH. Кортизол даже оказывает отрицательный эффект обратной связи на интерлейкин-1. [17] — особенно полезно для лечения заболеваний, которые заставляют гипоталамус выделять слишком много CRH, например, вызванных эндотоксическими бактериями. GRMF не влияет на иммунные клетки-супрессоры. [87] поэтому эффективная уставка иммунных клеток может быть даже выше, чем уставка физиологических процессов. GRMF влияет в первую очередь на печень (а не на почки) при некоторых физиологических процессах. [89]
Среды с высоким содержанием калия (которые стимулируют секрецию альдостерона in vitro ) также стимулируют секрецию кортизола из пучковой зоны надпочечников собак. [90] [91] — в отличие от кортикостерона, на который калий не влияет. [92]
Нагрузка калием также увеличивает АКТГ и кортизол у людей. [93] Вероятно, это причина того, что дефицит калия приводит к снижению кортизола (как уже упоминалось) и вызывает уменьшение превращения 11-дезоксикортизола в кортизол. [94] Это также может иметь значение при боли при ревматоидном артрите; клеточный калий при РА всегда низкий. [95]
Также было показано, что присутствие аскорбиновой кислоты, особенно в высоких дозах, опосредует реакцию на психологический стресс и ускоряет снижение уровня циркулирующего кортизола в организме после стресса. Об этом можно судить по снижению систолического и диастолического артериального давления и снижению уровня кортизола в слюне после лечения аскорбиновой кислотой. [96]
Факторы, повышающие уровень кортизола
[ редактировать ]- Вирусные инфекции повышают уровень кортизола за счет активации оси HPA цитокинами. [97]
- Интенсивные (высокое VO 2 max ) или продолжительные аэробные упражнения временно повышают уровень кортизола, усиливая глюконеогенез и поддерживая уровень глюкозы в крови; [98] однако уровень кортизола снижается до нормального уровня после еды (т. е. восстанавливается нейтральный энергетический баланс ). [99]
- Тяжелая травма или стрессовые события могут повысить уровень кортизола в крови на длительный период времени. [100]
- Низкоуглеводные диеты вызывают кратковременное повышение уровня кортизола в состоянии покоя (≈3 недели) и усиливают реакцию кортизола на аэробные упражнения в краткосрочной и долгосрочной перспективе. [101]
- Увеличение концентрации грелина, гормона, стимулирующего голод, повышает уровень кортизола. [102]
Биохимия
[ редактировать ]Биосинтез
[ редактировать ]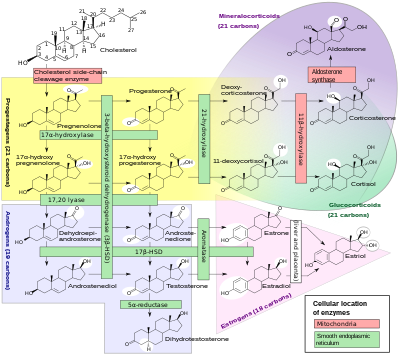
Кортизол синтезируется из холестерина . Синтез происходит в пучковой зоне коры надпочечников . [104] [105] [106]
Название «кортизол» происходит от слова «кора». Кортекс означает «внешний слой» — ссылка на кору надпочечников, часть надпочечников, где вырабатывается кортизол. [107]
В то время как кора надпочечников у людей также вырабатывает альдостерон в клубочковой зоне и некоторые половые гормоны в сетчатой зоне , у людей и некоторых других видов основным секретом является кортизол. [105] У крупного рогатого скота уровень кортикостерона может приближаться к [108] или превысить [8] уровень кортизола. [109] [110] У человека мозговое вещество надпочечников лежит под его корой, секретируя в основном катехоламины адреналин (адреналин) и норадреналин (норадреналин) под симпатической стимуляцией. [111]
надпочечниках стимулируется передней долей гипофиза Синтез кортизола в АКТГ; Производство АКТГ, в свою очередь, стимулируется КРГ, который высвобождается гипоталамусом. АКТГ увеличивает концентрацию холестерина во внутренней мембране митохондрий посредством регуляции стероидогенного острого регуляторного белка. Он также стимулирует основной лимитирующий этап синтеза кортизола, на котором холестерин превращается в прегненолон и катализируется цитохромом P450SCC ( ферментом расщепления боковой цепи ). [112]
Метаболизм
[ редактировать ]11бета-гидроксистероиддегидрогеназы
[ редактировать ]Кортизол обратимо метаболизируется до кортизона. [113] системой 11-бета-гидроксистероиддегидрогеназы (11-бета-HSD), которая состоит из двух ферментов: 11-бета-HSD1 и 11-бета-HSD2 . Метаболизм кортизола в кортизон включает окисление гидроксильной группы в положении 11-бета. [114]
- 11-бета HSD1 использует кофактор НАДФН для преобразования биологически инертного кортизона в биологически активный кортизол.
- 11-бета HSD2 использует кофактор НАД+ для преобразования кортизола в кортизон.
В целом, конечный эффект заключается в том, что 11-бета-HSD1 способствует увеличению локальных концентраций биологически активного кортизола в данной ткани; 11-бета-HSD2 служит для снижения локальных концентраций биологически активного кортизола. Если присутствует гексозо-6-фосфатдегидрогеназа (H6PDH), равновесие может способствовать активности 11-бета-HSD1. H6PDH регенерирует НАДФН, что увеличивает активность 11-бета-HSD1 и снижает активность 11-бета-HSD2. [115]
Было высказано предположение, что изменение 11-бета-HSD1 играет роль в патогенезе ожирения резистентности к , гипертонии и инсулину , известного как метаболический синдром . [116]
Изменение 11-бета-HSD2 связано с эссенциальной гипертензией и, как известно, приводит к синдрому кажущегося избытка минералокортикоидов (SAME).
А-кольцевые редуктазы (5альфа- и 5бета-редуктазы)
[ редактировать ]Кортизол также необратимо метаболизируется в 5-альфа-тетрагидрокортизол (5-альфа-ТГФ) и 5-бета-тетрагидрокортизол (5-бета-ТГФ), реакции, для которых являются 5-альфа-редуктаза и 5-бета-редуктаза лимитирующими факторами соответственно. 5-бета-редуктаза также является фактором, ограничивающим скорость превращения кортизона в тетрагидрокортизон . [ нужна медицинская ссылка ]
Цитохром P450, семейство 3, подсемейство А монооксигеназы
[ редактировать ]Кортизол также необратимо метаболизируется в 6β-гидроксикортизол монооксигеназами цитохрома p450-3A, главным образом, CYP3A4 . [117] [118] [113] [119] Препараты, индуцирующие CYP3A4, могут ускорять клиренс кортизола. [120]
Химия
[ редактировать ]Кортизол представляет собой природный прегнановый кортикостероид , также известный как 11β,17α,21-тригидроксипрегн-4-ен-3,20-дион .
Животные
[ редактировать ]У животных кортизол часто используется как индикатор стресса, и его можно измерить в крови. [121] слюна, [121] моча, [122] волосы, [123] и фекалии. [123] [124]
См. также
[ редактировать ]- Кортизон , гормон
- Реакция пробуждения кортизола
- Список кортикостероидов
- Мембранный глюкокортикоидный рецептор
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я Лайтман С.Л., Бирни М.Т., Конвей-Кэмпбелл Б.Л. (июнь 2020 г.). «Динамика секреции АКТГ и кортизола и последствия для болезней» . Эндокринные обзоры . 41 (3). дои : 10.1210/endrev/bnaa002 . ПМК 7240781 . ПМИД 32060528 .
- ^ Тейвес, доктор медицинских наук, Гомес-Санчес CE, Сома К.К. (июль 2011 г.). «Вненадпочечниковые глюкокортикоиды и минералокортикоиды: данные о местном синтезе, регуляции и функции» . Американский журнал физиологии. Эндокринология и обмен веществ . 301 (1): Е11-24. дои : 10.1152/ajpendo.00100.2011 . ПМК 3275156 . ПМИД 21540450 .
- ^ Хен К., Мариб Э.Н. (2010). Анатомия и физиология человека . Сан-Франциско: Бенджамин Каммингс. ISBN 978-0-321-60261-9 .
- ^ Jump up to: а б Чюн Ю.С., Крем Б.Е., Раис Л.Г. (февраль 1984 г.). «Кортизол уменьшает образование костей, подавляя пролиферацию периостальных клеток». Эндокринология . 114 (2): 477–80. дои : 10.1210/эндо-114-2-477 . ПМИД 6690287 .
- ^ ДеРейк Р.Х., Шааф М., де Клоет Э.Р. (июнь 2002 г.). «Варианты глюкокортикоидных рецепторов: клиническое значение». Журнал биохимии стероидов и молекулярной биологии . 81 (2): 103–122. дои : 10.1016/S0960-0760(02)00062-6 . ПМИД 12137800 . S2CID 24650907 .
- ^ Хани С., Тайек Дж.А. (декабрь 2001 г.). «Кортизол усиливает глюконеогенез у людей: его роль в метаболическом синдроме» (PDF) . Клин Сай (Лондон) . 101 (6): 739–47. дои : 10.1042/cs1010739 . ПМИД 11724664 . Архивировано из оригинала 11 марта 2024 года . Проверено 2 февраля 2024 г.
- ^ Jump up to: а б с д Лэйкок Дж. Ф. (2013). Интегрированная эндокринология . Миран, Карим. Чичестер, Западный Суссекс, Великобритания: Уайли-Блэквелл. ISBN 978-1-118-45064-2 . OCLC 794973804 .
- ^ Jump up to: а б с Мартин П.А., Крамп М.Х. (2003). «Надпочечники». В Дули М.П., Пинеда М.Х. (ред.). Ветеринарная эндокринология и репродукция Макдональдса (5-е изд.). Эймс, Айова: Издательство штата Айова. ISBN 978-0-8138-1106-2 .
- ^ Кодер Л., Шривастава А.К., Чиассон Дж.Л. (июнь 1991 г.). «Роль глюкокортикоидов в регуляции метаболизма гликогена в скелетных мышцах». Американский журнал физиологии . 260 (6 Ч. 1): E927–32. дои : 10.1152/ajpendo.1991.260.6.E927 . ПМИД 1905485 .
- ^ Макфарлейн Д.П., Форбс С., Уокер Б.Р. (май 2008 г.). «Глюкокортикоиды и метаболизм жирных кислот у человека: стимулирование перераспределения жира при метаболическом синдроме» . Журнал эндокринологии . 197 (2): 189–204. дои : 10.1677/JOE-08-0054 . ПМИД 18434349 .
- ^ Куо Т., МакКуин А., Чен Т.К., Ван Дж.К. (2015). «Регуляция гомеостаза глюкозы глюкокортикоидами». В Ван Дж.К., Харрис С. (ред.). Глюкокортикоидная сигнализация . Достижения экспериментальной медицины и биологии. Том. 872. Спрингер. стр. 99–126. дои : 10.1007/978-1-4939-2895-8_5 . ISBN 978-1-4939-2895-8 . ПМК 6185996 . ПМИД 26215992 .
- ^ Слюйтер Дж.К., Фрингс-Дрезен М.Х., Мейман Т.Ф., ван дер Бик А.Дж. (май 2000 г.). «Реактивность и восстановление после различных видов работы, измеряемые катехоламинами и кортизолом: систематический обзор литературы» . Оккуп Энвирон Мед . 57 (5): 298–315. дои : 10.1136/oem.57.5.298 . ПМЦ 1739955 . ПМИД 10769296 .
- ^ Симмонс П.С., Майлз Дж.М., Герих Дж.Э., Хеймонд М.В. (февраль 1984 г.). «Увеличенный протеолиз. Эффект увеличения уровня кортизола в плазме в пределах физиологического диапазона» . Журнал клинических исследований . 73 (2): 412–20. дои : 10.1172/JCI111227 . ПМК 425032 . ПМИД 6365973 .
- ^ Джурхуус CB, Гравхольт СН, Нильсен С, Менгель А, Кристиансен ЙС, Шмитц ОЕ и др. (июль 2002 г.). «Влияние кортизола на липолиз и региональные уровни интерстициального глицерина у людей». Американский журнал физиологии. Эндокринология и обмен веществ . 283 (1): E172–7. дои : 10.1152/ajpendo.00544.2001 . ПМИД 12067858 . S2CID 2609285 .
- ^ Еленков И.Ю. (июнь 2004 г.). «Глюкокортикоиды и баланс Th1/Th2» . Анналы Нью-Йоркской академии наук . 1024 (1): 138–46. Бибкод : 2004NYASA1024..138E . дои : 10.1196/анналы.1321.010 . PMID 15265778 . S2CID 9575617 . Архивировано из оригинала 11 декабря 2022 года . Проверено 11 сентября 2018 г.
- ^ Паласиос Р., Сугавара I (январь 1982 г.). «Гидрокортизон подавляет пролиферацию Т-клеток в реакции аутологичных смешанных лимфоцитов, делая Т-клетки-продуценты интерлейкина-2 нечувствительными к интерлейкину-1 и неспособными синтезировать фактор роста Т-клеток». Скандинавский журнал иммунологии . 15 (1): 25–31. дои : 10.1111/j.1365-3083.1982.tb00618.x . ПМИД 6461917 . S2CID 41292936 .
- ^ Jump up to: а б с Беседовский Х.О., Дель Рей А., Соркин Э. (1986). «Интеграция активированных продуктов иммунных клеток в цепи иммунной эндокринной обратной связи». В Оппенгейме JJ, Джейкобс DM (ред.). Лейкоциты и защита хозяина . Прогресс в биологии лейкоцитов. Том. 5. Нью-Йорк: Алан Р. Лисс. п. 200.
- ^ Демерс Лоуренс М (2008). «Нарушения коры надпочечников». В Бертисе Карле А., Эшвуде Эдварде Р., Брунсе Дэвиде Э., Сойере, Барбаре Дж. (ред.). Титц «Основы клинической химии» . Сент-Луис, Миссури: Сондерс Эль-Севьер. стр. 749–765.
- ^ Мерфи, Кеннет (2012). «Гуморальный иммунный ответ». Иммунобиология Джейнвей, 8-е изд . Нью-Йорк, штат Нью-Йорк: Garland Science Taylor & Francisco Group. п. 387.
- ^ Мавунгу Э., Бую-Акотет М.К., Кремснер П.Г. (февраль 2005 г.). «Влияние пролактина и кортизола на экспрессию поверхности естественных клеток-киллеров (NK) и функцию естественных рецепторов цитотоксичности человека (NKp46, NKp44 и NKp30)» . Клиническая и экспериментальная иммунология . 139 (2): 287–96. дои : 10.1111/j.1365-2249.2004.02686.x . ПМК 1809301 . ПМИД 15654827 .
- ^ Флоэ Л., Бекман Р., Гирц Х., Лошен Г. (1985). «Кислородцентрированные свободные радикалы как медиаторы воспаления». В Сиесе Х (ред.). Окислительный стресс . Лондон: Орландо. п. 405. ИСБН 978-0-12-642760-8 .
- ^ Антони М.Х., Дхабхар Ф.С. (2019). «Влияние психосоциального стресса и управления стрессом на иммунные реакции у больных раком» . Рак . 125 (9): 1417–1431. дои : 10.1002/cncr.31943 . ПМК 6467795 . PMID 30768779 .
- ^ Jump up to: а б Пал Р. (май 2020 г.). «COVID-19, гипоталамо-гипофизарно-надпочечниковая ось и клинические последствия» . Эндокринный . 68 (2): 251–252. дои : 10.1007/s12020-020-02325-1 . ПМЦ 7186765 . ПМИД 32346813 .
- ^ Jump up to: а б Акбас Э.М., Акбас Н. (март 2021 г.). «COVID-19, надпочечники, глюкокортикоиды и надпочечниковая недостаточность» . Biomed Pap Med Fac Univ Palacky Olomouc Чешская Республика . 165 (1): 1–7. дои : 10.5507/bp.2021.011 . ПМИД 33542545 .
- ^ Буфорд Т.В., Уиллоуби Д.С. (июнь 2008 г.). «Влияние DHEA(S) и кортизола на иммунную функцию при старении: краткий обзор». Appl Physiol Nutr Metab . 33 (3): 429–33. дои : 10.1139/H08-013 . ПМИД 18461094 .
- ^ Шернтанер-Рейтер М.Х., Вольф П., Вила Г., Люгер А. (2021 г.). «Взаимодействие синдромов инсулина и гормонов гипофиза» . Фронт Эндокринол (Лозанна) . 12 : 626427. дои : 10.3389/fendo.2021.626427 . ПМЦ 8113952 . ПМИД 33995272 .
- ^ Король МБ (2005). Ланге: вопросы и ответы . Нью-Йорк: МакГроу-Хилл, Медицинский паб. Разделение. ISBN 978-0-07-144578-8 .
- ^ Бэйнс Дж., Доминичак М. (2009). Медицинская биохимия . Мосби Эльзевир. ISBN 978-0-323-05371-6 .
- ^ Jump up to: а б Найт Р.П., Корнфельд Д.С., Глейзер Г.Х., Бонди П.К. (февраль 1955 г.). «Влияние внутривенного гидрокортизона на электролиты сыворотки и мочи у человека». Журнал клинической эндокринологии и метаболизма . 15 (2): 176–81. doi : 10.1210/jcem-15-2-176 . ПМИД 13233328 .
- ^ Немецкий E (апрель 1978 г.). «[Патогенез тромбоцитопении. 2. Нарушения распределения, псевдотромбоцитопении]». Достижения медицины (на немецком языке). 96 (14): 761–2. ПМИД 346457 .
- ^ Кучарз Э.Дж. (1988). «Гормональный контроль метаболизма коллагена. Часть II». Эндокринология . 26 (4): 229–37. ПМИД 3062759 .
- ^ Манчестер, КЛ (1964). «Места гормональной регуляции белкового обмена». В Эллисон, Нью-Хэмпшир, Манро Дж.Б. (ред.). Белковый обмен млекопитающих . Нью-Йорк: Академическая пресса. п. 229? 273?.
- ^ Муж Эй Джей, Брэндон М.Р., Ласселлес А.К. (октябрь 1973 г.). «Влияние кортикостероидов на всасывание и эндогенную продукцию иммуноглобулинов у телят». Австралийский журнал экспериментальной биологии и медицинской науки . 51 (5): 707–10. дои : 10.1038/icb.1973.67 . ПМИД 4207041 .
- ^ Поузи WC, Нельсон HS, Бранч B, Перлман DS (декабрь 1978 г.). «Влияние острой кортикостероидной терапии астмы на уровень иммуноглобулина в сыворотке». Журнал аллергии и клинической иммунологии . 62 (6): 340–8. дои : 10.1016/0091-6749(78)90134-3 . ПМИД 712020 .
- ^ Ли X, Сян X, Ху Дж., Госвами Р., Ян С., Чжан А. и др. (2016). «Связь между кортизолом сыворотки и хронической болезнью почек у пациентов с эссенциальной гипертонией» . Давление крови в почках . 41 (4): 384–91. дои : 10.1159/000443435 . ПМИД 27344357 . S2CID 8633049 .
- ^ Хилл К.Дж., Ламберс ER, Эльбурн I (февраль 1988 г.). «Действие кортизола на функцию почек плода». Дж. Дев Физиол . 10 (1): 85–96. ПМИД 3351211 .
- ^ Бибер Дж., Мурер Х., Мохебби Н., Вагнер Калифорния (апрель 2014 г.). «Обращение с фосфатами и сульфатами в почках» (PDF) . Компр Физиол . 4 (2): 771–92. дои : 10.1002/cphy.c120031 . ISBN 9780470650714 . ПМИД 24715567 . Архивировано (PDF) из оригинала 1 ноября 2023 года . Проверено 16 сентября 2023 г.
- ^ Маккей Л.И., Цидловски Дж.А. (2003). «Физиологические и фармакологические эффекты кортикостероидов» . В Куре Д.В., Поллок Р.Э., Вайхзельбаум Р.Р., Баст Р.К., Ганглиер Т.С., Холланд Дж.Ф., Фрей Э. (ред.). Медицина рака Холланд-Фрай (6-е изд.). Гамильтон, Онтарио: Декер. ISBN 978-1-55009-213-4 .
- ^ Сэндл Дж.И., Кейр М.Дж., Record CO (1981). «Влияние гидрокортизона на транспорт воды, натрия и глюкозы в тощей кишке. Перфузионные исследования у нормальных субъектов и пациентов с целиакией». Скандинавский журнал гастроэнтерологии . 16 (5): 667–71. дои : 10.3109/00365528109182028 . ПМИД 7323700 .
- ^ Мейсон П.А., Фрейзер Р., Мортон Дж.Дж., Семпл П.Ф., Уилсон А. (август 1977 г.). «Влияние депривации натрия и инфузии ангиотензина II на концентрации 18-гидроксикортикостерона, альдостерона и других кортикостероидов в периферической плазме у человека». Журнал биохимии стероидов . 8 (8): 799–804. дои : 10.1016/0022-4731(77)90086-3 . ПМИД 592808 .
- ^ Горбман А., Дикхофф В.В., Винья С.Р., Кларк Н.Б., Мюллер А.Ф. (1983). Сравнительная эндокринология . Нью-Йорк: Уайли. ISBN 978-0-471-06266-0 .
- ^ Мюллер А. Ф., Оконнор К. М. (1958). Международный симпозиум по альдостерону . Литтл Браун и Ко. с. 58.
- ^ Соффер Л.Дж., Дорфман Р.И., Габрилове Дж.Л. (1961). Надпочечники человека . Филадельфия: Леа и Фебигер.
- ^ Kokoshchuk GI, Pakhmurnyĭ BA (May 1979). "Role of glucocorticoids in regulating the acid-excreting function of the kidneys". Fiziologicheskii Zhurnal SSSR Imeni I. M. Sechenova . 65 (5): 751–4. PMID 110627 .
- ^ Кеннеди Р. «Кортизол (гидрокортизон)» . Медицинская библиотека врачей. Архивировано из оригинала 3 июля 2013 года . Проверено 14 июня 2013 г.
- ^ Маколи М.Т., Кенни Р.А., Кирквуд Т.Б., Уилкинсон Д.Д., Джонс Дж.Дж., Миллер В.М. (март 2009 г.). «Математическая модель дисфункции гиппокампа, связанной со старением и кортизолом» . BMC Нейронаука . 10:26 . дои : 10.1186/1471-2202-10-26 . ПМК 2680862 . ПМИД 19320982 .
- ^ Лундберг У (2010). «Нейроэндокринные мероприятия». В Contrada R, Баум А. (ред.). Справочник по науке о стрессе: биология, психология и здоровье . Нью-Йорк: Издательская компания Springer. п. 351. ИСБН 978-0-8261-1771-7 . Архивировано из оригинала 11 марта 2024 года . Проверено 12 марта 2020 г.
[...] адреналин, норадреналин и кортизол считаются наиболее важными «гормонами стресса», хотя стресс влияет и на ряд других гормонов [...].
- ^ Мешер Э.Дж., Платцкер А.С., Баллард П.Л., Киттерман Дж.А., Клементс Дж.А., Тули WH (декабрь 1975 г.). «Онтогенез трахеальной жидкости, легочного сурфактанта и кортикоидов плазмы у плода ягненка». Журнал прикладной физиологии . 39 (6): 1017–21. дои : 10.1152/яп.1975.39.6.1017 . ПМИД 2573 .
- ^ Хеннесси Д.П., Коглан Дж.П., Харди К.Дж., Скоггинс Б.А., Винтур Э.М. (октябрь 1982 г.). «Происхождение кортизола в крови плода овцы». Журнал эндокринологии . 95 (1): 71–9. дои : 10.1677/joe.0.0950071 . ПМИД 7130892 .
- ^ Мадьяр Д.М., Фридшал Д., Эльснер К.В., Глатц Т., Элиот Дж., Кляйн А.Х. и др. (июль 1980 г.). «Анализ временных тенденций концентрации кортизола в плазме у плода овцы в зависимости от родов». Эндокринология . 107 (1): 155–9. дои : 10.1210/эндо-107-1-155 . ПМИД 7379742 .
- ^ Рикеттс АП, Флинт АП (август 1980 г.). «Начало синтеза прогестерона овечьей плацентой». Журнал эндокринологии . 86 (2): 337–47. дои : 10.1677/joe.0.0860337 . ПМИД 6933207 .
- ^ Аль-Губори К.Х., Солари А., Мирман Б. (1999). «Влияние лютеэктомии на поддержание беременности, концентрацию циркулирующего прогестерона и продуктивность окота у овец». Воспроизводство, рождаемость и развитие . 11 (6): 317–22. дои : 10.1071/RD99079 . ПМИД 10972299 .
- ^ Мусто АК, Бирни АК, Корган АК, Санто ЖБ, Френч ЖА (февраль 2012 г.). «Естественные вариации гестационного кортизола связаны с особенностями роста мартышек (Callithrix geoffroyi)» . Общая и сравнительная эндокринология . 175 (3): 519–26. дои : 10.1016/j.ygcen.2011.12.020 . ПМК 3268124 . ПМИД 22212825 .
- ^ Бьянки Л., Кампи Б., Сесса М.Р., Де Марко Г., Феррарини Э., Зукки Р. и др. (ноябрь 2019 г.). «Измерение свободного кортизола в моче с помощью ЖХ-МС-МС: принятие литературного эталонного диапазона и сравнение с нашим текущим иммунометрическим методом». Джей Эндокринол Инвест . 42 (11): 1299–1305. дои : 10.1007/s40618-019-01050-5 . ПМИД 31012054 . S2CID 128361547 .
- ^ Пантон К.К., Миккельсен Г., Иргенс В.О., Ховде А.К., Киллингмо М.В., Ойен М.А. и др. (сентябрь 2019 г.). «Новые референтные интервалы для кортизола, кортизол-связывающего глобулина и индекса свободного кортизола у женщин, использующих этинилэстрадиол» . Scand J Clin Lab Invest . 79 (5): 314–319. дои : 10.1080/00365513.2019.1622031 . HDL : 11250/2739760 . ПМИД 31161807 . S2CID 174806302 .
- ^ Пай Дж., Джоши Р.К., Бхаскар С., Патил С., Бв С., Р.С. и др. (ноябрь 2023 г.). «Возврат референсных диапазонов кортизола у людей: роль демографии» (PDF) . Эндокринный . 82 (2): 414–418. дои : 10.1007/s12020-023-03456-x . ПМИД 37501014 . S2CID 260246830 . Архивировано (PDF) из оригинала 3 декабря 2023 года . Проверено 2 февраля 2024 г.
- ^ Йасмин С., Улла А., Ву Б, Чжан Х, Ченг Л.Дж. (2023). «Ферментные имитаторы для чувствительного и селективного обнаружения стероидных метаболитов» . Прикладные материалы и интерфейсы ACS . 15 (11). дои : 10.1021/acsami.2c21980 . ПМИД 36908226 . S2CID 257494057 . Архивировано из оригинала 8 апреля 2023 года . Проверено 8 апреля 2023 г.
- ^ Jump up to: а б с д Референтные диапазоны биохимии в больнице Доброй Надежды, данные получены 8 ноября 2009 г.
- ^ Ротман-Пикиельни П., Руах В., Чен О., Гур Х.Г., Лимор Р., Стерн Н. (август 2006 г.). «Уровни кортизола в сыворотке у пациентов, поступивших в медицинское отделение: прогностические корреляции и влияние возраста, инфекции и сопутствующих заболеваний». Доктор медицинских наук . 332 (2): 61–67. дои : 10.1097/00000441-200608000-00002 . ПМИД 16909051 . S2CID 5004535 .
- ^ Jump up to: а б с д Получено на основе молярных значений с использованием молярной массы 362 г/моль.
- ^ Ди Далмази Г., Фанелли Ф., Заватта Г., Риччи Битти С., Меццулло М., Репачи А. и др. (ноябрь 2019 г.). «Стероидный профиль инциденталом надпочечников: подтипирование субъектов с высоким сердечно-сосудистым риском» . J Clin Эндокринол Метаб . 104 (11): 5519–5528. дои : 10.1210/jc.2019-00365 . ПМИД 31381072 . S2CID 199437371 .
- ^ Скотт С.М., Ваттерберг К.Л. (январь 1995 г.). «Влияние гестационного возраста, постнатального возраста и заболевания на концентрацию кортизола в плазме у недоношенных детей» . Педиатр Рес . 37 (1): 112–6. дои : 10.1203/00006450-199501000-00021 . ПМИД 7700725 . S2CID 21870513 .
- ^ Jump up to: а б Пересчитано из мкг/24 часа с использованием молярной массы 362,460 г/моль.
- ^ Jump up to: а б Гёргес Р., Кнаппе Г., Герл Х., Венц М., Шталь Ф. (апрель 1999 г.). «Диагностика синдрома Кушинга: повторная оценка полуночного кортизола в плазме по сравнению со свободным кортизолом в моче и тест на подавление низкой дозы дексаметазона в большой группе пациентов». Журнал эндокринологических исследований . 22 (4): 241–9. дои : 10.1007/bf03343551 . ПМИД 10342356 . S2CID 1239611 .
- ^ Jump up to: а б Энциклопедия MedlinePlus : Кортизол – моча
- ^ Jump up to: а б Пересчитано из нмоль/24 часа с использованием молярной массы 362,460 г/моль.
- ^ Идзава С., Сугая Н., Огава Н., Широцуки К., Номура С. (апрель 2021 г.). «Исследование валидации кортизола в ногтях: корреляция с месячными уровнями кортизола, оцененными по образцам волос и слюны» . Стресс . 24 (6): 734–741. дои : 10.1080/10253890.2021.1895113 . ПМИД 33792492 . S2CID 232481968 .
- ^ Турпейнен У, Хямяляйнен Э (декабрь 2013 г.). «Определение кортизола в сыворотке, слюне и моче». Лучшие практики и исследования. Клиническая эндокринология и обмен веществ . 27 (6): 795–801. дои : 10.1016/j.beem.2013.10.008 . ПМИД 24275191 .
- ^ Доломи-Фагур Л., Коркафф Дж.Б. (2008). «[Полезно ли измерение уровня свободного кортизола в плазме в отделении интенсивной терапии?]». Annales de Biologie Clinique (на французском языке). 66 (1): 31–41. doi : 10.1684/abc.2008.0189 (неактивен 31 января 2024 г.). ПМИД 18227002 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Майдана П., Бруно О.Д., Меш В. (2013). «[Критический анализ измерений кортизола: обновление]». Медицина (на испанском языке). 73 (6): 579–84. ПМИД 24356273 .
- ^ Вербетен К.К., Ахмет А.Х. (январь 2018 г.). «Роль кортикостероидсвязывающего глобулина в оценке надпочечниковой недостаточности» . Журнал детской эндокринологии и метаболизма . 31 (2): 107–115. дои : 10.1515/jpem-2017-0270 . ПМИД 29194043 . S2CID 28588420 .
- ^ Хенли Д., Лайтман С., Каррелл Р. (октябрь 2016 г.). «Кортизол и КБГ — доставка кортизола в нужное место в нужное время» (PDF) . Фармакология и терапия . 166 : 128–135. doi : 10.1016/j.pharmthera.2016.06.020 . hdl : 1983/d7ed507d-52d5-496b-ae1f-de220ae1b190 . ПМИД 27411675 . Архивировано (PDF) из оригинала 20 августа 2023 года . Проверено 8 марта 2023 г.
- ^ де Медейрос Г.Ф., Лафенетр П., Джантахин Ю., Серпа Дж.К., Чжан К.Л., Мехта М.М. и др. (2019). «Дефицит кортикостероидсвязывающего глобулина специфически ухудшает консолидацию контекстуальной и узнавающей памяти у мышей-самцов». Нейроэндокринология . 109 (4): 322–332. дои : 10.1159/000499827 . ПМИД 30904918 . S2CID 85498121 .
- ^ Хенли Д.Э., Лайтман С.Л. (апрель 2011 г.). «Новые сведения о кортикостероидсвязывающем глобулине и доставке глюкокортикоидов». Нейронаука . 180 : 1–8. doi : 10.1016/j.neuroscience.2011.02.053 . ПМИД 21371536 . S2CID 26843500 .
- ^ Сальсано К, Сарасино Дж, Кардилло Дж (октябрь 2021 г.). «Возможное поражение надпочечников при синдроме длительного COVID» . Медицина (Каунас) . 57 (10): 1087. doi : 10.3390/medicina57101087 . ПМЦ 8537520 . ПМИД 34684123 .
- ^ Грейнджер Д.А., Хибель Л.К., Фортунато К.К., Капелевски CH (ноябрь 2009 г.). «Влияние лекарств на кортизол в слюне: тактика и стратегия минимизации воздействия в науке о поведении и развитии». Психонейроэндокринология . 34 (10): 1437–48. doi : 10.1016/j.psyneuen.2009.06.017 . ПМИД 19632788 . S2CID 3100315 .
- ^ Лейн Дж (2006). «Могут ли неинвазивные измерения глюкокортикоидов использоваться в качестве надежных индикаторов стресса у животных?». Благополучие животных . 15 (4): 331–342. дои : 10.1017/S0962728600030657 . S2CID 80026053 .
- ^ Лин Г.К., Смайлходжич М., Бандиан А.М., Фридл Х.П., Лейтгеб Т., Оертер С. и др. (август 2020 г.). «Барьерная модель эпителия подчелюстной слюнной железы человека in vitro на основе одного клеточного клона клеточной линии HTB-41: создание и применение для исследований транспорта биомаркеров» . Биомедицины . 8 (9): 302. doi : 10.3390/biomedicines8090302 . ПМЦ 7555419 . ПМИД 32842479 .
- ^ Ли Д.И., Ким Э., Чой М.Х. (2015). «Технические и клинические аспекты кортизола как биохимического маркера хронического стресса» . Отчеты БМБ . 48 (4): 209–216. дои : 10.5483/bmbrep.2015.48.4.275 . ПМЦ 4436856 . ПМИД 25560699 .
- ^ Эль-Фархан Н., Рис Д.А., Эванс С. (май 2017 г.). «Измерение кортизола в сыворотке, моче и слюне – достаточно ли хороши наши анализы?» . Анналы клинической биохимии . 54 (3): 308–322. дои : 10.1177/0004563216687335 . ПМИД 28068807 . S2CID 206397561 .
- ^ «Синдром Кушинга» . Библиотека медицинских концепций Lecturio . Архивировано из оригинала 22 сентября 2021 года . Проверено 11 июля 2021 г.
- ^ «Синдром Кушинга» . Национальная информационная служба эндокринных и метаболических заболеваний (NEMDIS). Июль 2008 г. Архивировано из оригинала 10 февраля 2015 г. Проверено 16 марта 2015 г.
Эти доброкачественные или нераковые опухоли гипофиза секретируют дополнительный АКТГ. У большинства людей с этим расстройством имеется одна аденома. Эта форма синдрома, известная как болезнь Кушинга.
- ^ Форбис П (2005). Медицинские эпонимы Стедмана (2-е изд.). Балтимор, Мэриленд: Липпинкотт Уильямс и Уилкинс. п. 167. ИСБН 978-0-7817-5443-9 . Архивировано из оригинала 24 февраля 2024 года . Проверено 30 августа 2017 г.
- ^ Дэвис Э., Кеньон С.Дж., Фрейзер Р. (июнь 1985 г.). «Роль ионов кальция в механизме стимуляции АКТГ синтеза кортизола». Стероиды . 45 (6): 551–60. дои : 10.1016/0039-128X(85)90019-4 . ПМИД 3012830 . S2CID 24454836 .
- ^ Плотский П.М., Отто С., Сапольский Р.М. (сентябрь 1986 г.). «Ингибирование секреции иммунореактивного кортикотропин-рилизинг фактора в гипофизарно-портальную циркуляцию за счет задержки глюкокортикоидной обратной связи». Эндокринология . 119 (3): 1126–30. дои : 10.1210/эндо-119-3-1126 . ПМИД 3015567 .
- ^ Минтон Дж. Э., Парсонс К. М. (март 1993 г.). «Адренокортикотропный гормон и реакция кортизола на фактор, высвобождающий кортикотропин, и лизин-вазопрессин у свиней». Журнал зоотехники . 71 (3): 724–9. дои : 10.2527/1993.713724x . ПМИД 8385088 .
- ^ Jump up to: а б Фэйрчайлд С.С., Шеннон К., Кван Э., Мишелл Р.И. (февраль 1984 г.). «Фактор, модифицирующий глюкостероидный ответ Т-клеточного происхождения (GRMFT): уникальный лимфокин, вырабатываемый нормальными Т-лимфоцитами и гибридомой Т-клеток» . Журнал иммунологии . 132 (2): 821–7. дои : 10.4049/jimmunol.132.2.821 . ПМИД 6228602 . S2CID 27300153 .
- ^ Дворжак М (1971). «Уровни 17-гидроксикортикостероида в плазме у здоровых телят и телят с диареей». Британский ветеринарный журнал . 127 : 372.
- ^ Стит Р.Д., МакКаллум Р.Э. (1986). «Общее влияние эндотоксина на глюкокортикоидные рецепторы в тканях млекопитающих». Циркуляторный шок . 18 (4): 301–9. ПМИД 3084123 .
- ^ Микоша А.С., Пушкаров И.С., Челнакова И.С., Ременников Г.Я. (1991). «Калиевая регуляция биосинтеза гормонов в надпочечниках морских свинок под действием дигидропиридинов: возможные механизмы изменения стероидогенеза, индуцированного 1,4,дигидропиридинами, в диспергированных адренокортицитах». Физиол. [Киев] . 37:60 .
- ^ «Амир Саадаллах Аль-Зако» (PDF) . Архивировано из оригинала (PDF) 11 ноября 2013 года . Проверено 11 июля 2013 г.
- ^ Мендельсон Ф.А., Маки С. (июль 1975 г.). «Связь внутриклеточного К + и стероидогенеза в изолированных клетках клубочковой зоны надпочечников и пучковых клетках». Клиническая наука и молекулярная медицина . 49 (1): 13–26. дои : 10.1042/cs0490013 . ПМИД 168026 . S2CID 24873537 .
- ^ Уэда Ю., Хонда М., Цучия М., Ватанабэ Х., Изуми Ю., Ширацучи Т. и др. (апрель 1982 г.). «Реакция плазменного АКТГ и гормонов надпочечников на нагрузку калием при гипертонической болезни» . Японский тиражный журнал . 46 (4): 317–22. дои : 10.1253/jcj.46.317 . ПМИД 6283190 .
- ^ Бауман К., Мюллер Дж. (1972). «Влияние калия на окончательный статус биосинтеза альдостерона у крыс. I 18-гидроксилирование и 18-гидроксидегидрирование. II бета-гидроксилирование». Акта Эндокринол . 69 (4): I 701–717, II 718–730. дои : 10.1530/acta.0.0690701 . ПМИД 5067076 .
- ^ ЛаСелле П.Л., Морган Э.С., Этуотер ЕС (1964). «Исследование общего содержания калия в организме у больных ревматоидным артритом». Труды ежегодного собрания Американской ассоциации ревматистов, артрит и ревматизм . 7 (3): 321.
- ^ Броуди С., Преут Р., Шоммер К., Шюрмейер Т.Х. (январь 2002 г.). «Рандомизированное контролируемое исследование высоких доз аскорбиновой кислоты для снижения артериального давления, кортизола и субъективных реакций на психологический стресс». Психофармакология . 159 (3): 319–24. дои : 10.1007/s00213-001-0929-6 . ПМИД 11862365 . S2CID 2778669 .
- ^ Сильверман М.Н., Пирс Б.Д., Бирон К.А., Миллер А.Х. (2005). «Иммунная модуляция оси гипоталамо-гипофиз-надпочечники (HPA) во время вирусной инфекции» . Вирусная иммунология . 18 (1): 41–78. дои : 10.1089/vim.2005.18.41 . ПМЦ 1224723 . ПМИД 15802953 .
- ^ Робсон П.Дж., Бланнин А.К., Уолш Н.П., Кастелл Л.М., Глисон М. (февраль 1999 г.). «Влияние интенсивности, продолжительности и восстановления упражнений на функцию нейтрофилов in vitro у спортсменов-мужчин». Международный журнал спортивной медицины . 20 (2): 128–35. дои : 10.1055/s-2007-971106 . ПМИД 10190775 . S2CID 2572545 .
- ^ Фукуа Дж.С., Рогол А.Д. (июль 2013 г.). «Нейроэндокринные изменения у тренирующегося человека: значение для энергетического гомеостаза». Метаболизм . 62 (7): 911–21. дои : 10.1016/j.metabol.2013.01.016 . ПМИД 23415825 .
- ^ Смит Дж.Л., Гроппер С.А., Грофф Дж.Л. (2009). Расширенное питание и обмен веществ человека . Бельмонт, Калифорния: Обучение Уодсворта Cengage. п. 247. ИСБН 978-0-495-11657-8 .
- ^ Уиттакер Дж., Харрис М. (март 2022 г.). «Низкоуглеводные диеты и мужской кортизол и тестостерон: систематический обзор и метаанализ» . Питание и здоровье . 28 (4): 543–554. дои : 10.1177/02601060221083079 . ПМЦ 9716400 . ПМИД 35254136 . S2CID 247251547 .
- ^ Стахович М., Лебедзиньска А. (декабрь 2016 г.). «Влияние компонентов диеты на уровень кортизола» . Европейские исследования и технологии в области пищевых продуктов . 242 (12): 2001–2009. дои : 10.1007/s00217-016-2772-3 . ISSN 1438-2385 . S2CID 88721472 .
- ^ Хэггстрем М., Ричфилд Д. (2014). «Схема путей стероидогенеза человека» . Викижурнал медицины . 1 (1). дои : 10.15347/wjm/2014.005 . ISSN 2002-4436 .
- ^ Сталдер Т., Киршбаум С. (2013). «Кортизол». Энциклопедия поведенческой медицины . стр. 507–512. дои : 10.1007/978-1-4419-1005-9_171 . ISBN 978-1-4419-1005-9 .
- ^ Jump up to: а б Микула Р., Прушиньска-Ошмалек Е., Пщола М., Жонсиньска Дж., Сассек М., Новак К.В. и др. (20 ноября 2021 г.). «Изменения метаболического и гормонального профиля в переходный период у молочного скота – роль спексина» . Ветеринарное исследование BMC . 17 (1): 359. doi : 10.1186/s12917-021-03069-4 . ПМЦ 8605515 . ПМИД 34798894 .
- ^ Пейн А.Х., Хейлз Д.Б. (декабрь 2004 г.). «Обзор стероидогенных ферментов на пути от холестерина к активным стероидным гормонам». Эндокр преп . 25 (6): 947–70. дои : 10.1210/er.2003-0030 . ПМИД 15583024 .
- ^ «Кортизол | Этимология кортизола по этимонлину» . Архивировано из оригинала 22 декабря 2022 года . Проверено 23 апреля 2024 г.
- ^ Уиллетт Л.Б., Эрб Р.Э. (январь 1972 г.). «Краткосрочные изменения уровня кортикоидов в плазме у молочного скота». Журнал зоотехники . 34 (1): 103–11. дои : 10.2527/jas1972.341103x . ПМИД 5062063 .
- ^ «Гипокалиемия у взрослого крупного рогатого скота – метаболические нарушения» . Архивировано из оригинала 9 декабря 2023 года . Проверено 23 апреля 2024 г.
- ^ «Кортикостероиды у животных – Фармакология» . Архивировано из оригинала 20 апреля 2024 года . Проверено 23 апреля 2024 г.
- ^ Шифрин А.Л., Раффаэлли М., Рэндольф Г.В., Гимм О. (19 января 2022 г.). Комплексное руководство по экзамену по эндокринной хирургии . Спрингер. ISBN 978-3-030-84737-1 .
- ^ Маргиорис А.Н., Цацанис С (2011). «Действие АКТГ на надпочечники» . В Chrousos G (ред.). Физиология и заболевания надпочечников . Эндотекст.орг. Архивировано из оригинала 29 ноября 2011 года . Проверено 5 июня 2012 г.
- ^ Jump up to: а б Финкен М.Дж., Эндрюс Р.К., Эндрю Р., Уокер Б.Р. (сентябрь 1999 г.). «Метаболизм кортизола у здоровых молодых людей: половой диморфизм в активности редуктаз А-кольца, но не 11бета-гидроксистероиддегидрогеназ» . Журнал клинической эндокринологии и метаболизма . 84 (9): 3316–3321. дои : 10.1210/jcem.84.9.6009 . ПМИД 10487705 .
- ^ Дамманн С., Стапельфельд С., Мазер Э. (апрель 2019 г.). «Экспрессия и активность кортизол-активирующего фермента 11β-гидроксистероиддегидрогеназы типа 1 ткане- и видоспецифичны». Химико-биологические взаимодействия . 303 : 57–61. Бибкод : 2019CBI...303...57D . дои : 10.1016/j.cbi.2019.02.018 . ПМИД 30796905 . S2CID 73467693 .
- ^ Атанасов А.Г., Нашев Л.Г., Швейцер Р.А., Фрик С., Одерматт А. (июль 2004 г.). «Гексозо-6-фосфатдегидрогеназа определяет направление реакции 11бета-гидроксистероиддегидрогеназы типа 1 как оксоредуктазы» . Письма ФЭБС . 571 (1–3): 129–133. Бибкод : 2004FEBSL.571..129A . doi : 10.1016/j.febslet.2004.06.065 . ПМИД 15280030 . S2CID 6360244 .
- ^ Томлинсон Дж.В., Уокер Э.А., Буялска И.Дж., Дрейпер Н., Лавери Г.Г., Купер М.С. и др. (октябрь 2004 г.). «11бета-гидроксистероиддегидрогеназа типа 1: тканеспецифичный регулятор глюкокортикоидного ответа» . Эндокринные обзоры . 25 (5): 831–66. дои : 10.1210/er.2003-0031 . ПМИД 15466942 .
- ^ «6бета-гидроксикортизол» . Архивировано из оригинала 20 ноября 2021 года . Проверено 20 ноября 2021 г.
- ^ Луцери Ф, Фаттори С, Луцери С, Зорн М, Маннаиони П, Мессери Дж (декабрь 2001 г.). «Газовая хроматография-масс-спектрометрическое измерение соотношения 6бета-ОН-кортизол/кортизол в моче человека: специфический маркер ферментативной индукции». Клин Химическая Лаборатория Мед . 39 (12): 1234–9. дои : 10.1515/CCLM.2001.198 . ПМИД 11798083 . S2CID 12216877 .
- ^ Хуан Ф.Р., Чжоу С., Чжан XY, Шэнь Ю, Чжан Х.В., Ван Ю.К. и др. (октябрь 2021 г.). «Влияние генотипа CYP2C19 на воздействие вориконазола и влияние вориконазола на активность CYP3A у пациентов со злокачественными новообразованиями крови». Ксенобиотика . 51 (10): 1199–1206. дои : 10.1080/00498254.2021.1969481 . ПМИД 34402388 . S2CID 237150260 .
- ^ Акинос Б.М., Гарсиа Арабегети Х., Кантерос Т.М., де Мигель В., Сцибона П., Файнштейн-Дэй П. (2021). «[Криз надпочечников, связанный с употреблением модафинила]». Medicina (B Aires) (на испанском языке). 81 (5): 846–849. ПМИД 34633961 .
- ^ Jump up to: а б ван Стааверен Н., Тейшейра Д.Л., Хэнлон А., Бойл Л.А. (2015). «Влияние смешивания целых свиней-самцов перед транспортировкой на убой на поведение, благополучие и повреждения туш» . ПЛОС ОДИН . 10 (4): e0122841. Бибкод : 2015PLoSO..1022841V . дои : 10.1371/journal.pone.0122841 . ПМЦ 4382277 . ПМИД 25830336 .
- ^ Шальке Э., Стихнот Дж., Отт С., Джонс-Бааде Р. (2007). «Клинические признаки, вызванные использованием электродрессировочных ошейников на собаках в бытовых ситуациях». Прикладная наука о поведении животных . 105 (4): 369–380. дои : 10.1016/j.applanim.2006.11.002 . S2CID 31552322 .
- ^ Jump up to: а б Аккорси П.А., Карлони Э., Вальсекки П., Виджиани Р., Гамберони М., Таманини С. и др. (январь 2008 г.). «Определение кортизола в волосах и фекалиях домашних кошек и собак». Общая и сравнительная эндокринология . 155 (2): 398–402. дои : 10.1016/j.ygcen.2007.07.002 . ПМИД 17727851 .
- ^ Мёстль Э., Мессманн С., Багу Э., Робиа К., Пальме Р. (декабрь 1999 г.). «Измерение концентрации метаболитов глюкокортикоидов в фекалиях домашнего скота». Центральный журнал ветеринарной медицины. Ряд А. 46 (10): 621–631. дои : 10.1046/j.1439-0442.1999.00256.x . ПМИД 10638300 .
Внешние ссылки
[ редактировать ]- Кортизол МС Спектр
- Кортизол: монография по аналитам – Ассоциация клинической биохимии и лабораторной медицины