Дофамин
 Скелетная формула дофамина | |
 Шаровидная модель молекулы дофамина в растворе. В твердом состоянии дофамин принимает цвиттер-ионную форму. [1] [2] | |
| Клинические данные | |
|---|---|
| Другие имена |
|
| Физиологические данные | |
| Исходные ткани | черная субстанция ; вентральная покрышка ; многие другие |
| Целевые ткани | Общесистемный |
| Рецепторы | Д1 , Д2 2 , Д3 3 , Д4 4 , Д5 5 , ТААР1 [3] |
| Агонисты | Прямые: апоморфин , бромокриптин. Косвенное : кокаин , амфетамин. |
| Антагонисты | Нейролептики , метоклопрамид , домперидон. |
| Предшественник | Фенилаланин , тирозин и L-ДОФА. |
| Биосинтез | ДОФА-декарбоксилаза |
| Метаболизм | МАО , КОМТ [3] |
| Идентификаторы | |
| Номер CAS | |
| ПабХим CID | |
| ИЮФАР/БПС | |
| Лекарственный Банк | |
| ХимическийПаук | |
| НЕКОТОРЫЙ | |
| КЕГГ | |
| Панель управления CompTox ( EPA ) | |
| Информационная карта ECHA | 100.000.101 |
| Химические и физические данные | |
| Формула | С 8 Н 11 Н О 2 |
| Молярная масса | 153.181 g·mol −1 |
| 3D model ( JSmol ) | |
Дофамин ( DA сокращение 3,4- ) дигидроксифенэтиламина нейромодулирующую играет . молекулу , представляет собой , которая несколько важных ролей в клетках Это органическое химическое вещество семейств катехоламинов и фенэтиламинов . Дофамин составляет около 80% содержания катехоламинов в мозге. Это амин, синтезируемый путем удаления карбоксильной группы из молекулы его предшественника химического L-ДОФА , который синтезируется в мозге и почках. Дофамин также синтезируется в растениях и большинстве животных. В мозге дофамин действует как нейротрансмиттер — химическое вещество, выделяемое нейронами (нервными клетками) для отправки сигналов другим нервным клеткам. Нейротрансмиттеры синтезируются в определенных областях мозга, но системно влияют на многие регионы. Мозг включает в себя несколько различных путей дофамина , один из которых играет важную роль в мотивационном компоненте поведения, мотивированного вознаграждением . Ожидание большинства видов вознаграждений повышает уровень дофамина в мозгу. [4] и многие вызывающие привыкание, наркотики, увеличивают высвобождение дофамина или блокируют его обратный захват в нейроны после высвобождения. [5] Другие пути дофамина в мозге участвуют в контроле моторики и контроле высвобождения различных гормонов. Эти пути и группы клеток образуют дофаминовую систему, которая является нейромодулирующей . [5]
В популярной культуре и средствах массовой информации дофамин часто изображается как основной химикат удовольствия, но в настоящее время в фармакологии существует мнение, что вместо этого дофамин придает мотивационную значимость ; [6] [7] [8] другими словами, дофамин сигнализирует о воспринимаемой мотивационной значимости (т. е. о желательности или неприятии) результата, который, в свою очередь, подталкивает поведение организма к достижению этого результата или от него. [8] [9] Именно эндоканнабиноид , 2-арахидоноилглицерин (2-AG: C 23 H 38 O 4 ; 20: 4 , ω-6 ), формирует аккумбальное кодирование сигнал - мотивированного поведения посредством активации рецептора CB1 в вентральной покрышке и тем самым модулирует сигнал. -вызванные кратковременные выбросы дофамина во время погони за наградой . [ нужны разъяснения ] [10]
За пределами центральной нервной системы дофамин действует главным образом как местный паракринный мессенджер. В кровеносных сосудах он ингибирует высвобождение норадреналина и действует как сосудорасширяющее средство ; в почках увеличивает экскрецию натрия и диурез; в поджелудочной железе снижает выработку инсулина; в пищеварительной системе снижает перистальтику желудочно-кишечного тракта и защищает слизистую оболочку кишечника ; а в иммунной системе снижает активность лимфоцитов . За исключением кровеносных сосудов, дофамин в каждой из этих периферических систем синтезируется локально и оказывает свое действие вблизи клеток, которые его выделяют.
Некоторые важные заболевания нервной системы связаны с дисфункциями дофаминовой системы, и некоторые из ключевых лекарств, используемых для их лечения, действуют путем изменения эффектов дофамина. Болезнь Паркинсона , дегенеративное состояние, вызывающее тремор и двигательные нарушения, вызвана потерей секретирующих дофамин нейронов в области среднего мозга, называемой черной субстанцией . Его метаболический предшественник L-DOPA может быть произведен; Леводопа , чистая форма L-ДОФА, является наиболее широко используемым средством лечения болезни Паркинсона. Есть доказательства того, что шизофрения связана с изменением уровня активности дофамина, и большинство антипсихотических препаратов, используемых для лечения этого заболевания, представляют собой антагонисты дофамина , которые снижают активность дофамина. [11] Подобные препараты-антагонисты дофамина также являются одними из наиболее эффективных средств против тошноты . Синдром беспокойных ног и синдром дефицита внимания с гиперактивностью (СДВГ) связаны со снижением активности дофамина. [12] Дофаминергические стимуляторы могут вызывать привыкание в высоких дозах, но некоторые из них используются в более низких дозах для лечения СДВГ. Сам дофамин доступен в виде готового лекарства для внутривенных инъекций . Он полезен при лечении тяжелой сердечной недостаточности или кардиогенного шока . [13] У новорожденных детей его можно применять при гипотонии и септическом шоке . [14]
Структура [ править ]
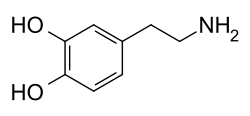
Молекула дофамина состоит из катехиновой структуры ( бензольное кольцо с двумя боковыми гидроксильными группами) с одной аминогруппой , присоединенной через этильную цепь. [15] Таким образом, дофамин является простейшим возможным катехоламином , семейством, которое также включает нейротрансмиттеры норадреналин и адреналин . [16] Наличие бензольного кольца с присоединенным амином делает его замещенным фенэтиламином , семейством, включающим многочисленные психоактивные вещества . [17]
Как и большинство аминов, дофамин представляет собой органическое основание . [18] Как основание , он обычно протонируется в кислой среде (при кислотно-основной реакции ). [18] Протонированная форма хорошо растворима в воде и относительно стабильна, но может окисляться при воздействии кислорода или других окислителей . [18] В основных средах дофамин не протонируется. [18] В этой форме свободного основания оно менее водорастворимо, а также более реакционноспособно. [18] Из-за повышенной стабильности и водорастворимости протонированной формы дофамин поставляется для химического или фармацевтического использования в виде гидрохлорида дофамина , то есть гидрохлоридной соли , которая образуется при соединении дофамина с соляной кислотой . [18] В сухом виде гидрохлорид дофамина представляет собой мелкий порошок от белого до желтого цвета. [19]


Биохимия [ править ]
Синтез [ править ]
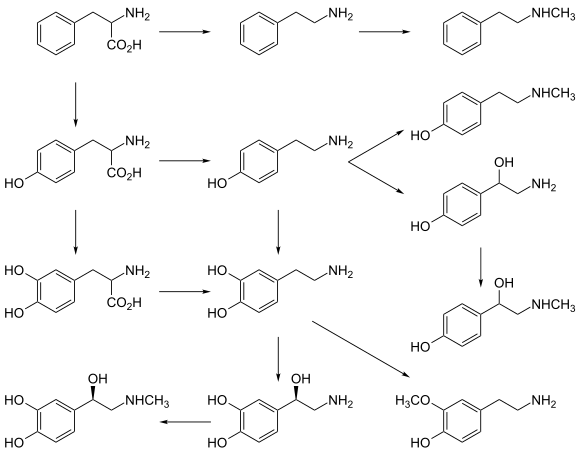
Дофамин синтезируется в ограниченном наборе типов клеток, главным образом в нейронах и клетках мозгового надпочечников слоя . [23] Первичными и второстепенными путями метаболизма являются соответственно:
- Первичный: L -фенилаланин → L -тирозин → L -ДОФА → Дофамин. [20] [21]
- Второстепенные: L -фенилаланин → L -тирозин → п -тирамин → дофамин. [20] [21] [22]
- Второстепенные: L -фенилаланин → м -тирозин → м -тирамин → дофамин. [22] [24] [25]
Прямой предшественник дофамина, L -ДОФА , может быть синтезирован опосредованно из незаменимой аминокислоты фенилаланина или непосредственно из заменимой аминокислоты тирозина . [26] Эти аминокислоты содержатся почти в каждом белке и поэтому легко доступны в пище, наиболее распространенным из которых является тирозин. Хотя дофамин также содержится во многих видах пищи, он не способен преодолевать гематоэнцефалический барьер , который окружает и защищает мозг. [27] Поэтому он должен синтезироваться внутри мозга для выполнения своей нейрональной активности . [27]
L -Фенилаланин превращается в L -тирозин под действием фермента фенилаланингидроксилазы с молекулярным кислородом (O 2 ) и тетрагидробиоптерином в качестве кофакторов . L -тирозин превращается в L -ДОФА ферментом тирозингидроксилазой с тетрагидробиоптерином, O 2 и железом (Fe 2+ ) в качестве кофакторов. [26] L -ДОФА превращается в дофамин под действием фермента декарбоксилазы ароматических L -аминокислот (также известного как декарбоксилаза ДОФА) с пиридоксальфосфатом в качестве кофактора. [26]
Сам дофамин используется в качестве предшественника в синтезе нейротрансмиттеров норадреналина и адреналина. [26] Дофамин превращается в норадреналин под действием фермента дофамин-β-гидроксилазы с участием O 2 и L -аскорбиновой кислоты в качестве кофакторов. [26] Норадреналин превращается в адреналин под действием фермента фенилэтаноламин- N -метилтрансферазы с S -аденозил- L -метионином в качестве кофактора. [26]
Некоторые из кофакторов также требуют собственного синтеза. [26] Дефицит любой необходимой аминокислоты или кофактора может нарушить синтез дофамина, норадреналина и адреналина. [26]
Деградация [ править ]
Дофамин расщепляется на неактивные метаболиты под действием ряда ферментов — моноаминоксидазы (МАО), катехол -О -метилтрансферазы (КОМТ) и альдегиддегидрогеназы (АЛДГ), действующих последовательно. [28] Обе изоформы моноаминоксидазы, МАО-А и МАО-В , эффективно метаболизируют дофамин. [26] Существуют разные пути распада, но основным конечным продуктом является гомованилиновая кислота (ГВА), биологическая активность которой неизвестна. [28] Из кровотока гомованилиновая кислота фильтруется почками и затем выводится с мочой. [28] Двумя основными метаболическими путями, которые превращают дофамин в ГВА, являются: [29]
- Дофамин → ДОПАЛ → ДОФАК → ГВА – катализируется МАО, АЛДГ и КОМТ соответственно.
- Дофамин → 3-метокситирамин → ГВА – катализируется КОМТ и МАО+АЛДГ соответственно.
В клинических исследованиях шизофрении измерения гомованилиновой кислоты в плазме использовались для оценки уровня активности дофамина в мозге. Однако сложность этого подхода заключается в выделении высокого уровня гомованилиновой кислоты в плазме, обусловленного метаболизмом норадреналина. [30] [31]
Хотя дофамин обычно расщепляется ферментом оксидоредуктазой , он также подвержен окислению в результате прямой реакции с кислородом, в результате чего образуются хиноны и различные свободные радикалы . в качестве продуктов [32] Скорость окисления может быть увеличена за счет присутствия трехвалентного железа или других факторов. Хиноны и свободные радикалы, образующиеся в результате автоокисления дофамина, могут отравлять клетки , и есть доказательства того, что этот механизм может способствовать гибели клеток, которая происходит при болезни Паркинсона и других состояниях. [33]
Функции [ править ]
Клеточные эффекты [ править ]
| Семья | Рецептор | Ген | Тип | Механизм |
|---|---|---|---|---|
| Д 1 -подобный | Д 1 | ДРД1 | Г с -связанный. | Повышение внутриклеточного уровня цАМФ путем активации аденилатциклазы . |
| Д 5 | ДРД5 | |||
| Д 2 -подобный | DД2 | ДРД2 | Г я -связанный. | Снижение внутриклеточного уровня цАМФ путем ингибирования аденилатциклазы . |
| Д 3 | ДРД3 | |||
| Д 4 | ДРД4 | |||
| ТААР | ТААР1 | ТААР1 | Г с -связанный. G q -связанный. | Повышение внутриклеточного уровня цАМФ и внутриклеточная концентрация кальция. |
Дофамин оказывает свое действие путем связывания и активации рецепторов клеточной поверхности . [23] У людей дофамин обладает высокой аффинностью связывания с дофаминовыми рецепторами и рецептором 1, связанным с следами аминов человека (hTAAR1). [3] [34] У млекопитающих пять подтипов рецепторов дофамина идентифицировано , обозначенных от D1 до D5 . [23] Все они функционируют как метаботропные , рецепторы связанные с G-белком , а это означает, что они оказывают свое воздействие через сложную систему вторичных мессенджеров . [35] Эти рецепторы можно разделить на два семейства, известные как D1 - подобные и D2 - подобные . [23] Для рецепторов, расположенных на нейронах нервной системы, конечным эффектом D1 - подобной активации (D1 и D5 ) может быть возбуждение (через открытие натриевых каналов ) или торможение (через открытие калиевых каналов ); Конечным эффектом D2 - подобной активации (D2 , D3 и D4 ) обычно является торможение нейрона-мишени. [35] Следовательно, неверно описывать сам дофамин как возбуждающий или тормозящий: его влияние на нейрон-мишень зависит от того, какие типы рецепторов присутствуют на мембране этого нейрона, а также от внутренних реакций этого нейрона на второй мессенджер цАМФ . [35] Рецепторы D 1 — самые многочисленные дофаминовые рецепторы в нервной системе человека; Следующими идут D2 - рецепторы; D3 Рецепторы , D4 и D5 присутствуют на значительно более низких уровнях. [35]
выпуск и повторный захват Хранение ,
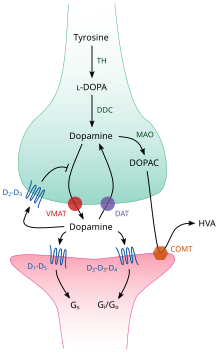
TH: тирозингидроксилаза
ДОФА: L-ДОФА
DAT: транспортер дофамина
DDC: ДОФА-декарбоксилаза
VMAT: везикулярный переносчик моноаминов 2
МАО: моноаминоксидаза.
COMT: катехол-О-метилтрансфераза.
ЧТО: Гомованилиновая кислота
Внутри мозга дофамин действует как нейромедиатор и нейромодулятор и контролируется набором механизмов, общих для всех моноаминовых нейротрансмиттеров . [23] После синтеза дофамин транспортируется из в синаптические пузырьки — растворенным переносчиком везикулярным переносчиком моноаминов VMAT2 цитозоля . [36] Дофамин сохраняется в этих пузырьках до тех пор, пока не будет выброшен в синаптическую щель . В большинстве случаев высвобождение дофамина происходит посредством процесса, называемого экзоцитозом , который вызывается потенциалами действия , но это также может быть вызвано активностью внутриклеточного рецептора, связанного с следами аминов , TAAR1 . [34] TAAR1 представляет собой рецептор с высоким сродством к дофамину, следовым аминам и некоторым замещенным амфетаминам , который расположен вдоль мембран во внутриклеточной среде пресинаптической клетки; [34] Активация рецептора может регулировать передачу сигналов дофамина, вызывая ингибирование обратного захвата и оттока дофамина , а также ингибируя возбуждение нейронов посредством разнообразного набора механизмов. [34] [37]
Попадая в синапс, дофамин связывается с дофаминовыми рецепторами и активирует их. [38] могут быть постсинаптические рецепторы дофамина, расположенные на дендритах (постсинаптический нейрон), или пресинаптические ( например, D2sh Это ауторецепторы и пресинаптические рецепторы D3 ) , расположенные на мембране окончания аксона (пресинаптический нейрон). . [23] [38] После того как постсинаптический нейрон вызывает потенциал действия, молекулы дофамина быстро отрываются от своих рецепторов. Затем они всасываются обратно в пресинаптическую клетку посредством обратного захвата , опосредованного либо переносчиком дофамина , либо переносчиком моноаминов плазматической мембраны . [39] Вернувшись в цитозоль, дофамин может либо расщепляться моноаминоксидазой , либо переупаковываться в везикулы с помощью VMAT2, что делает его доступным для будущего высвобождения. [36]
В мозге уровень внеклеточного дофамина модулируется двумя механизмами: фазической и тонической передачей . [40] Фазическое высвобождение дофамина, как и высвобождение большинства нейромедиаторов в нервной системе, напрямую обусловлено потенциалами действия в дофаминсодержащих клетках. [40] Тоническая передача дофамина происходит, когда небольшие количества дофамина высвобождаются без предшествующих пресинаптических потенциалов действия. [40] Тоническая передача регулируется множеством факторов, включая активность других нейронов и обратный захват нейромедиаторов. [40]
Центральная нервная система [ править ]

Внутри мозга дофамин играет важную роль в исполнительных функциях , контроле моторики , мотивации , возбуждении , подкреплении и вознаграждении , а также в функциях более низкого уровня, включая лактацию , сексуальное удовлетворение и тошноту . Группы дофаминергических клеток и пути составляют дофаминовую систему, которая является нейромодулирующей .
Дофаминергические нейроны (нервные клетки, продуцирующие дофамин) сравнительно немногочисленны — всего около 400 000 в человеческом мозге. [41] — и их клеточные тела сосредоточены группами в нескольких относительно небольших областях мозга. [42] Однако их аксоны проецируются на многие другие области мозга и оказывают мощное воздействие на свои цели. [42] Эти группы дофаминергических клеток были впервые нанесены на карту в 1964 году Анникой Дальстрём и Кьеллом Фуксе, которые присвоили им метки, начинающиеся с буквы «А» (что означает «аминергические»). [43] На их схеме области от А1 до А7 содержат нейромедиатор норадреналин, тогда как от А8 до А14 содержат дофамин. Они определили дофаминергические области: черную субстанцию (группы 8 и 9); вентральная покрышка (группа 10); задний гипоталамус (группа 11); дугообразное ядро (группа 12); ( неясная зона группа 13) и перивентрикулярное ядро (группа 14). [43]
Черная субстанция — небольшая область среднего мозга, которая является компонентом базальных ганглиев . Он состоит из двух частей — входной области, называемой pars reticulata , и выходной области, называемой pars Compacta . Дофаминергические нейроны обнаруживаются преимущественно в компактной части (группа клеток А8) и вблизи нее (группа А9). [42] У людей проекция дофаминергических нейронов из компактной части черной субстанции в дорсальное полосатое тело, называемая нигростриарным путем , играет значительную роль в контроле двигательной функции и в обучении новым двигательным навыкам . [44] Эти нейроны особенно уязвимы к повреждениям, и когда большое их количество погибает, возникает паркинсонический синдром . [45]
Вентральная покрышка (ВТА) — еще одна область среднего мозга. Наиболее известная группа дофаминергических нейронов VTA проецируется в префронтальную кору через мезокортикальный путь , а другая, меньшая группа, проецируется в прилежащее ядро через мезолимбический путь . Вместе эти два пути называются мезокортиколимбической проекцией . [42] [44] ВТА также посылает дофаминергические проекции в миндалевидное тело , поясную извилину , гиппокамп и обонятельную луковицу . [42] [44] Мезокортиколимбические нейроны играют центральную роль в вознаграждении и других аспектах мотивации. [44] Накопление литературы показывает, что дофамин также играет решающую роль в аверсивном обучении, воздействуя на ряд областей мозга. [46] [47] [48]
В заднем гипоталамусе есть дофаминовые нейроны, которые передаются в спинной мозг, но их функция недостаточно изучена. [49] Есть некоторые свидетельства того, что патология в этой области играет роль в синдроме беспокойных ног — состоянии, при котором люди испытывают трудности со сном из-за непреодолимой потребности постоянно двигать частями тела, особенно ногами. [49]
В дугообразном ядре и перивентрикулярном ядре гипоталамуса имеются дофаминовые нейроны, образующие важную проекцию — тубероинфундибулярный путь , идущий к гипофизу , где он влияет на секрецию гормона пролактина . [50] Дофамин является основным нейроэндокринным ингибитором секреции пролактина передней долей гипофиза . [50] Дофамин, вырабатываемый нейронами дугообразного ядра, секретируется в гипофизарную портальную систему срединного возвышения , которая снабжает гипофиз . [50] Пролактиновые клетки , вырабатывающие пролактин, в отсутствие дофамина постоянно секретируют пролактин; дофамин подавляет эту секрецию. [50]
Неопределенная зона, сгруппированная между дугообразными и перивентрикулярными ядрами, проецируется на несколько областей гипоталамуса и участвует в контроле высвобождения гонадотропин-рилизинг-гормона , который необходим для активации развития мужской и женской репродуктивной системы после полового созревания. [50]
Дополнительная группа нейронов, секретирующих дофамин, обнаружена в сетчатке глаза. [51] Эти нейроны являются амакриновыми клетками , то есть у них нет аксонов. [51] Они выделяют дофамин во внеклеточную среду и особенно активны в светлое время суток, замолкая ночью. [51] Этот ретинальный дофамин усиливает активность колбочек сетчатки, одновременно подавляя палочки . В результате повышается чувствительность к цвету и контрастности в условиях яркого освещения за счет снижения чувствительности при тусклом свете. [51]
Базальные ганглии [ править ]

Крупнейшими и наиболее важными источниками дофамина в мозге позвоночных являются черная субстанция и вентральная покрышка. [42] Обе структуры являются компонентами среднего мозга, тесно связанными друг с другом и во многом функционально схожими. [42] Самый крупный компонент базальных ганглиев — полосатое тело. [52] Черная субстанция посылает дофаминергические проекции в дорсальное полосатое тело , тогда как вентральная покрышка посылает дофаминергические проекции аналогичного типа в вентральное полосатое тело . [42]
Прогресс в понимании функций базальных ганглиев был медленным. [52] Наиболее популярные гипотезы в широком смысле предполагают, что базальные ганглии играют центральную роль в выборе действия . [53] Теория выбора действий в своей простейшей форме предполагает, что, когда человек или животное находится в ситуации, когда возможны несколько вариантов поведения, активность базальных ганглиев определяет, какое из них будет выполнено, освобождая эту реакцию от торможения, продолжая при этом тормозить другие двигательные системы. что, если его активировать, это приведет к конкурирующему поведению. [54] Таким образом, согласно этой концепции, базальные ганглии отвечают за инициирование поведения, но не за определение деталей того, как оно осуществляется. Другими словами, они по сути образуют систему принятия решений. [54]
Базальные ганглии можно разделить на несколько секторов, каждый из которых участвует в управлении определенными типами действий. [55] Вентральный сектор базальных ганглиев (содержащий вентральное полосатое тело и вентральную покрышку) действует на самом высоком уровне иерархии, выбирая действия на уровне всего организма. [54] Дорсальные сектора (содержащие дорсальное полосатое тело и черную субстанцию) действуют на более низких уровнях, выбирая конкретные мышцы и движения, которые используются для реализации заданной модели поведения. [55]
Дофамин участвует в процессе выбора действия по крайней мере двумя важными способами. Во-первых, он устанавливает «порог» начала действий. [53] Чем выше уровень активности дофамина, тем меньший стимул требуется для вызова определенного поведения. [53] Как следствие, высокий уровень дофамина приводит к высокому уровню двигательной активности и импульсивному поведению ; низкий уровень дофамина приводит к оцепенению и замедлению реакций. [53] Болезнь Паркинсона, при которой уровни дофамина в цепи черной субстанции значительно снижены, характеризуется скованностью и трудностями при начале движения, однако, когда люди с этим заболеванием сталкиваются с сильными раздражителями, такими как серьезная угроза, их реакции могут быть столь же энергичными, как и таковые у здорового человека. [56] И наоборот, препараты, увеличивающие высвобождение дофамина, такие как кокаин или амфетамин, могут вызывать повышенный уровень активности, включая, в крайнем случае, психомоторное возбуждение и стереотипные движения . [57]
Второй важный эффект дофамина — это «обучающий» сигнал. [53] Когда за действием следует увеличение активности дофамина, схема базальных ганглиев изменяется таким образом, что ту же реакцию легче вызвать, когда аналогичные ситуации возникнут в будущем. [53] Это форма оперантного обусловливания , при которой дофамин играет роль сигнала вознаграждения. [54]
Награда [ править ]

На языке, используемом для обсуждения системы вознаграждения, вознаграждение — это привлекательное и мотивационное свойство стимула, которое вызывает аппетитное поведение (также известное как поведение приближения) и законченное поведение . [58] Поощрительный стимул – это стимул, который может побудить организм приблизиться к нему и принять решение его употребить. [58] Удовольствие , обучение (например, классическое и оперантное обусловливание ) и подход к поведению — три основные функции вознаграждения. [58] Как аспект вознаграждения удовольствие дает определение вознаграждения; [58] однако, хотя все приятные стимулы приносят удовольствие, не все приносящие удовольствие стимулы являются приятными (например, внешние вознаграждения, такие как деньги). [58] [59] Мотивационный или желательный аспект стимулов вознаграждения отражается в поведении, которое они вызывают, тогда как удовольствие от внутренних вознаграждений возникает в результате их потребления после их приобретения. [58] Нейропсихологической моделью, которая различает эти два компонента стимула, приносящего по своей сути вознаграждение, является модель значимости стимулов , в которой «желание» или желание (реже «поиск») [60] ) соответствует аппетитному или приближающемуся поведению, тогда как «нравится» или удовольствие соответствует завершенному поведению. [58] [6] [61] У людей -наркоманов «желание» диссоциируется с «пристрастием», поскольку желание употреблять вызывающий привыкание наркотик увеличивается, а удовольствие, получаемое от его употребления, уменьшается из-за толерантности к наркотику . [6]
В мозге дофамин частично действует как глобальный сигнал вознаграждения. Первоначальная дофаминовая реакция на поощряющий стимул кодирует информацию о значимости , ценности и контексте вознаграждения. [58] В контексте обучения, связанного с вознаграждением, дофамин также действует как сигнал ошибки прогнозирования вознаграждения , то есть степени, в которой ценность вознаграждения является неожиданной. [58] Согласно этой гипотезе, предложенной Монтегю, Даяном и Сейновским, [62] Ожидаемые награды не вызывают второй фазовой реакции дофамина в определенных дофаминергических клетках, но награды, которые являются неожиданными или превышают ожидаемые, вызывают кратковременное увеличение синаптического дофамина, тогда как отсутствие ожидаемого вознаграждения фактически приводит к высвобождению дофамина. опуститься ниже фонового уровня. [58] Гипотеза «ошибки прогнозирования» вызвала особый интерес у вычислительных нейробиологов, поскольку влиятельный метод вычислительного обучения, известный как обучение с временными разностями, интенсивно использует сигнал, который кодирует ошибку прогнозирования. [58] Такое слияние теории и данных привело к плодотворному взаимодействию между нейробиологами и учеными-компьютерщиками, интересующимися машинным обучением . [58]
Данные микроэлектродных записей головного мозга животных показывают, что дофаминовые нейроны в вентральной покрышке (VTA) и черной субстанции сильно активируются самыми разными событиями вознаграждения. [58] Эти дофаминовые нейроны, реагирующие на вознаграждение, в VTA и черной субстанции имеют решающее значение для познания, связанного с вознаграждением, и служат центральным компонентом системы вознаграждения. [6] [63] [64] Функция дофамина варьируется в каждой проекции аксона от VTA и черной субстанции; [6] например, проекция VTA- прилежащее ядро присваивает значимость стимулов («хочу») стимулам вознаграждения и связанным с ними сигналам , проекция VTA- префронтальная кора обновляет ценность различных целей в соответствии с их значимостью стимулов, VTA-амигдала и Проекции VTA-гиппокамп опосредуют консолидацию воспоминаний, связанных с вознаграждением, а пути VTA-прилежащее ядро и путь черной субстанции-дорсальное полосатое тело участвуют в обучении двигательным реакциям, которые облегчают приобретение стимулов вознаграждения. [6] [65] Некоторая активность в дофаминергических проекциях VTA, по-видимому, также связана с предсказанием вознаграждения. [6] [65]
Удовольствие [ править ]
Хотя дофамин играет центральную роль в возникновении «желания», связанного с поведенческими реакциями аппетита или приближения к поощряющим стимулам, детальные исследования показали, что дофамин нельзя просто приравнивать к гедонистическому «пристрастию» или удовольствию, что отражается в законченной поведенческой реакции. [59] Дофаминовая нейротрансмиссия участвует в некоторых, но не во всех аспектах познания, связанного с удовольствием, поскольку центры удовольствия были идентифицированы как внутри дофаминовой системы (т. е. в прилежащем ядре), так и за ее пределами (т. е. вентральное паллидум и парабрахиальное ядро ). [59] [61] [66] Например, прямая электрическая стимуляция дофаминовых путей с помощью имплантированных в мозг электродов воспринимается как удовольствие, и многие виды животных готовы работать, чтобы получить ее. [67] Антипсихотические препараты снижают уровень дофамина и имеют тенденцию вызывать ангедонию — снижение способности испытывать удовольствие. [68] Многие виды приятных переживаний, таких как половой акт, еда и видеоигры, увеличивают выброс дофамина. [69] Все наркотики, вызывающие привыкание, прямо или косвенно влияют на нейротрансмиссию дофамина в прилежащем ядре; [6] [67] эти наркотики усиливают «тягу к наркотикам», что приводит к компульсивному употреблению наркотиков при неоднократном приеме в высоких дозах, предположительно за счет повышения чувствительности стимулов . [61] К препаратам, повышающим концентрацию синаптического дофамина, относятся психостимуляторы , такие как метамфетамин и кокаин. Они вызывают усиление «желательного» поведения, но не сильно меняют выражение удовольствия или уровень насыщения. [61] [67] Однако опиатные наркотики, такие как героин и морфин, приводят к увеличению проявлений поведения «нравится» и «желать». [61] Более того, животные, у которых вентральная покрышка дофаминовой системы отключена, не ищут еды и умрут от голода, если оставить их себе, но если еду положить им в рот, они съедят ее и продемонстрируют выражение лица, указывающее на удовольствие. [70]
Клиническое исследование, проведенное в январе 2019 года, в котором оценивалось влияние предшественника дофамина ( леводопы ), антагониста дофамина ( рисперидона ) и плацебо на реакцию вознаграждения на музыку, включая степень удовольствия, испытываемого во время музыкального озноба , измеряемого по изменениям электродермальной активности. а также субъективные оценки — обнаружено, что манипулирование нейротрансмиссией дофамина двунаправленно регулирует познание удовольствия (в частности, гедоническое воздействие музыки ) у людей. [71] [72] Это исследование показало, что повышенная нейротрансмиссия дофамина действует как обязательное условие приятных гедонистических реакций на музыку у людей. [71] [72]
Исследование, опубликованное в журнале Nature в 1998 году, обнаружило доказательства того, что видеоигры высвобождают дофамин в полосатом теле человека. Этот дофамин связан с обучением, подкреплением поведения, вниманием и сенсомоторной интеграцией. [73] Исследователи использовали позитронно-эмиссионную томографию и 11 С-меченный раклоприд для отслеживания уровня дофамина в мозге во время целенаправленных двигательных задач и обнаружил, что высвобождение дофамина положительно коррелирует с выполнением задач и является наибольшим в вентральном полосатом теле . Это было первое исследование, продемонстрировавшее поведенческие условия, при которых у людей высвобождается дофамин. Это подчеркивает способность позитронно-эмиссионной томографии обнаруживать потоки нейромедиаторов во время изменений в поведении. Согласно исследованиям, потенциально проблематичное использование видеоигр связано с такими личностными качествами, как низкая самооценка и низкая самоэффективность, тревожность, агрессия, а также клинические симптомы депрессии и тревожных расстройств. [74] Кроме того, причины, по которым люди играют в видеоигры, различаются и могут включать в себя преодоление трудностей , социализацию и личное удовлетворение. DSM -5 определяет расстройство, связанное с азартными играми, как психическое расстройство, тесно связанное с расстройством, связанным с азартными играми. Это было поддержано некоторыми исследователями, но также вызвало споры.
Вне центральной нервной системы [ править ]
Дофамин не проникает через гематоэнцефалический барьер, поэтому его синтез и функции в периферических областях в значительной степени независимы от его синтеза и функций в головном мозге. [27] В кровотоке циркулирует значительное количество дофамина, однако его функции там не совсем ясны. [28] Дофамин обнаруживается в плазме крови на уровнях, сравнимых с уровнями адреналина, но у людей более 95% дофамина в плазме находится в форме сульфата дофамина , конъюгата, продуцируемого ферментом сульфотрансферазой 1А3/1А4, действующим на свободный дофамин. [28] Основная часть этого сульфата дофамина вырабатывается в брыжеечных органах. [28] Считается, что выработка сульфата дофамина является механизмом детоксикации дофамина, который поступает с пищей или вырабатывается в процессе пищеварения — его уровень в плазме обычно повышается более чем в пятьдесят раз после еды. [28] Сульфат дофамина не имеет известных биологических функций и выводится с мочой. [28]
Относительно небольшое количество неконъюгированного дофамина в кровотоке может вырабатываться симпатической нервной системой , пищеварительной системой или, возможно, другими органами. [28] Он может действовать на дофаминовые рецепторы в периферических тканях, метаболизироваться или превращаться в норадреналин под действием фермента дофамин-бета-гидроксилазы , который высвобождается в кровоток мозговым веществом надпочечников. [28] Некоторые дофаминовые рецепторы расположены в стенках артерий, где они действуют как сосудорасширяющее средство и ингибитор высвобождения норадреналина из окончаний постганглионарных симпатических нервов (дофамин может ингибировать высвобождение норадреналина, воздействуя на пресинаптические дофаминовые рецепторы, а также на пресинаптические α-1-рецепторы, как и сам норадреналин). [75] Эти реакции могут быть активированы дофамином, высвобождаемым из каротидного тела в условиях низкого содержания кислорода, но неизвестно, выполняют ли артериальные рецепторы дофамина другие биологически полезные функции. [75]
Помимо своей роли в модуляции кровотока, существует несколько периферических систем, в которых дофамин циркулирует в пределах ограниченной области и выполняет экзокринную или паракринную функцию. [28] Периферические системы, в которых дофамин играет важную роль, включают иммунную систему , почки и поджелудочную железу .
Иммунная система [ править ]
В иммунной системе дофамин действует на рецепторы, присутствующие на иммунных клетках, особенно на лимфоцитах . [76] Дофамин также может влиять на иммунные клетки селезенки , костного мозга и системы кровообращения . [77] Кроме того, дофамин может синтезироваться и выделяться самими иммунными клетками. [76] Основное воздействие дофамина на лимфоциты заключается в снижении уровня их активации. Функциональное значение этой системы неясно, но она обеспечивает возможный путь взаимодействия между нервной системой и иммунной системой и может иметь отношение к некоторым аутоиммунным заболеваниям. [77]
Почки [ править ]
Почечная дофаминергическая система расположена в клетках нефрона почки , где присутствуют все подтипы дофаминовых рецепторов. [78] синтезируется дофамин Там же клетками канальцев и выбрасывается в канальцевую жидкость . Его действия включают усиление кровоснабжения почек, повышение скорости клубочковой фильтрации и усиление выведения натрия с мочой. Следовательно, нарушения функции дофамина в почках могут привести к снижению экскреции натрия и, как следствие, к развитию высокого кровяного давления . Имеются убедительные доказательства того, что нарушения в выработке дофамина или рецепторах могут привести к ряду патологий, включая окислительный стресс , отеки и генетическую или эссенциальную гипертензию. Окислительный стресс сам по себе может вызвать гипертонию. [79] Дефекты в системе также могут быть вызваны генетическими факторами или высоким кровяным давлением. [80]
Поджелудочная железа [ править ]
В поджелудочной железе роль дофамина несколько сложна. Поджелудочная железа состоит из двух частей: экзокринного и эндокринного компонентов. Экзокринная часть синтезирует и секретирует пищеварительные ферменты и другие вещества, в том числе дофамин, в тонкую кишку. [81] Функция этого секретируемого дофамина после его попадания в тонкую кишку четко не установлена — возможные варианты включают защиту слизистой оболочки кишечника от повреждения и снижение моторики желудочно-кишечного тракта (скорости, с которой содержимое перемещается через пищеварительную систему). [81]
Островки поджелудочной железы составляют эндокринную часть поджелудочной железы и синтезируют и секретируют гормоны, включая инсулин . в кровоток [81] Имеются доказательства того, что бета-клетки островков, синтезирующих инсулин, содержат рецепторы дофамина и что дофамин снижает количество выделяемого ими инсулина. [81] Источник поступления дофамина четко не установлен: он может поступать из дофамина, который циркулирует в кровотоке и поступает из симпатической нервной системы, или он может синтезироваться локально другими типами клеток поджелудочной железы. [81]
Медицинское использование

Дофамин как промышленное лекарство продается, среди прочего, под торговыми названиями Интропин, Допастат и Ревимин. Он включен в Список основных лекарственных средств Всемирной организации здравоохранения . [82] Он чаще всего используется в качестве стимулирующего препарата при лечении тяжелого низкого кровяного давления , замедления сердечного ритма и остановки сердца . Это особенно важно при лечении этих заболеваний у новорожденных . [83] [14] Его вводят внутривенно. Поскольку период полувыведения дофамина в плазме очень короткий — примерно одна минута у взрослых, две минуты у новорожденных и до пяти минут у недоношенных детей — его обычно вводят непрерывно внутривенно капельно, а не однократно. [84]
Его эффекты, в зависимости от дозировки, включают увеличение выведения натрия почками, увеличение диуреза, увеличение частоты сердечных сокращений и повышение артериального давления . [84] В низких дозах он действует через симпатическую нервную систему, увеличивая силу сокращения сердечной мышцы и частоту сердечных сокращений, тем самым увеличивая сердечный выброс и артериальное давление. [85] Более высокие дозы также вызывают сужение сосудов , что еще больше повышает кровяное давление. [85] [86] В более старой литературе также описываются очень низкие дозы, которые, как считается, улучшают функцию почек без других последствий, но недавние обзоры пришли к выводу, что дозы на таких низких уровнях не эффективны, а иногда могут быть вредными. [87] некоторые эффекты возникают в результате стимуляции дофаминовых рецепторов, выраженные сердечно-сосудистые эффекты возникают в результате действия дофамина α1 Хотя , β1 на и β2 адренергические рецепторы . [88] [89]
Побочные эффекты дофамина включают негативное воздействие на функцию почек и нерегулярное сердцебиение . [85] 50 Установлено, что LD , или смертельная доза, которая, как ожидается, окажется фатальной для 50% населения, составляет: 59 мг/кг (мышь; вводится внутривенно ); 95 мг/кг (мыши; вводят внутрибрюшинно ); 163 мг/кг (крыса; вводят внутрибрюшинно); 79 мг/кг (собака; вводится внутривенно). [90]
Болезни, фармакология и расстройства
Дофаминовая система играет центральную роль в ряде серьезных заболеваний, включая болезнь Паркинсона , синдром дефицита внимания и гиперактивности , синдром Туретта , шизофрению , биполярное расстройство и зависимость . Помимо самого дофамина, существует множество других важных препаратов, которые действуют на дофаминовые системы в различных частях мозга или тела. Некоторые из них используются в медицинских или рекреационных целях, но нейрохимики также разработали множество исследовательских препаратов, некоторые из которых связываются с высоким сродством к определенным типам дофаминовых рецепторов и либо агонизируют , либо противодействуют их эффектам, а многие из них влияют на другие аспекты физиологии дофамина. , [91] включая ингибиторы переносчика дофамина , ингибиторы VMAT и ингибиторы ферментов .
Старение мозга [ править ]
В ряде исследований сообщалось о связанном с возрастом снижении синтеза дофамина и плотности рецепторов дофамина (т. е. количества рецепторов) в головном мозге. [92] Было показано, что это снижение происходит в полосатом теле и экстрастриарных областях. [93] Снижение D1 рецепторов , D2 хорошо и D3 документировано . [94] [95] [96] Считается, что снижение уровня дофамина с возрастом является причиной многих неврологических симптомов, частота которых увеличивается с возрастом, таких как уменьшение размахивания руками и повышенная ригидность . [97] Изменения уровня дофамина также могут вызывать возрастные изменения когнитивной гибкости. [97]
Рассеянный склероз [ править ]
Исследования показали, что дисбаланс дофамина влияет на утомляемость при рассеянном склерозе . [98] У пациентов с рассеянным склерозом дофамин ингибирует выработку IL-17 и IFN-γ мононуклеарными клетками периферической крови. [99]
Болезнь Паркинсона [ править ]
Болезнь Паркинсона — это возрастное заболевание, характеризующееся двигательными расстройствами , такими как скованность тела, замедление движений и дрожание конечностей, когда они не используются. [56] На поздних стадиях заболевание прогрессирует до слабоумия и, в конечном итоге, до смерти. [56] Основные симптомы вызваны потерей клеток черной субстанции, секретирующих дофамин. [100] Эти дофаминовые клетки особенно уязвимы к повреждениям, а различные травмы, включая энцефалит (как показано в книге и фильме «Пробуждение , связанные со спортом »), повторяющиеся сотрясения мозга , и некоторые формы химического отравления, такие как МФТП , могут привести к значительной потере клеток. , вызывая паркинсонический синдром , который по своим основным признакам сходен с болезнью Паркинсона. [101] Однако большинство случаев болезни Паркинсона являются идиопатическими , а это означает, что причину гибели клеток невозможно определить. [101]
Наиболее широко используемым лечением паркинсонизма является введение L-ДОФА, метаболического предшественника дофамина. [27] L-ДОФА преобразуется в дофамин в мозге и различных частях тела с помощью фермента ДОФА-декарбоксилазы. [26] Используется L-ДОФА, а не сам дофамин, поскольку, в отличие от дофамина, он способен преодолевать гематоэнцефалический барьер . [27] Его часто назначают совместно с ферментным ингибитором периферического декарбоксилирования , таким как карбидопа или бенсеразид , чтобы уменьшить количество, превращающееся в дофамин на периферии, и тем самым увеличить количество L-ДОФА, попадающего в мозг. [27] различные неприятные побочные эффекты, такие как дискинезия При регулярном применении L-ДОФА в течение длительного периода времени часто начинают проявляться ; даже в этом случае он считается лучшим доступным вариантом долгосрочного лечения для большинства случаев болезни Паркинсона. [27]
Лечение L-ДОФА не может восстановить утраченные клетки дофамина, но оно заставляет оставшиеся клетки вырабатывать больше дофамина, тем самым компенсируя потерю, по крайней мере, в некоторой степени. [27] На поздних стадиях лечение начинает неэффективным, поскольку потеря клеток настолько серьезна, что оставшиеся не могут производить достаточное количество дофамина независимо от уровня L-ДОФА. [27] Другие препараты, усиливающие функцию дофамина, такие как бромокриптин и перголид , также иногда используются для лечения паркинсонизма, но в большинстве случаев L-ДОФА, по-видимому, обеспечивает лучший компромисс между положительными эффектами и отрицательными побочными эффектами. [27]
Дофаминергические препараты, используемые для лечения болезни Паркинсона, иногда связаны с развитием синдрома дисрегуляции дофамина , который включает в себя чрезмерное использование дофаминергических препаратов и вызванное лекарствами компульсивное участие в естественных вознаграждениях, таких как азартные игры и сексуальная активность. [102] [103] Последнее поведение похоже на поведение, наблюдаемое у людей с поведенческой зависимостью . [102]
Наркомания и психостимуляторы [ править ]

Кокаин , замещенные амфетамины (включая метамфетамин ), аддералл , метилфенидат (продаваемый как риталин или концерта ) и другие психостимуляторы оказывают свое действие главным образом или частично за счет повышения уровня дофамина в мозге с помощью различных механизмов. [104] Кокаин и метилфенидат являются блокаторами переносчика дофамина или ингибиторами обратного захвата ; [105] они неконкурентно ингибируют обратный захват дофамина, что приводит к увеличению концентрации дофамина в синаптической щели. [106] [107] : 54–58 Подобно кокаину, замещенные амфетамины и амфетамин также повышают концентрацию дофамина в синаптической щели , но по разным механизмам. [37] [107] : 147–150
Эффекты психостимуляторов включают увеличение частоты сердечных сокращений, температуры тела и потоотделения; улучшение бдительности, внимания и выносливости; увеличение удовольствия от награждения событиями; но при более высоких дозах возникает возбуждение, тревога или даже потеря контакта с реальностью . [104] Наркотики этой группы могут иметь высокий потенциал привыкания из-за их активирующего воздействия на опосредованную дофамином систему вознаграждения в мозге. [104] Однако некоторые из них также могут быть полезны в более низких дозах для лечения синдрома дефицита внимания и гиперактивности (СДВГ) и нарколепсии . [108] [109] Важным дифференцирующим фактором является начало и продолжительность действия. [104] Кокаин может начать действовать за секунды, если его вводить или вдыхать в форме свободного основания; Эффект длится от 5 до 90 минут. [110] Это быстрое и кратковременное действие делает его эффект легко ощутимым и, следовательно, дает ему высокий потенциал привыкания. [104] Напротив, метилфенидат, принимаемый в форме таблеток, может занять два часа, чтобы достичь пикового уровня в кровотоке. [108] и в зависимости от состава эффект может длиться до 12 часов. [111] Преимущество этих составов длительного действия заключается в уменьшении возможности злоупотребления и улучшении приверженности к лечению за счет использования более удобных режимов дозирования. [112]

Различные наркотики, вызывающие привыкание, вызывают увеличение активности дофамина, связанного с вознаграждением. [104] Стимуляторы, такие как никотин , кокаин и метамфетамин, способствуют повышению уровня дофамина, который, по-видимому, является основным фактором, вызывающим зависимость. Что касается других наркотиков, вызывающих привыкание, таких как опиоидный героин, повышенный уровень дофамина в системе вознаграждения может играть лишь незначительную роль в возникновении зависимости. [113] Когда люди, зависимые от стимуляторов, проходят абстиненцию, они не испытывают физических страданий, связанных с от алкоголя или абстиненцией опиатов; вместо этого они испытывают тягу к наркотику, сильное желание употреблять наркотик, характеризующееся раздражительностью, беспокойством и другими симптомами возбуждения. [114] вызвано психологической зависимостью .
Дофаминовая система играет решающую роль в некоторых аспектах зависимости. На самой ранней стадии генетические различия, которые изменяют экспрессию дофаминовых рецепторов в мозгу, могут предсказать, сочтет ли человек стимуляторы привлекательными или отталкивающими. [115] Потребление стимуляторов приводит к повышению уровня дофамина в мозге, которое длится от минут до часов. [104] Наконец, хроническое повышение уровня дофамина, сопровождающее повторяющееся употребление высоких доз стимуляторов, запускает широкий спектр структурных изменений в мозге, которые ответственны за поведенческие отклонения, характеризующие зависимость. [116] Лечение зависимости от стимуляторов очень сложно, потому что даже если потребление прекращается, тяга, сопровождающая психологическую абстиненцию, не исчезает. [114] Даже когда тяга к наркотику кажется угасшей, она может возникнуть вновь при столкновении со раздражителями, связанными с наркотиком, такими как друзья, места и ситуации. [114] Сети ассоциаций в мозге тесно взаимосвязаны. [117]
и антипсихотические препараты Психоз
Психиатры в начале 1950-х годов обнаружили, что класс препаратов, известных как типичные антипсихотики (также известные как основные транквилизаторы ), часто эффективен для уменьшения психотических симптомов шизофрении. [118] Появление в 1950-х годах первого широко используемого антипсихотика, хлорпромазина (торазина), привело в последующие годы к освобождению многих пациентов с шизофренией из учреждений. [118] К 1970-м годам исследователи поняли, что эти типичные антипсихотики действуют как антагонисты D2 рецепторов . [118] [119] Это осознание привело к так называемой дофаминовой гипотезе шизофрении , которая постулирует, что шизофрения в значительной степени вызвана гиперактивностью дофаминовых систем мозга. [120] Гипотеза дофамина получила дополнительную поддержку в наблюдении, что психотические симптомы часто усиливаются стимуляторами, усиливающими дофамин, такими как метамфетамин, и что эти наркотики также могут вызывать психоз у здоровых людей, если их принимать в достаточно больших дозах. [120] В последующие десятилетия были разработаны другие атипичные антипсихотики , которые имели меньше серьезных побочных эффектов. [118] Многие из этих новых препаратов не действуют непосредственно на дофаминовые рецепторы, а вместо этого косвенно вызывают изменения в активности дофамина. [121] Эти препараты также использовались для лечения других психозов. [118] Антипсихотические препараты оказывают широкое подавляющее действие на большинство типов активного поведения и, в частности, уменьшают бредовое и возбужденное поведение, характерное для явного психоза. [119]
Однако более поздние наблюдения привели к тому, что гипотеза дофамина потеряла популярность, по крайней мере, в ее простой первоначальной форме. [120] Во-первых, у пациентов с шизофренией обычно не наблюдается заметного повышения уровня активности дофамина в мозге. [120] Несмотря на это, многие психиатры и нейробиологи продолжают считать, что шизофрения связана с дисфункцией дофаминовой системы. [118] Однако по мере развития «дофаминовой гипотезы» виды дисфункций, которые она постулирует, имеют тенденцию становиться все более тонкими и сложными. [118]
Психофармаколог Стивен М. Шталь предположил в обзоре 2018 года, что во многих случаях психоза, включая шизофрению, три взаимосвязанные сети, основанные на дофамине, серотонине и глутамате — каждая по отдельности или в различных комбинациях — способствуют перевозбуждению дофамина D 2. Рецепторы вентрального полосатого тела . [122]
Синдром дефицита внимания с гиперактивностью [ править ]
Измененная нейротрансмиссия дофамина вовлечена в синдром дефицита внимания с гиперактивностью (СДВГ), состояние, связанное с нарушением когнитивного контроля , что, в свою очередь, приводит к проблемам с регуляцией внимания ( контроль внимания ), торможению поведения ( тормозящий контроль ), а также к забывчивости вещей или упущению деталей ( работа). память ), среди других проблем. [123] Существуют генетические связи между рецепторами дофамина, переносчиком дофамина и СДВГ, а также связи с другими рецепторами и переносчиками нейромедиаторов. [124] Самая важная связь между дофамином и СДВГ связана с лекарствами, которые используются для лечения СДВГ. [125] Одними из наиболее эффективных терапевтических средств при СДВГ являются психостимуляторы, такие как метилфенидат (риталин, концерта) и амфетамин (эвекео, аддералл, декседрин), препараты, которые повышают уровни дофамина и норадреналина в мозге. [125] Клинические эффекты этих психостимуляторов при лечении СДВГ опосредованы через непрямую активацию рецепторов дофамина и норадреналина, в частности дофаминового рецептора D1 и адренорецептора α2 , в префронтальной коре. [123] [126] [127]
Боль [ править ]
Дофамин играет роль в обработке боли на нескольких уровнях центральной нервной системы, включая спинной мозг, околоводопроводную серую часть , таламус , базальные ганглии и поясную кору . [128] Снижение уровня дофамина связано с болезненными симптомами, которые часто возникают при болезни Паркинсона. [128] Нарушения дофаминергической нейротрансмиссии также возникают при некоторых болезненных клинических состояниях, включая синдром жжения во рту , фибромиалгию и синдром беспокойных ног. [128]
Тошнота [ править ]
Тошнота и рвота в значительной степени определяются активностью постремной области в продолговатом , мозге ствола мозга в области, известной как триггерная зона хеморецепторов . [129] Эта область содержит большую популяцию дофаминовых рецепторов типа D2 . [129] Следовательно, препараты, активирующие рецепторы D2 , могут вызывать тошноту. [129] В эту группу входят некоторые лекарства, назначаемые при болезни Паркинсона, а также другие агонисты дофамина, такие как апоморфин . [130] В некоторых случаях антагонисты D 2 -рецепторов, такие как метоклопрамид, полезны в качестве средств против тошноты . [129]
биология Сравнительная и эволюция
Микроорганизмы [ править ]
Нет никаких сообщений о дофамине у архей , но он был обнаружен у некоторых типов бактерий и у простейших, называемых Tetrahymena . [131] Возможно, еще важнее то, что существуют виды бактерий, которые содержат гомологи всех ферментов, которые животные используют для синтеза дофамина. [132] Было высказано предположение, что животные получили свой механизм синтеза дофамина от бактерий посредством горизонтального переноса генов , который мог произойти относительно поздно в эволюционном времени, возможно, в результате симбиотического включения бактерий в эукариотические клетки, которые дали начало митохондриям . [132]
Животные [ править ]
Дофамин используется в качестве нейромедиатора у большинства многоклеточных животных. [133] У губок имеется только одно сообщение о присутствии дофамина без указания его функции; [134] однако дофамин был обнаружен в нервной системе многих других -симметричных видов, включая книдарий радиально , гидру и некоторые кораллы . [135] Это датирует появление дофамина как нейромедиатора самым ранним появлением нервной системы, более 500 миллионов лет назад, в кембрийский период. Дофамин действует как нейромедиатор у позвоночных , иглокожих , членистоногих , моллюсков и некоторых видов червей . [136] [137]
Было замечено, что у каждого типа животных, которые были исследованы, дофамин изменяет двигательное поведение. [133] В модельном организме , нематоде Caenorhabditis elegans , он снижает локомоцию и увеличивает движения, связанные с поиском пищи; у плоских червей совершает «винтовые» движения; у пиявок он тормозит плавание и способствует ползанию. У широкого круга позвоночных дофамин оказывает «активирующее» влияние на переключение поведения и выбор реакции, сравнимое с его действием у млекопитающих. [133] [138]
Также неоднократно доказывалось, что дофамин играет роль в обучении с вознаграждением во всех группах животных. [133] Как и всех позвоночных, беспозвоночных, таких как круглые черви , плоские черви , моллюски и обыкновенные плодовые мухи , можно научить повторять действие, если за ним постоянно следует повышение уровня дофамина. [133] У плодовых мух отдельные элементы обучения вознаграждению предполагают модульную структуру системы обработки вознаграждения насекомых, которая во многом аналогична структуре системы млекопитающих. [139] Например, дофамин регулирует краткосрочное и долгосрочное обучение обезьян; [140] у плодовых мух разные группы дофаминовых нейронов передают сигналы вознаграждения за кратковременную и долговременную память. [141]
Долгое время считалось, что членистоногие являются исключением из этого правила, а дофамин оказывает неблагоприятное воздействие. Было замечено, что вознаграждение вместо этого опосредовано октопамином , нейротрансмиттером, тесно связанным с норэпинефрином. [142] Однако более поздние исследования показали, что дофамин действительно играет роль в обучении с помощью вознаграждения у плодовых мух. Также было обнаружено, что полезный эффект октопамина обусловлен его активацией ряда дофаминергических нейронов, ранее не исследованных в исследованиях. [142]
Растения [ править ]

Многие растения, в том числе разнообразные пищевые, в разной степени синтезируют дофамин. [143] Самые высокие концентрации наблюдались в бананах: мякоть красных и желтых бананов содержит дофамин в количестве от 40 до 50 частей на миллион по весу. [143] Картофель, авокадо, брокколи и брюссельская капуста также могут содержать дофамин в количестве 1 часть на миллион или более; апельсины, помидоры, шпинат, фасоль и другие растения содержат измеримые концентрации менее 1 части на миллион. [143] Дофамин в растениях синтезируется из аминокислоты тирозина по биохимическим механизмам, аналогичным тем, которые используют животные. [143] Он может метаболизироваться различными способами с образованием меланина и различных алкалоидов в качестве побочных продуктов. [143] Функции растительных катехоламинов четко не установлены, но есть свидетельства того, что они играют роль в реакции на стрессоры, такие как бактериальная инфекция, в некоторых ситуациях действуют как факторы, стимулирующие рост, и изменяют способ метаболизма сахаров. Рецепторы, опосредующие эти действия, еще не идентифицированы, как и внутриклеточные механизмы, которые они активируют. [143]
Дофамин, потребляемый с пищей, не может действовать на мозг, поскольку не может преодолеть гематоэнцефалический барьер. [27] Однако есть также множество растений, которые содержат L-ДОФА, метаболический предшественник дофамина. [144] Самые высокие концентрации обнаруживаются в листьях и стручках растений рода Mucuna , особенно в Mucuna pruriens (бархатные бобы), которые использовались в качестве источника L-ДОФА в качестве лекарственного средства. [145] Еще одним растением, содержащим значительное количество L-ДОФА, является Vicia faba , растение, производящее конские бобы (также известные как «бобы»). Однако уровень L-ДОФА в бобах намного ниже, чем в оболочках стручков и других частях растения. [146] Семена деревьев кассии и баухинии также содержат значительное количество L-ДОФА. [144]
В одном из видов морских зеленых водорослей Ulvaria obscura , основном компоненте цветения некоторых водорослей , дофамин присутствует в очень высоких концентрациях, оцениваемых в 4,4% от сухого веса. Есть доказательства того, что этот дофамин действует как защита от травоядных животных , снижая потребление его улитками и изоподами . [147]
Как предшественник меланина [ править ]
Меланины — это семейство темно-пигментированных веществ, обнаруженных в широком спектре организмов. [148] Химически они тесно связаны с дофамином, и существует тип меланина, известный как дофамин-меланин , который может быть синтезирован путем окисления дофамина с помощью фермента тирозиназы . [148] Меланин, который затемняет кожу человека, не относится к этому типу: он синтезируется по пути, в котором в качестве предшественника используется L-ДОФА, а не дофамин. [148] Однако имеются убедительные доказательства того, что нейромеланин , придающий темный цвет черной субстанции мозга, по крайней мере частично представляет собой дофамин-меланин. [149]
Меланин, производный дофамина, вероятно, появляется по крайней мере в некоторых других биологических системах. Некоторая часть дофамина в растениях, вероятно, будет использоваться в качестве предшественника дофамина-меланина. [150] Считается, что сложные узоры, которые появляются на крыльях бабочек, а также черно-белые полосы на телах личинок насекомых, вызваны пространственно структурированными скоплениями дофамина-меланина. [151]
и развитие История
Дофамин был впервые синтезирован в 1910 году Джорджем Баргером и Джеймсом Юэнсом в лабораториях Wellcome в Лондоне, Англия. [152] и впервые идентифицирован в человеческом мозге Кэтрин Монтегю потому что это моноамин леводопа - в 1957 году . L , предшественником которого в синтезе Баргера-Юэнса является 3,4-дигидроксифенилаланин или Он был назван дофамином , ( ДОФА). . Функция дофамина как нейромедиатора была впервые признана в 1958 году Арвидом Карлссоном и Нильсом-Оке Хилларпом в Лаборатории химической фармакологии Национального института сердца Швеции . [153] Карлссон был удостоен Нобелевской премии по физиологии и медицине 2000 года за доказательство того, что дофамин является не только предшественником норадреналина (норадреналина) и адреналина (адреналина), но и сам по себе является нейромедиатором. [154]
Полидофамин [ править ]
Исследования, посвященные адгезивным полифенольным белкам мидий, привели к открытию в 2007 году, что самые разнообразные материалы, если их поместить в раствор дофамина со слабоосновным pH , покроются слоем полимеризованного дофамина, часто называемого полидофамином . [155] [156] Этот полимеризованный дофамин образуется в результате спонтанной реакции окисления и формально является разновидностью меланина. [157] Кроме того, самополимеризацию дофамина можно использовать для модуляции механических свойств гелей на основе пептидов. [158] Синтез полидофамина обычно включает реакцию гидрохлорида дофамина с Трисом в качестве основания в воде. Структура полидофамина неизвестна. [156]
Полидофаминовые покрытия могут образовываться на объектах размером от наночастиц до больших поверхностей. [157] Слои полидофамина обладают химическими свойствами, которые могут оказаться чрезвычайно полезными, и многочисленные исследования изучали их возможные применения. [157] На самом простом уровне их можно использовать для защиты от повреждения светом или для формирования капсул для доставки лекарств. [157] На более сложном уровне их адгезионные свойства могут сделать их полезными в качестве субстратов для биосенсоров или других биологически активных макромолекул. [157]
См. также [ править ]
Ссылки [ править ]
- ^ Круикшанк Л., Кеннеди А.Р., Шенкленд Н. (2013). «Вход в CSD TIRZAX: 5-(2-аммониоэтил)-2-гидроксифенолят, дофамин» . Кембриджская структурная база данных: структуры доступа . Кембриджский центр кристаллографических данных . дои : 10.5517/cc10m9nl .
- ^ Круикшанк Л., Кеннеди А.Р., Шенкленд Н. (2013). «Таутомерные и ионизационные формы дофамина и тирамина в твердом состоянии». Дж. Мол. Структура. 1051 : 132–36. Бибкод : 2013JMoSt1051..132C . doi : 10.1016/j.molstruc.2013.08.002 .
- ^ Jump up to: Перейти обратно: а б с д «Дофамин: Биологическая активность» . Руководство IUPHAR/BPS по фармакологии . Международный союз фундаментальной и клинической фармакологии . Проверено 29 января 2016 г.
- ^ Берридж К.К. (апрель 2007 г.). «Дебаты о роли дофамина в вознаграждении: аргументы в пользу значимости стимулов». Психофармакология . 191 (3): 391–431. дои : 10.1007/s00213-006-0578-x . ПМИД 17072591 . S2CID 468204 .
- ^ Jump up to: Перейти обратно: а б Уайз Р.А., Роббл М.А. (январь 2020 г.). «Дофамин и зависимость» . Ежегодный обзор психологии . 71 (1): 79–106. doi : 10.1146/annurev-psych-010418-103337 . ПМИД 31905114 . S2CID 210043316 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). Сидор А., Браун Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 147–48, 366–67, 375–76. ISBN 978-0-07-148127-4 .
- ^ Балики М.Н., Мансур А., Бария А.Т., Хуанг Л., Бергер С.Е., Филдс Х.Л., Апкариан А.В. (октябрь 2013 г.). «Разделение прилежащего человека на предполагаемое ядро и оболочку диссоциирует кодирование ценностей вознаграждения и боли» . Журнал неврологии . 33 (41): 16383–93. doi : 10.1523/JNEUROSCI.1731-13.2013 . ПМЦ 3792469 . ПМИД 24107968 .
- ^ Jump up to: Перейти обратно: а б Венцель Дж. М., Раушер Н. А., Чер Дж. Ф., Олесон Э. Б. (январь 2015 г.). «Роль фазового высвобождения дофамина в прилежащем ядре в кодировании отвращения: обзор нейрохимической литературы» . ACS Химическая нейронаука . 6 (1): 16–26. дои : 10.1021/cn500255p . ПМЦ 5820768 . ПМИД 25491156 .
Таким образом, стимулы, вызывающие страх, способны по-разному изменять фазовую передачу дофамина в субрегионах NAcc. Авторы предполагают, что наблюдаемое увеличение дофамина оболочки NAcc, вероятно, отражает общую мотивационную значимость, возможно, из-за облегчения состояния страха, вызванного CS, когда не проводится УЗИ (шок для ног). Это рассуждение подтверждается отчетом Будыгина и его коллег. 112 показывая, что у анестезированных крыс прекращение зажима хвоста приводит к увеличению высвобождения дофамина в панцире.
- ^ Пуглиси-Аллегра С., Вентура Р. (июнь 2012 г.). «Префронтальная/аккумбальная система катехоламинов обеспечивает высокую мотивационную значимость» . Передний. Поведение. Нейроски . 6:31 . дои : 10.3389/fnbeh.2012.00031 . ПМК 3384081 . ПМИД 22754514 .
- ^ Олесон Э.Б., Беккерт М.В., Морра Дж.Т., Лансинк К.С., Качопе Р., Абдулла Р.А. и др. (январь 2012 г.). «Эндоканнабиноиды формируют аккумбальное кодирование сигнально-мотивированного поведения посредством активации рецептора CB1 в вентральной покрышке» . Нейрон . 73 (2): 360–373. дои : 10.1016/j.neuron.2011.11.018 . ПМК 3269037 . ПМИД 22284189 .
- ^ Монкрифф Дж (2008). Миф о химическом лечении. Критика психиатрического лечения наркозависимости . Бейзингсток, Великобритания: Пэлгрейв Макмиллан. ISBN 978-0-230-57432-8 .
- ^ Волков Н.Д., Ван Г.Дж., Коллинз С.Х., Вигал Т.Л., Ньюкорн Дж.Х., Теланг Ф., Фаулер Дж.С., Чжу В., Логан Дж., Ма Ю., Прадхан К., Вонг С., Суонсон Дж.М. (сентябрь 2009 г.). «Оценка пути вознаграждения дофамина при СДВГ: клинические последствия» . ДЖАМА . 302 (10): 1084–91. дои : 10.1001/jama.2009.1308 . ПМЦ 2958516 . ПМИД 19738093 .
- ^ «Вливание дофамина» (PDF) . Проверено 13 октября 2023 г.
- ^ Jump up to: Перейти обратно: а б «Шок и гипотония у новорожденных. Лекарства: альфа/бета-адренергические агонисты, вазодилататоры, инотропные средства, расширители объема, антибиотики и другие» . emedicine.medscape.com . Проверено 13 октября 2023 г.
- ^ «Дофамин» . ПабХим . Проверено 21 сентября 2015 г.
- ^ «Катехоламин» . Британника . Проверено 21 сентября 2015 г.
- ^ «Фенилэтиламин» . ChemicalLand21.com . Проверено 21 сентября 2015 г.
- ^ Jump up to: Перейти обратно: а б с д и ж Картер Дж. Э., Джонсон Дж. Х., Бааске Д. М. (1982). «Дофамина гидрохлорид». Аналитические профили лекарственных веществ . 11 : 257–72. дои : 10.1016/S0099-5428(08)60266-X . ISBN 978-0122608117 .
- ^ «Технический лист» . www.sigmaaldrich.com . Проверено 13 сентября 2019 г.
- ^ Jump up to: Перейти обратно: а б с Бродли К.Дж. (март 2010 г.). «Сосудистые эффекты следовых аминов и амфетаминов». Фармакология и терапия . 125 (3): 363–375. doi : 10.1016/j.pharmthera.2009.11.005 . ПМИД 19948186 .
- ^ Jump up to: Перейти обратно: а б с Линдеманн Л., Хонер MC (май 2005 г.). «Ренессанс следовых аминов, вдохновленный новым семейством GPCR». Тенденции в фармакологических науках . 26 (5): 274–281. дои : 10.1016/j.tips.2005.03.007 . ПМИД 15860375 .
- ^ Jump up to: Перейти обратно: а б с д Ван X, Ли Дж, Донг Дж, Юэ Дж (февраль 2014 г.). «Эндогенные субстраты CYP2D мозга». Европейский журнал фармакологии . 724 : 211–218. дои : 10.1016/j.ejphar.2013.12.025 . ПМИД 24374199 .
- ^ Jump up to: Перейти обратно: а б с д и ж Симан П. (2009). «Глава 1: Исторический обзор: Знакомство с дофаминовыми рецепторами». В Неве К. (ред.). Дофаминовые рецепторы . Спрингер. стр. 1–22. ISBN 978-1-60327-333-6 .
- ^ «EC 1.14.16.2 – Тирозин-3-монооксигеназа (Homo sapiens)» . БРЕНДА . Технический университет Брауншвейга. июль 2016 года . Проверено 7 октября 2016 г.
Субстраты: L-фенилаланин + тетрагидробиоптерин + O2.
Продукт: L-тирозин + 3-гидроксифенилаланин [(он же м-тирозин)] + дигидроптеридин + H2O.
Организм: Человек разумный.
Диаграмма реакции - ^ «EC 4.1.1.28 – Декарбоксилаза ароматических L-аминокислот (Homo sapiens)» . БРЕНДА . Технический университет Брауншвейга. июль 2016 года . Проверено 7 октября 2016 г.
Субстрат: м-тирозин
Продукт: м-тирамин + CO2.
Организм: Человек разумный.
Диаграмма реакции - ^ Jump up to: Перейти обратно: а б с д и ж г час я дж Мусаккио Дж. М. (2013). «Глава 1: Ферменты, участвующие в биосинтезе и деградации катехоламинов». В Айверсоне Л. (ред.). Биохимия биогенных аминов . Спрингер. стр. 1–35. ISBN 978-1-4684-3171-1 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к Национальный сотрудничающий центр хронических состояний, изд. (2006). «Симптоматическая фармакологическая терапия при болезни Паркинсона» . Болезнь Паркинсона . Лондон: Королевский колледж врачей. стр. 59–100. ISBN 978-1-86016-283-1 . Архивировано из оригинала 24 сентября 2010 года . Проверено 24 сентября 2015 г.
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к Эйзенхофер Г., Копин И.Ю., Гольдштейн Д.С. (сентябрь 2004 г.). «Метаболизм катехоламинов: современный взгляд с последствиями для физиологии и медицины». Фармакологические обзоры . 56 (3): 331–49. дои : 10.1124/пр.56.3.1 . ПМИД 15317907 . S2CID 12825309 .
- ^ Захур I, Шафи А, Хак Э (декабрь 2018 г.). «Фармакологическое лечение болезни Паркинсона: Рисунок 1: [Метаболический путь синтеза дофамина...]» . В Stoker TB, Greenland JC (ред.). Болезнь Паркинсона: патогенез и клинические аспекты [Интернет] . Брисбен (Австралия): Публикации кодонов.
- ^ Амин Ф., Дэвидсон М., Дэвис К.Л. (1992). «Измерение гомованилиновой кислоты в клинических исследованиях: обзор методологии» . Бюллетень шизофрении . 18 (1): 123–48. дои : 10.1093/schbul/18.1.123 . ПМИД 1553492 .
- ^ Амин Ф., Дэвидсон М., Кан Р.С., Шмейдлер Дж., Стерн Р., Нотт П.Дж., Аптер С. (1995). «Оценка центрального дофаминергического индекса ГВА плазмы при шизофрении» . Бюллетень шизофрении . 21 (1): 53–66. дои : 10.1093/schbul/21.1.53 . ПМИД 7770741 .
- ^ Зульцер Д., Зекка Л. (февраль 2000 г.). «Внутринейронный синтез дофамин-хинона: обзор». Исследования нейротоксичности . 1 (3): 181–95. дои : 10.1007/BF03033289 . ПМИД 12835101 . S2CID 21892355 .
- ^ Миядзаки I, Асанума М (июнь 2008 г.). «Окислительный стресс, специфичный для дофаминергических нейронов, вызванный самим дофамином» (PDF) . Акта Медика Окаяма . 62 (3): 141–50. дои : 10.18926/AMO/30942 . ПМИД 18596830 .
- ^ Jump up to: Перейти обратно: а б с д и Гранди Д.К., Миллер ГМ, Ли Дж.С. (февраль 2016 г.). « «TAARgeting наркомания» - Аламо является свидетелем еще одной революции: обзор пленарного симпозиума Конференции по поведению, биологии и химии 2015 года» . Наркотическая и алкогольная зависимость . 159 : 9–16. doi : 10.1016/j.drugalcdep.2015.11.014 . ПМЦ 4724540 . ПМИД 26644139 .
TAAR1 представляет собой рецептор с высоким сродством к METH/AMPH и DA.
- ^ Jump up to: Перейти обратно: а б с д Романелли Р.Дж., Уильямс Дж.Т., Нив К.А. (2009). «Глава 6: Передача сигналов дофаминовых рецепторов: внутриклеточные пути поведения». В Неве К.А. (ред.). Дофаминовые рецепторы . Спрингер. стр. 137–74. ISBN 978-1-60327-333-6 .
- ^ Jump up to: Перейти обратно: а б Эйден Л.Е., Шефер М.К., Вейхе Э., Шютц Б. (февраль 2004 г.). «Семейство везикулярных переносчиков аминов (SLC18): антипортеры аминов / протонов, необходимые для везикулярного накопления и регулируемой экзоцитотической секреции моноаминов и ацетилхолина». Архив Пфлюгерса . 447 (5): 636–40. дои : 10.1007/s00424-003-1100-5 . ПМИД 12827358 . S2CID 20764857 .
- ^ Jump up to: Перейти обратно: а б Миллер GM (январь 2011 г.). «Новая роль рецептора 1, связанного с следами аминов, в функциональной регуляции переносчиков моноаминов и дофаминергической активности» . Журнал нейрохимии . 116 (2): 164–76. дои : 10.1111/j.1471-4159.2010.07109.x . ПМК 3005101 . ПМИД 21073468 .
- ^ Jump up to: Перейти обратно: а б Болье Ж.М., Гайнетдинов Р.Р. (март 2011 г.). «Физиология, передача сигналов и фармакология дофаминовых рецепторов». Фармакологические обзоры . 63 (1): 182–217. дои : 10.1124/пр.110.002642 . ПМИД 21303898 . S2CID 2545878 .
- ^ Торрес Г.Е., Гайнетдинов Р.Р., Кэрон М.Г. (январь 2003 г.). «Переносчики моноаминов плазматической мембраны: структура, регуляция и функции». Обзоры природы. Нейронаука . 4 (1): 13–25. дои : 10.1038/nrn1008 . ПМИД 12511858 . S2CID 21545649 .
- ^ Jump up to: Перейти обратно: а б с д Райс М.Э., Патель Дж.К., Крэгг С.Дж. (декабрь 2011 г.). «Выброс дофамина в базальных ганглиях» . Нейронаука . 198 : 112–37. doi : 10.1016/j.neuroscience.2011.08.066 . ПМК 3357127 . ПМИД 21939738 .
- ^ Шульц В. (2007). «Множественные функции дофамина в разное время». Ежегодный обзор неврологии . 30 : 259–88. дои : 10.1146/annurev.neuro.28.061604.135722 . ПМИД 17600522 . S2CID 13503219 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час Бьорклунд А., Даннетт С.Б. (май 2007 г.). «Дофаминовые нейронные системы головного мозга: обновление». Тенденции в нейронауках . 30 (5): 194–202. дои : 10.1016/j.tins.2007.03.006 . ПМИД 17408759 . S2CID 14239716 .
- ^ Jump up to: Перейти обратно: а б Дальстрём А, Фукс К (1964). «Доказательства существования моноаминсодержащих нейронов в центральной нервной системе. I. Демонстрация моноаминов в телах клеток нейронов ствола головного мозга». Acta Physiologica Scandinavica. Дополнение . 232 (Приложение): 1–55. ПМИД 14229500 .
- ^ Jump up to: Перейти обратно: а б с д Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 6: Широко распространенные системы: моноамины, ацетилхолин и орексин». В Сидоре А., Брауне Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 147–48, 154–57. ISBN 978-0-07-148127-4 .
- ^ Кристин К.В., Аминофф MJ (сентябрь 2004 г.). «Клиническая дифференциация паркинсонических синдромов: прогностическая и терапевтическая значимость». Американский медицинский журнал . 117 (6): 412–19. doi : 10.1016/j.amjmed.2004.03.032 . ПМИД 15380498 .
- ^ Фадок Дж.П., Дикерсон Т.М., Пальмитер Р.Д. (сентябрь 2009 г.). «Дофамин необходим для обусловленного сигналом обусловливания страха» . Журнал неврологии . 29 (36): 11089–97. doi : 10.1523/JNEUROSCI.1616-09.2009 . ПМК 2759996 . ПМИД 19741115 .
- ^ Тан В., Кочубей О., Кинчер М., Шнеггенбургер Р. (апрель 2020 г.). «ВТА на проекцию дофамина в базальной миндалевидном теле способствует передаче сигналов о значимых соматосенсорных событиях во время обучения страху» . Журнал неврологии . 40 (20): JN–RM–1796–19. doi : 10.1523/JNEUROSCI.1796-19.2020 . ПМЦ 7219297 . ПМИД 32277045 .
- ^ Джо Ю.С., Хейманн Г., Цвайфель Л.С. (ноябрь 2018 г.). «Дофаминовые нейроны отражают неопределенность при генерализации страха» . Нейрон . 100 (4): 916–925.e3. дои : 10.1016/j.neuron.2018.09.028 . ПМК 6226002 . ПМИД 30318411 .
- ^ Jump up to: Перейти обратно: а б Паулюс В., Шомбург Э.Д. (июнь 2006 г.). «Дофамин и спинной мозг при синдроме беспокойных ног: обнаруживает ли физиология спинного мозга основу для увеличения?». Обзоры медицины сна . 10 (3): 185–96. дои : 10.1016/j.smrv.2006.01.004 . ПМИД 16762808 .
- ^ Jump up to: Перейти обратно: а б с д и Бен-Джонатан Н., Хнаско Р. (декабрь 2001 г.). «Дофамин как ингибитор пролактина (ПРЛ)» . Эндокринные обзоры . 22 (6): 724–63. дои : 10.1210/er.22.6.724 . ПМИД 11739329 .
- ^ Jump up to: Перейти обратно: а б с д Витковский П. (январь 2004 г.). «Дофамин и функция сетчатки» . Документа Офтальмологическая. Достижения офтальмологии . 108 (1): 17–40. дои : 10.1023/B:DOOP.0000019487.88486.0a . ПМИД 15104164 . S2CID 10354133 .
- ^ Jump up to: Перейти обратно: а б Исправьте Джей Ди (2008). «Базальные ганглии и полосатая моторная система». Нейроанатомия (серия обзоров совета директоров) (4-е изд.). Балтимор: Вултерс Клювер и Липпинкотт Уильямс и Уилкинс. стр. 274–81. ISBN 978-0-7817-7245-7 .
- ^ Jump up to: Перейти обратно: а б с д и ж Чакраварти В.С., Джозеф Д., Бапи Р.С. (сентябрь 2010 г.). «Что делают базальные ганглии? Перспектива моделирования» . Биологическая кибернетика . 103 (3): 237–53. дои : 10.1007/s00422-010-0401-y . ПМИД 20644953 . S2CID 853119 .
- ^ Jump up to: Перейти обратно: а б с д Флореско СБ (январь 2015 г.). «Прилежащее ядро: интерфейс между познанием, эмоциями и действием» . Ежегодный обзор психологии . 66 : 25–52. doi : 10.1146/annurev-psych-010213-115159 . ПМИД 25251489 . S2CID 28268183 .
- ^ Jump up to: Перейти обратно: а б Баллейн Б.В., Дезфули А., Ито М., Дойя К. (2015). «Иерархический контроль целенаправленных действий в сети корково-базальных ганглиев». Современное мнение в области поведенческих наук . 5 : 1–7. дои : 10.1016/j.cobeha.2015.06.001 . S2CID 53148662 .
- ^ Jump up to: Перейти обратно: а б с Янкович Дж. (апрель 2008 г.). «Болезнь Паркинсона: клиника и диагностика» . Журнал неврологии, нейрохирургии и психиатрии . 79 (4): 368–76. дои : 10.1136/jnnp.2007.131045 . ПМИД 18344392 .
- ^ Паттидж Т., Вандершурен LJ (апрель 2008 г.). «Нейрофармакология импульсивного поведения» . Тенденции в фармакологических науках . 29 (4): 192–99. дои : 10.1016/j.tips.2008.01.002 . ПМИД 18304658 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к л м Шульц В. (июль 2015 г.). «Нейронное вознаграждение и сигналы решения: от теорий к данным» . Физиологические обзоры . 95 (3): 853–951. doi : 10.1152/physrev.00023.2014 . ПМЦ 4491543 . ПМИД 26109341 .
- ^ Jump up to: Перейти обратно: а б с Робинсон Т.Е., Берридж К.К. (1993). «Нейронная основа тяги к наркотикам: теория наркозависимости, стимулирующая сенсибилизацию». Исследования мозга. Обзоры исследований мозга . 18 (3): 247–91. дои : 10.1016/0165-0173(93)90013-п . hdl : 2027.42/30601 . ПМИД 8401595 . S2CID 13471436 .
- ^ Райт Дж.С., Панксепп Дж. (2012). «Эволюционная основа для понимания добывания пищи, желания и желания: нейропсихология системы ПОИСКА» . Нейропсихоанализ . 14 (1): 5–39. дои : 10.1080/15294145.2012.10773683 . S2CID 145747459 . Проверено 24 сентября 2015 г.
- ^ Jump up to: Перейти обратно: а б с д и Берридж К.К., Робинсон Т.Э., Олдридж Дж.В. (февраль 2009 г.). «Анализ компонентов вознаграждения: «нравится», «желать» и учиться» . Современное мнение в фармакологии . 9 (1): 65–73. дои : 10.1016/j.coph.2008.12.014 . ПМК 2756052 . ПМИД 19162544 .
- ^ Монтегю П.Р., Даян П., Сейновски Т.Дж. (март 1996 г.). «Система мезэнцефалических дофаминовых систем, основанная на прогнозирующем обучении Хебба» . Журнал неврологии . 16 (5): 1936–47. doi : 10.1523/JNEUROSCI.16-05-01936.1996 . ПМК 6578666 . ПМИД 8774460 .
- ^ Бромберг-Мартин Э.С., Мацумото М., Хикосака О. (декабрь 2010 г.). «Дофамин в мотивационном контроле: вознаграждение, отвращение и предупреждение» . Нейрон . 68 (5): 815–34. дои : 10.1016/j.neuron.2010.11.022 . ПМК 3032992 . ПМИД 21144997 .
- ^ Ягер Л.М., Гарсия А.Ф., Вунш А.М., Фергюсон С.М. (август 2015 г.). «Все тонкости полосатого тела: роль в наркозависимости» . Нейронаука . 301 : 529–41. doi : 10.1016/j.neuroscience.2015.06.033 . ПМЦ 4523218 . ПМИД 26116518 .
- ^ Jump up to: Перейти обратно: а б Саддорис, член парламента, Каччапалья Ф, Вайтман Р.М., Карелли Р.М. (август 2015 г.). «Дифференциальная динамика высвобождения дофамина в ядре и оболочке прилежащего ядра выявляет дополнительные сигналы для прогнозирования ошибок и побудительной мотивации» . Журнал неврологии . 35 (33): 11572–82. doi : 10.1523/JNEUROSCI.2344-15.2015 . ПМК 4540796 . ПМИД 26290234 .
- ^ Берридж К.К., Крингельбах М.Л. (май 2015 г.). «Системы удовольствия в мозге» . Нейрон . 86 (3): 646–64. дои : 10.1016/j.neuron.2015.02.018 . ПМЦ 4425246 . ПМИД 25950633 .
- ^ Jump up to: Перейти обратно: а б с Мудрый РА (1996). «Наркотики, вызывающие привыкание, и награда за стимуляцию мозга». Ежегодный обзор неврологии . 19 : 319–40. дои : 10.1146/annurev.ne.19.030196.001535 . ПМИД 8833446 .
- ^ Мудрый РА (октябрь 2008 г.). «Дофамин и вознаграждение: гипотеза ангедонии 30 лет спустя» . Исследования нейротоксичности . 14 (2–3): 169–83. дои : 10.1007/BF03033808 . ПМК 3155128 . ПМИД 19073424 .
- ^ Ариас-Каррион О, Пеппель Э (2007). «Дофамин, обучение и поведение, направленное на вознаграждение» . Акта Нейробиол Эксп . 67 (4): 481–88. дои : 10.55782/ane-2007-1664 . ПМИД 18320725 .
- ^ Икемото С. (ноябрь 2007 г.). «Схема вознаграждения дофамина: две проекционные системы от вентральной части среднего мозга к комплексу прилежащего ядра и обонятельного бугорка» . Обзоры исследований мозга . 56 (1): 27–78. дои : 10.1016/j.brainresrev.2007.05.004 . ПМК 2134972 . ПМИД 17574681 .
- ^ Jump up to: Перейти обратно: а б Феррери Л., Мас-Эрреро Э., Заторре Р.Х., Риполлес П., Гомес-Андрес А., Аликарт Х., Оливе Г., Марко-Пальярес Х., Антонихоан Р.М., Валле М., Риба Х., Родригес-Форнеллс А. (2019). «Дофамин модулирует вознаграждение, вызываемое музыкой» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (9): 3793–98. Бибкод : 2019PNAS..116.3793F . дои : 10.1073/pnas.1811878116 . ПМК 6397525 . ПМИД 30670642 .
Прослушивание приятной музыки часто сопровождается измеримыми телесными реакциями, такими как мурашки по коже или озноб, обычно называемые «ознобом» или «дрожью». ... В целом, наши результаты прямо показали, что фармакологические вмешательства двунаправленно модулируют реакции вознаграждения, вызываемые музыкой. В частности, мы обнаружили, что рисперидон ухудшал способность участников испытывать музыкальное удовольствие, тогда как леводопа усиливала ее. ... Здесь, напротив, изучая реакции на абстрактные вознаграждения у людей, мы показываем, что манипуляция дофаминергической передачей влияет как на удовольствие (т.е. количество времени, в течение которого сообщается о ознобе, и эмоциональное возбуждение, измеряемое EDA), так и на мотивационные компоненты музыкального вознаграждения. (деньги готовы потратить). Эти данные позволяют предположить, что дофаминергическая передача сигналов является непременным условием не только для мотивационных реакций, как было показано на примере первичных и вторичных вознаграждений, но также и для гедонистических реакций на музыку. Этот результат подтверждает недавние результаты, показывающие, что дофамин также опосредует восприятие приятности, достигаемой от других типов абстрактных вознаграждений (37), и бросает вызов предыдущим данным на животных моделях о первичных вознаграждениях, таких как еда (42, 43).
- ^ Jump up to: Перейти обратно: а б Гупиль Л., Окутюрье Ж.Ж. (февраль 2019 г.). «Музыкальное удовольствие и музыкальные эмоции» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (9): 3364–66. Бибкод : 2019PNAS..116.3364G . дои : 10.1073/pnas.1900369116 . ПМК 6397567 . ПМИД 30770455 .
В фармакологическом исследовании, опубликованном в PNAS, Ferreri et al. (1) представили доказательства того, что усиление или ингибирование передачи сигналов дофамина с помощью леводопы или рисперидона модулирует удовольствие, испытываемое при прослушивании музыки. ... В последней попытке установить не только корреляционную, но и причинно-следственную связь дофамина с музыкальным удовольствием, авторы обратились к прямому манипулированию дофаминергической передачей сигналов в полосатом теле, сначала применяя возбуждающую и тормозящую транскраниальную магнитную стимуляцию к мозгу участников. левая дорсолатеральная префронтальная кора, область, которая, как известно, модулирует функцию полосатого тела (5), и, наконец, в текущем исследовании, путем введения фармацевтических агентов, способных изменить доступность синапсов дофамина (1), оба из которых влияют на восприятие удовольствия, физиологические показатели возбуждения, и денежная стоимость, присвоенная музыке в предсказанном направлении. ... Хотя вопрос о музыкальном выражении эмоций имеет долгую историю исследований, в том числе в PNAS (6), а психофизиологическое направление исследований 1990-х годов уже установило, что музыкальное удовольствие может активировать вегетативную нервную систему (7), Демонстрация авторами влияния системы вознаграждения на музыкальные эмоции была воспринята как первое доказательство того, что это настоящие эмоции, исследование которых имеет полную законность для информирования нейробиологии о наших повседневных когнитивных, социальных и аффективных функциях (8). Кстати, это направление работы, кульминацией которого стала статья Феррери и др. (1), вероятно, сделал больше для привлечения финансирования исследований в области музыкальных наук, чем кто-либо другой в этом сообществе.
Доказательства Феррери и др. (1) обеспечивает новейшую поддержку убедительной нейробиологической модели, в которой музыкальное удовольствие возникает в результате взаимодействия древних систем вознаграждения/оценки (полосатого, лимбического и паралимбического) с более филогенетически развитыми системами восприятия/предсказания (темпорофронтальными). - ^ Кепп М.Дж., Ганн Р.Н., Лоуренс А.Д., Каннингем В.Дж., Дагер А., Джонс Т. и др. (май 1998 г.). «Доказательства выброса дофамина в полосатом теле во время видеоигры». Природа . 393 (6682): 266–268. Бибкод : 1998Natur.393..266K . дои : 10.1038/30498 . ПМИД 9607763 . S2CID 205000565 .
- ^ фон дер Хайден Дж. М., Браун Б., Мюллер К.В., Эглофф Б. (2019). «Связь между видеоиграми и психологическим функционированием» . Границы в психологии . 10 : 1731. дои : 10.3389/fpsyg.2019.01731 . ПМК 6676913 . ПМИД 31402891 .
- ^ Jump up to: Перейти обратно: а б Миссейл С., Нэш С.Р., Робинсон С.В., Джабер М., Кэрон М.Г. (январь 1998 г.). «Дофаминовые рецепторы: от структуры к функции» (PDF) . Физиологические обзоры . 78 (1): 189–225. дои : 10.1152/physrev.1998.78.1.189 . ПМИД 9457173 . S2CID 223462 . Архивировано из оригинала (PDF) 2 марта 2019 года.
- ^ Jump up to: Перейти обратно: а б Буттарелли ФР, Фанчулли А, Пелликано С, Понтьери ФЭ (июнь 2011 г.). «Дофаминергическая система в лимфоцитах периферической крови: от физиологии к фармакологии и возможному применению при нервно-психических расстройствах» . Современная нейрофармакология . 9 (2): 278–88. дои : 10.2174/157015911795596612 . ПМК 3131719 . ПМИД 22131937 .
- ^ Jump up to: Перейти обратно: а б Саркар С., Басу Б., Чакроборти Д., Дасгупта П.С., Басу С. (май 2010 г.). «Иммунорегулирующая роль дофамина: обновленная информация» . Мозг, поведение и иммунитет . 24 (4): 525–28. дои : 10.1016/j.bbi.2009.10.015 . ПМЦ 2856781 . ПМИД 19896530 .
- ^ Хусейн Т., Локхандвала М.Ф. (февраль 2003 г.). «Почечные дофаминовые рецепторы и гипертония». Экспериментальная биология и медицина . 228 (2): 134–42. дои : 10.1177/153537020322800202 . ПМИД 12563019 . S2CID 10896819 .
- ^ Чой М.Р., Куюмдзян Н.М., Рукавина Микусич Н.Л., Кравец М.К., Розон М.И., Родригес Фермепин М., Фернандес Б.Е. (май 2015 г.). «Почечная дофаминергическая система: патофизиологические последствия и клинические перспективы» . Всемирный журнал нефрологии . 4 (2): 196–212. дои : 10.5527/wjn.v4.i2.196 . ПМЦ 4419129 . ПМИД 25949933 .
- ^ Кэри Р.М. (сентябрь 2001 г.). «Лекция Теодора Купера: Почечная дофаминовая система: паракринный регулятор гомеостаза натрия и артериального давления» . Гипертония . 38 (3): 297–302. дои : 10.1161/hy0901.096422 . PMID 11566894 .
- ^ Jump up to: Перейти обратно: а б с д и Руби Б., Меклер П. (декабрь 2010 г.). «Мини-обзор: новая роль периферического дофамина в метаболическом контроле и росте опухолей: давайте искать баланс» . Эндокринология . 151 (12): 5570–81. дои : 10.1210/en.2010-0745 . ПМИД 21047943 .
- ^ «Примерный перечень основных лекарственных средств ВОЗ» (PDF) . Всемирная организация здравоохранения . Октябрь 2013 г. Архивировано (PDF) из оригинала 10 февраля 2014 г. Проверено 24 сентября 2015 г.
- ^ Нури С., Фридлих П., Сери I (2003). «Фармакологический обзор регулируемых сердечно-сосудистых, почечных и нейроэндокринных эффектов дофамина, связанных с развитием» . НеоОбзоры . 4 (10): с283–е288. дои : 10.1542/neo.4-10-e283 . S2CID 71902752 . Проверено 24 сентября 2015 г.
- ^ Jump up to: Перейти обратно: а б Бхатт-Мехта V, Нахата MC (1989). «Дофамин и добутамин в педиатрической терапии». Фармакотерапия . 9 (5): 303–14. дои : 10.1002/j.1875-9114.1989.tb04142.x . ПМИД 2682552 . S2CID 25614283 .
- ^ Jump up to: Перейти обратно: а б с Бронвен Дж.Б., Найтс КМ (2009). Фармакология для медицинских работников (2-е изд.). Эльзевир Австралия. п. 192. ИСБН 978-0-7295-3929-6 .
- ^ Де Бакер Д., Бистон П., Девриендт Дж., Мадл С., Чочрад Д., Альдекоа С. и др. (март 2010 г.). «Сравнение дофамина и норадреналина при лечении шока» (PDF) . Медицинский журнал Новой Англии . 362 (9): 779–89. doi : 10.1056/NEJMoa0907118 . ПМИД 20200382 . S2CID 2208904 . Архивировано из оригинала (PDF) 28 февраля 2019 года.
- ^ Картик С., Лиссабон А (2006). «Низкие дозы дофамина в отделении интенсивной терапии». Семинары по диализу . 19 (6): 465–71. дои : 10.1111/j.1525-139X.2006.00208.x . ПМИД 17150046 . S2CID 22538344 .
- ^ Мозес С. «Дофамин» . Тетрадь семейной практики . Проверено 1 февраля 2016 года .
- ^ Катрицис Д.Г., Герш Б.Дж., Камм А.Дж. (2013). Клиническая кардиология: Современные практические рекомендации . ОУП Оксфорд. ISBN 978-0-19-150851-6 .
Дофамин связывается с бета-1, бета-2, альфа-1 и дофаминергическими рецепторами.
- ^ Льюис Р.Дж. (2004). Опасные свойства промышленных материалов Сакса (11-е изд.). Хобокен, Нью-Джерси: Wiley & Sons. п. 1552. ИСБН 978-0-471-47662-7 .
- ^ Стандарт Д.Г., Уолш Р.Р. (2011). «Фармакология дофаминергической нейротрансмиссии». В Ташджиане А.Х., Армстронге Э.Дж., Голане Д.Э. (ред.). Принципы фармакологии: патофизиологические основы лекарственной терапии . Липпинкотт Уильямс и Уилкинс. стр. 186–206. ISBN 978-1-4511-1805-6 .
- ^ Резюме Моббса, Hof PR (2009). Справочник по нейробиологии старения . Амстердам: Elsevier/Academic Press. ISBN 978-0-12-374898-0 . OCLC 299710911 .
- ^ Ота М, Ясуно Ф, Ито Х, Секи С, Нодзаки С, Асада Т, Сухара Т (июль 2006 г.). «Возрастное снижение синтеза дофамина в мозге живого человека, измеренное с помощью позитронно-эмиссионной томографии с L-[бета-11C]ДОФА». Науки о жизни . 79 (8): 730–36. дои : 10.1016/j.lfs.2006.02.017 . ПМИД 16580023 .
- ^ Каасинен В., Вилкман Х., Хиетала Дж., Ногрен К., Хелениус Х., Олссон Х., Фарде Л., Ринне Дж. (2000). «Возрастная потеря дофаминовых рецепторов D2/D3 в экстрастриарных областях человеческого мозга». Нейробиология старения . 21 (5): 683–68. дои : 10.1016/S0197-4580(00)00149-4 . ПМИД 11016537 . S2CID 40871554 .
- ^ Ван Й., Чан Г.Л., Холден Дж.Э., Добко Т., Мак Э., Шульцер М., Хузер Дж.М., Сноу Б.Дж., Рут Т.Дж., Калн Д.Б., Штессл А.Дж. (сентябрь 1998 г.). «Возрастное снижение дофаминовых рецепторов D1 в мозге человека: исследование ПЭТ». Синапс . 30 (1): 56–61. doi : 10.1002/(SICI)1098-2396(199809)30:1<56::AID-SYN7>3.0.CO;2-J . ПМИД 9704881 . S2CID 31445572 .
- ^ Вонг Д.Ф., Вагнер Х.Н., Данналс Р.Ф., Линкс Дж.М., Фрост Дж.Дж., Раверт Х.Т., Уилсон А.А., Розенбаум А.Е., Гьедде А., Дуглас К.Х. (декабрь 1984 г.). «Влияние возраста на рецепторы дофамина и серотонина, измеренное с помощью позитронной томографии в мозгу живого человека». Наука . 226 (4681): 1393–96. Бибкод : 1984Sci...226.1393W . дои : 10.1126/science.6334363 . ПМИД 6334363 . S2CID 24278577 .
- ^ Jump up to: Перейти обратно: а б Ван Э, Снайдер С.Д. (1998). Справочник стареющего мозга . Сан-Диего, Калифорния: Academic Press. ISBN 978-0-12-734610-6 . OCLC 636693117 .
- ^ Добрякова Е., Дженова Х.М., ДеЛука Дж., Уайли Г.Р. (12 марта 2015 г.). «Гипотеза дисбаланса дофамина об усталости при рассеянном склерозе и других неврологических расстройствах» . Границы в неврологии . 6:52 . doi : 10.3389/fneur.2015.00052 . ПМЦ 4357260 . ПМИД 25814977 .
- ^ Марино Ф, Косентино М (апрель 2016 г.). «Рассеянный склероз: повторное использование дофаминергических препаратов для лечения рассеянного склероза - количество доказательств». Обзоры природы. Неврология . 12 (4): 191–92. дои : 10.1038/nrneurol.2016.33 . ПМИД 27020558 . S2CID 26319461 .
- ^ Диксон Д.В. (2007). «Невропатология двигательных расстройств». В Толосе Э., Янкович Дж. Дж. (ред.). Болезнь Паркинсона и двигательные расстройства . Хагерстаун, Мэриленд: Липпинкотт Уильямс и Уилкинс. стр. 271–83. ISBN 978-0-7817-7881-7 .
- ^ Jump up to: Перейти обратно: а б Туите П.Дж., Кравчевски К. (апрель 2007 г.). «Паркинсонизм: обзор системного подхода к диагностике». Семинары по неврологии . 27 (2): 113–22. дои : 10.1055/s-2007-971174 . ПМИД 17390256 . S2CID 260319916 .
- ^ Jump up to: Перейти обратно: а б Олсен CM (декабрь 2011 г.). «Естественные награды, нейропластичность и ненаркотическая зависимость» . Нейрофармакология . 61 (7): 1109–22. doi : 10.1016/j.neuropharm.2011.03.010 . ПМК 3139704 . ПМИД 21459101 .
- ^ Чераволо Р., Фросини Д., Росси С., Бонуччелли У. (ноябрь 2010 г.). «Спектр зависимостей при болезни Паркинсона: от синдрома дофаминовой дисрегуляции до нарушений контроля над импульсами». Журнал неврологии . 257 (Приложение 2): С276–83. дои : 10.1007/s00415-010-5715-0 . ПМИД 21080189 . S2CID 19277026 .
- ^ Jump up to: Перейти обратно: а б с д и ж г Годсе Х (2010). Наркотики Годсе и аддиктивное поведение: Руководство по лечению (4-е изд.). Издательство Кембриджского университета. стр. 87–92. ISBN 978-1-139-48567-8 .
- ^ Сицилиано, Калифорния, Джонс С.Р. (август 2017 г.). «Потенция кокаина в транспортере дофамина отслеживает дискретные мотивационные состояния во время самостоятельного приема кокаина» . Нейропсихофармакология . 42 (9): 1893–1904. дои : 10.1038/npp.2017.24 . ПМК 5520781 . ПМИД 28139678 .
- ^ Heal DJ, Пирс Д.М. (2006). «Метилфенидат и его изомеры: их роль в лечении синдрома дефицита внимания и гиперактивности с использованием системы трансдермальной доставки». Препараты ЦНС . 20 (9): 713–38. дои : 10.2165/00023210-200620090-00002 . ПМИД 16953648 . S2CID 39535277 .
- ^ Jump up to: Перейти обратно: а б Фрей Э (2009). Фармакология и злоупотребление кокаином, амфетамином, экстази и родственными дизайнерскими наркотиками: всесторонний обзор их механизма действия, лечения злоупотребления и интоксикации . Дордрехт: Спрингер. ISBN 978-90-481-2448-0 .
- ^ Jump up to: Перейти обратно: а б Кимко ХК, Кросс Дж.Т., Абернети Д.Р. (декабрь 1999 г.). «Фармакокинетика и клиническая эффективность метилфенидата». Клиническая фармакокинетика . 37 (6): 457–70. дои : 10.2165/00003088-199937060-00002 . ПМИД 10628897 . S2CID 397390 .
- ^ Миньо Э.Ж. (октябрь 2012 г.). «Практическое руководство по терапии синдромов нарколепсии и гиперсомнии» . Нейротерапия . 9 (4): 739–52. дои : 10.1007/s13311-012-0150-9 . ПМЦ 3480574 . ПМИД 23065655 .
- ^ Циммерман Дж.Л. (октябрь 2012 г.). «Кокаиновое опьянение». Клиники интенсивной терапии . 28 (4): 517–26. дои : 10.1016/j.ccc.2012.07.003 . ПМИД 22998988 .
- ^ «Quillivant XR – суспензия гидрохлорида метилфенидата пролонгированного действия» . dailymed.nlm.nih.gov . Проверено 11 июля 2020 г.
- ^ Лопес Ф.А., Леру-младший (сентябрь 2013 г.). «Стимуляторы длительного действия для лечения синдрома дефицита внимания и гиперактивности: акцент на составах пролонгированного действия и пролекарстве лиздексамфетамина димезилате для решения продолжающихся клинических проблем» . Синдром дефицита внимания и гиперактивности . 5 (3): 249–65. дои : 10.1007/s12402-013-0106-x . ПМЦ 3751218 . ПМИД 23564273 .
- ^ Натт DJ, Лингфорд-Хьюз А., Эрритцо Д., Стоукс PR (май 2015 г.). «Дофаминовая теория зависимости: 40 лет взлетов и падений» (PDF) . Обзоры природы. Нейронаука . 16 (5): 305–12. дои : 10.1038/nrn3939 . ПМИД 25873042 . S2CID 205511111 .
- ^ Jump up to: Перейти обратно: а б с Синха Р. (август 2013 г.). «Клиническая нейробиология тяги к наркотикам» . Современное мнение в нейробиологии . 23 (4): 649–54. дои : 10.1016/j.conb.2013.05.001 . ПМЦ 3735834 . ПМИД 23764204 .
- ^ Волков Н.Д., Балер Р.Д. (январь 2014 г.). «Наука о зависимостях: раскрытие нейробиологической сложности» . Нейрофармакология . 76 (Часть Б): 235–49. doi : 10.1016/j.neuropharm.2013.05.007 . ПМЦ 3818510 . ПМИД 23688927 .
- ^ Нестлер Э.Дж. (декабрь 2012 г.). «Транскрипционные механизмы наркомании» . Клиническая психофармакология и неврология . 10 (3): 136–43. дои : 10.9758/cpn.2012.10.3.136 . ПМК 3569166 . ПМИД 23430970 .
- ^ Йео Б.Т., Криенен Ф.М., Томб Дж., Сабунку М.Р., Лашкари Д., Холлинсхед М., Роффман Дж.Л., Смоллер Дж.В., Золлей Л., Полимени Дж.Р., Фишль Б., Лю Х., Бакнер Р.Л. (сентябрь 2011 г.). «Организация коры головного мозга человека по внутренней функциональной связности» . Журнал нейрофизиологии . 106 (3): 1125–65. дои : 10.1152/jn.00338.2011 . ПМК 3174820 . ПМИД 21653723 .
- ^ Jump up to: Перейти обратно: а б с д и ж г Хили Д. (2004). Создание психофармакологии . Издательство Гарвардского университета. стр. 37–73. ISBN 978-0-674-01599-9 .
- ^ Jump up to: Перейти обратно: а б Брантон Л. Гудман и «Фармакологические основы терапии» Гилмана (12-е изд.). МакГроу Хилл. стр. 417–55.
- ^ Jump up to: Перейти обратно: а б с д Хоуз О.Д., Капур С. (май 2009 г.). «Дофаминовая гипотеза шизофрении: версия III — последний общий путь» . Бюллетень шизофрении . 35 (3): 549–62. дои : 10.1093/schbul/sbp006 . ПМЦ 2669582 . ПМИД 19325164 .
- ^ Горачек Дж., Бубеникова-Валесова В., Копечек М., Паленичек Т., Докери С., Мор П., Хёшль С. (2006). «Механизм действия атипичных антипсихотических препаратов и нейробиология шизофрении». Препараты ЦНС . 20 (5): 389–409. дои : 10.2165/00023210-200620050-00004 . ПМИД 16696579 . S2CID 18226404 .
- ^ Шталь СМ (2018). «Помимо дофаминовой гипотезы шизофрении, существуют три нейронные сети психоза: дофамин, серотонин и глутамат» (PDF) . Спектр ЦНС . 23 (3): 187–91. дои : 10.1017/S1092852918001013 . ПМИД 29954475 . S2CID 49599226 . Архивировано (PDF) из оригинала 29 апреля 2020 г.
- ^ Jump up to: Перейти обратно: а б Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Главы 10 и 13». В Сидоре А., Брауне Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 266, 318–23. ISBN 978-0-07-148127-4 .
- ^ Ву Дж, Сяо Х, Сунь Х, Цзоу Л, Чжу LQ (июнь 2012 г.). «Роль дофаминовых рецепторов при СДВГ: систематический метаанализ». Молекулярная нейробиология . 45 (3): 605–20. дои : 10.1007/s12035-012-8278-5 . ПМИД 22610946 . S2CID 895006 .
- ^ Jump up to: Перейти обратно: а б Берридж CW, Devilbiss DM (июнь 2011 г.). «Психостимуляторы как усилители когнитивных функций: префронтальная кора, катехоламины и синдром дефицита внимания и гиперактивности» . Биологическая психиатрия . 69 (12): e101–11. doi : 10.1016/j.biopsych.2010.06.023 . ПМК 3012746 . ПМИД 20875636 .
- ^ Спенсер RC, Девилбисс DM, Берридж CW (июнь 2015 г.). «Эффекты психостимуляторов, улучшающие когнитивные функции, включают прямое воздействие на префронтальную кору» . Биологическая психиатрия . 77 (11): 940–50. doi : 10.1016/j.biopsych.2014.09.013 . ПМК 4377121 . ПМИД 25499957 .
- ^ Илиева И.П., Хук С.Дж., Фара М.Дж. (июнь 2015 г.). «Влияние рецептурных стимуляторов на здоровый тормозной контроль, рабочую память и эпизодическую память: метаанализ» . Журнал когнитивной нейронауки . 27 (6): 1069–89. дои : 10.1162/jocn_a_00776 . ПМИД 25591060 . S2CID 15788121 .
- ^ Jump up to: Перейти обратно: а б с Вуд ПБ (май 2008 г.). «Роль центрального дофамина в боли и анальгезии». Экспертный обзор нейротерапии . 8 (5): 781–97. дои : 10.1586/14737175.8.5.781 . ПМИД 18457535 . S2CID 24325199 .
- ^ Jump up to: Перейти обратно: а б с д Флейк З.А., Скалли Р.Д., Бейли А.Г. (март 2004 г.). «Практический выбор противорвотных средств» . Американский семейный врач . 69 (5): 1169–74. ПМИД 15023018 .
- ^ Коннолли Б.С., Ланг А.Э. (2014). «Фармакологическое лечение болезни Паркинсона: обзор». ДЖАМА . 311 (16): 1670–83. дои : 10.1001/jama.2014.3654 . ПМИД 24756517 .
- ^ Рощина В.В. (2010). «Эволюционные аспекты нейротрансмиттеров в микробных, растительных и животных клетках». В Lyte M, Primrose PE (ред.). Микробная эндокринология . Нью-Йорк: Спрингер. стр. 17–52. ISBN 978-1-4419-5576-0 .
- ^ Jump up to: Перейти обратно: а б Айер Л.М., Аравинд Л., Кун С.Л., Кляйн Д.С., Кунин Е.В. (июль 2004 г.). «Эволюция межклеточной передачи сигналов у животных: сыграл ли роль поздний горизонтальный перенос генов от бактерий?». Тенденции в генетике . 20 (7): 292–99. дои : 10.1016/j.tig.2004.05.007 . ПМИД 15219393 .
- ^ Jump up to: Перейти обратно: а б с д и Бэррон А.Б., Совик Э., Корниш Дж.Л. (2010). «Роль дофамина и родственных соединений в поведении, направленном на вознаграждение, среди животных разных типов» . Границы поведенческой нейронауки . 4 : 163. дои : 10.3389/fnbeh.2010.00163 . ПМЦ 2967375 . ПМИД 21048897 .
- ^ Лю Х., Мисима Ю., Фудзивара Т., Нагай Х., Китадзава А., Мине Ю. и др. (2004). «Выделение арагуспонгина М, нового стереоизомера алкалоида арагуспонгина/ксестоспонгина, и дофамина из морской губки Neopetrosia exigua, собранной на Палау» . Морские наркотики . 2 (4): 154–63. дои : 10.3390/md204154 . ПМЦ 3783253 .
- ^ Касс-Саймон Дж., Пьеробон П. (январь 2007 г.). «Книдарийская химическая нейротрансмиссия, обновленный обзор». Сравнительная биохимия и физиология. Часть A. Молекулярная и интегративная физиология . 146 (1): 9–25. дои : 10.1016/j.cbpa.2006.09.008 . ПМИД 17101286 .
- ^ Коттрелл Г.А. (январь 1967 г.). «Наличие дофамина и норадреналина в нервной ткани некоторых видов беспозвоночных» . Британский журнал фармакологии и химиотерапии . 29 (1): 63–69. дои : 10.1111/j.1476-5381.1967.tb01939.x . ПМК 1557178 . ПМИД 19108240 .
- ^ Киндт К.С., Кваст К.Б., Джайлз А.С., Де С., Хендри Д., Никастро И., Ранкин Ч.Х., Шафер В.Р. (август 2007 г.). «Дофамин опосредует контекстно-зависимую модуляцию сенсорной пластичности у C. elegans» . Нейрон . 55 (4): 662–76. дои : 10.1016/j.neuron.2007.07.023 . ПМИД 17698017 . S2CID 2092645 .
- ^ Каливас П.В., Стюарт Дж. (1 сентября 1991 г.). «Передача дофамина при инициировании и выражении сенсибилизации двигательной активности, вызванной лекарствами и стрессом». Исследования мозга. Обзоры исследований мозга . 16 (3): 223–44. дои : 10.1016/0165-0173(91)90007-У . ПМИД 1665095 . S2CID 10775295 .
- ^ Перри CJ, Barron AB (2013). «Нейральные механизмы вознаграждения у насекомых» (PDF) . Ежегодный обзор энтомологии . 58 (1): 543–62. doi : 10.1146/annurev-ento-120811-153631 . ПМИД 23020615 . S2CID 19678766 . Архивировано из оригинала (PDF) 5 июня 2020 года.
- ^ Такикава Ю., Кавагоэ Р., Хикосака О. (октябрь 2004 г.). «Возможная роль дофаминовых нейронов среднего мозга в краткосрочной и долгосрочной адаптации саккад к картированию положения-вознаграждения» (PDF) . Журнал нейрофизиологии . 92 (4): 2520–29. дои : 10.1152/jn.00238.2004 . ПМИД 15163669 . S2CID 12534057 . Архивировано из оригинала (PDF) 2 марта 2019 года.
- ^ Ямагата Н., Ичиносе Т., Асо Ю., Плас П.Ю., Фридрих А.Б., Сима Р.Дж. и др. (январь 2015 г.). «Отдельные дофаминовые нейроны передают сигналы вознаграждения для краткосрочной и долгосрочной памяти» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (2): 578–83. Бибкод : 2015PNAS..112..578Y . дои : 10.1073/pnas.1421930112 . ПМК 4299218 . ПМИД 25548178 .
- ^ Jump up to: Перейти обратно: а б Уодделл С. (июнь 2013 г.). «Передача сигналов подкрепления у дрозофилы; в конце концов, все это делает дофамин» . Современное мнение в нейробиологии . 23 (3): 324–29. дои : 10.1016/j.conb.2013.01.005 . ПМЦ 3887340 . ПМИД 23391527 .
- ^ Jump up to: Перейти обратно: а б с д и ж Кульма А., Шопа Дж. (2007). «Катехоламины являются активными соединениями растений». Наука о растениях . 172 (3): 433–40. doi : 10.1016/j.plantsci.2006.10.013 .
- ^ Jump up to: Перейти обратно: а б Ингл ПК (2003). «Подшипниковые растения L-ДОФА» (PDF) . Натуральный продукт: сияние . 2 : 126–33. Архивировано (PDF) из оригинала 2 марта 2014 г. Проверено 24 сентября 2015 г.
- ^ Вичерс Х.Дж., Виссер Дж.Ф., Хейзинг Х.Дж., Прас Н. (1993). «Наличие L-ДОФА и дофамина в растениях и культурах клеток Mucuna pruriens и влияние 2, 4-d и NaCl на эти соединения». Культура растительных клеток, тканей и органов . 33 (3): 259–64. дои : 10.1007/BF02319010 . S2CID 44814336 .
- ^ Лонго Р., Кастеллани А., Сберзе П., Тиболла М. (1974). «Распределение L-допы и родственных аминокислот в Vicia». Фитохимия . 13 (1): 167–71. Бибкод : 1974PChem..13..167L . дои : 10.1016/S0031-9422(00)91287-1 .
- ^ Ван Алстайн К.Л., Нельсон А.В., Вивиан-младший, канцлер Д.А. (июнь 2006 г.). «Дофамин действует как защита от травоядных животных у зеленой водоросли Ulvaria obscura умеренного климата». Экология 148 (2): 304–11. Бибкод : 2006Oecol.148..304V . дои : 10.1007/ s00442-006-0378-3 ПМИД 16489461 . S2CID 5029574 .
- ^ Jump up to: Перейти обратно: а б с Саймон Дж.Д., Пелеш Д., Вакамацу К., Ито С. (октябрь 2009 г.). «Текущие проблемы в понимании меланогенеза: химия мостов, биологический контроль, морфология и функции» . Исследование пигментных клеток и меланомы . 22 (5): 563–79. дои : 10.1111/j.1755-148X.2009.00610.x . ПМИД 19627559 .
- ^ Федороу Х, Трибл Ф, Холлидей Дж, Герлах М, Ридерер П, Дабл КЛ (февраль 2005 г.). «Нейромеланин в дофаминовых нейронах человека: сравнение с периферическими меланинами и значимость для болезни Паркинсона». Прогресс нейробиологии . 75 (2): 109–24. doi : 10.1016/j.pneurobio.2005.02.001 . ПМИД 15784302 . S2CID 503902 .
- ^ Эндрюс Р.С., Придэм Дж.Б. (1967). «Меланины из ДОФА-содержащих растений». Фитохимия . 6 (1): 13–18. Бибкод : 1967PChem...6...13A . дои : 10.1016/0031-9422(67)85002-7 .
- ^ Белдаде П., Брейкфилд, премьер-министр (июнь 2002 г.). «Генетика и эво-дево рисунков крыльев бабочек». Обзоры природы. Генетика . 3 (6): 442–52. дои : 10.1038/nrg818 . ПМИД 12042771 . S2CID 17417235 .
- ^ Фан С. (2008). «История применения дофамина и леводопы в лечении болезни Паркинсона». Двигательные расстройства . 23 (Приложение 3): S497–508. дои : 10.1002/mds.22028 . ПМИД 18781671 . S2CID 45572523 .
- ^ Бенеш FM (январь 2001 г.). «Карлссон и открытие дофамина». Тенденции в фармакологических науках . 22 (1): 46–47. дои : 10.1016/S0165-6147(00)01607-2 . ПМИД 11165672 .
- ^ Баронды С.Х. (2003). Лучше, чем Прозак . Нью-Йорк: Издательство Оксфордского университета. стр. 21–22, 39–40 . ISBN 978-0-19-515130-5 .
- ^ Ли Х, Деллаторе С.М., Миллер В.М., Мессерсмит П.Б. (октябрь 2007 г.). «Химия поверхности на основе мидий для многофункциональных покрытий» . Наука . 318 (5849): 426–30. Бибкод : 2007Sci...318..426L . дои : 10.1126/science.1147241 . ПМК 2601629 . ПМИД 17947576 .
- ^ Jump up to: Перейти обратно: а б Дрейер Д.Р., Миллер Д.Д., Фриман Б.Д., Пол Д.Р., Белявски К.В. (2013). «Перспективы поли(дофамина)». Химическая наука . 4 (10): 3796. doi : 10.1039/C3SC51501J .
- ^ Jump up to: Перейти обратно: а б с д и Люнг М.Е., ван дер Вестен Р., Постма А., Штедлер Б. (декабрь 2011 г.). «Полидофамин — природное полимерное покрытие для биомедицинских наук» (PDF) . Наномасштаб . 3 (12): 4916–28. Бибкод : 2011Nanos...3.4916L . дои : 10.1039/c1nr10969c . ПМИД 22024699 . Архивировано из оригинала 7 марта 2014 года.
- ^ Фихман Г., Шнайдер Дж. П. (март 2021 г.). «Самополимеризация дофамина как простой и мощный инструмент для модуляции вязкоупругих механических свойств гелей на основе пептидов» . Молекулы . 26 (5): 1363. doi : 10,3390/molecules26051363 . ПМЦ 7961423 . ПМИД 33806346 .
Внешние ссылки [ править ]
 Словарное определение дофамина в Викисловаре
Словарное определение дофамина в Викисловаре  СМИ, связанные с допамином, на Викискладе?
СМИ, связанные с допамином, на Викискладе?
- Дофамин
- Кардиостимуляторы
- Катехоламины
- Агонисты дофамина
- Гормоны гипоталамуса
- Гормоны гипоталамо-гипофизарно-пролактиновой оси
- Фенэтиламины
- Инотропные средства
- Мотивация
- Нейромедиаторы
- Агенты, высвобождающие норадреналин-дофамин
- Агонисты TAAR1
- Биология синдрома дефицита внимания с гиперактивностью
- Периферически селективные препараты
- Женская эндокринная система человека