Последняя область
| Последняя область | |
|---|---|
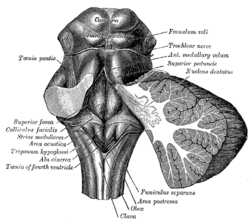 Ромбовидная ямка. (Область пострема отмечена внизу по центру.) | |
 Описание вида сзади каудального ствола мозга человека (Area postrema № 8) | |
| Подробности | |
| Часть | Медулла |
| Идентификаторы | |
| Акроним(ы) | АП |
| МеШ | Д031608 |
| Нейроимена | 772 |
| НейроЛекс ID | бирнлекс_2636 |
| ТА98 | A14.1.04.258 |
| ТА2 | 6009 |
| ФМА | 72607 |
| Анатомические термины нейроанатомии | |
Area postrema — парная структура в продолговатом мозге ствола мозга . [1] Это околожелудочковый орган, имеющий проницаемые капилляры и сенсорные нейроны , которые позволяют ему выполнять двойную роль: обнаруживать циркулирующие в крови химические посланники и преобразовывать их в нервные сигналы и сети . [2] [3] [4] Его расположение рядом с двусторонними ядрами одиночного тракта и роль сенсорного преобразователя позволяют ему интегрировать автономные функции крови и мозга . Такие функции области пострема включают обнаружение циркулирующих гормонов, участвующих в рвоте , жажде , голоде и контроле артериального давления . [1] [5]
Структура
[ редактировать ]Area postrema — парный выступ, расположенный у нижне-заднего края четвертого желудочка . [1] [5] специализированные эпендимальные клетки В области постремы обнаруживаются . Эти клетки незначительно отличаются от большинства эпендимальных клеток (эпендимоцитов), образующих одноклеточную эпителиальную выстилку желудочков и центрального канала . Area postrema отделена от треугольника блуждающего — нерва разделительным канатиком тонким полупрозрачным гребнем. [1] [5] Вагальный треугольник лежит над дорсальным ядром блуждающего нерва и расположен на каудальном конце ромбовидной ямки или «дна» четвертого желудочка. Область пострема расположена непосредственно перед обексом , нижней верхушкой дна каудального желудочка. И отделяющий канатик, и область постремы имеют одинаковое толстое покрытие таницитов, содержащее эпендиму. Эпендима и танициты могут участвовать в транспорте нейрохимических веществ и из в спинномозговую жидкость ее клеток или соседних нейронов, глии или сосудов. Эпендима и танициты также могут участвовать в хеморецепции. [1] [5]
Постремная область считается околожелудочковым органом из-за ее близости к желудочковой системе . [2] В морфологическом области постремы исследовании было показано, что капилляры в вентральной субобласти области постремы относительно непроницаемы, как и в мозге, тогда как капилляры медиальной и дорсальной области постремы имели микроскопические характеристики высокой проницаемости, характеристику, называемую синусоидальной . [6] Субрегиональная плотность капилляров области пострема была самой высокой вблизи границы желудочка и была почти в два раза выше, чем плотность капилляров соседнего одиночного ядра (SN) и дорсального двигательного ядра блуждающего нерва . [6] Таницитарный барьер частично компенсирует высокую проницаемость капилляров в области пострема. [7]
Физиологические субрегиональные исследования области постремы показали, что объем ее крови относительно велик, а кровоток и время прохождения маркеров крови относительно медленны, что усиливает способность восприятия циркулирующих соединений, таких как гормоны или трансмиттеры. [8]

Соединения
[ редактировать ]Область пострема соединяется с одиночным ядром или ядром солитарного тракта (NTS) и другими центрами автономного управления в стволе мозга . Он возбуждается висцеральными афферентными импульсами (симпатическими и вагусными), исходящими из желудочно-кишечного тракта и других периферических триггерных зон , а также гуморальными факторами . [2] Area postrema составляет часть дорсального вагусного комплекса, который является критическим местом окончания афферентных нервных волокон блуждающего нерва , наряду с дорсальным двигательным ядром блуждающего нерва и NTS.
Тошнота , скорее всего, вызывается стимуляцией постремной области через ее соединение с NTS, что может служить началом пути, вызывающего рвоту в ответ на различные рвотные воздействия . Однако эта структура не играет ключевой роли в возникновении тошноты, вызванной активацией волокон блуждающего нерва или движением, а ее функция при радиационно-индуцированной рвоте остается неясной. [9]
Поскольку область пострема и специализированная область НТС имеют проницаемые капилляры , [2] пептиды и другие гормональные сигналы в крови имеют прямой доступ к нейронам областей мозга, играющих жизненно важную роль в автономном контроле организма. [2] [6] В результате область пострема считается местом интеграции различных физиологических сигналов крови, когда они попадают в центральную нервную систему . [2] [3]
Функция
[ редактировать ]Хеморецепция
[ редактировать ]Area postrema — один из околожелудочковых органов . [10] обнаруживает токсины в крови и действует как центр, вызывающий рвоту. Постремная область является критическим гомеостатическим центром интеграции гуморальных и нервных сигналов благодаря своей функции триггерной зоны хеморецепторов рвоты в ответ на рвотные препараты . Это плотно васкуляризированная структура с субрегиональной специализацией капилляров, обеспечивающая высокую проницаемость для сигналов циркулирующей крови, что позволяет ей обнаруживать различные химические переносчики в крови и спинномозговой жидкости . [4] [6] Капиллярный кровоток, по-видимому, исключительно медленный в области постремы, что удлиняет время контакта гормонов, переносимых кровью, для взаимодействия с нейрональными рецепторами, участвующими в регуляции артериального давления, жидкостей организма и рвотных реакций. [4] [8]
Автономная регуляция
[ редактировать ]Фенестрированные синусоидальные капилляры постремной области и специализированная область NTS делают эту конкретную область продолговатого мозга важной в автономном контроле различных физиологических систем, включая сердечно-сосудистую систему и системы, контролирующие питание и обмен веществ. [2] [6] Ангиотензин II вызывает дозозависимое повышение артериального давления, не вызывая значительных изменений частоты сердечных сокращений , эффект, опосредованный постремной областью. [11]
Клиническое значение
[ редактировать ]Повреждать
[ редактировать ]Повреждение постремной области, вызванное в первую очередь повреждением или абляцией , препятствует нормальному функционированию постремной области. Эта абляция обычно проводится хирургическим путем с целью определения точного влияния постремной области на остальную часть тела. Поскольку область пострема действует как точка входа в мозг для информации от сенсорных нейронов желудка, кишечника, печени, почек, сердца и других внутренних органов, различные физиологические рефлексы полагаются на постремную область для передачи информации. Область пострема позволяет непосредственно контролировать химический статус организма. Поражения постремной области иногда называют «центральной ваготомией», поскольку они лишают мозг возможности контролировать физиологическое состояние организма через блуждающий нерв. [12] Таким образом, эти поражения служат для предотвращения обнаружения ядов и, следовательно, для предотвращения срабатывания естественной защиты организма. В одном из примеров эксперименты, проведенные Бернштейном и соавт. на крысах показало, что поражения области постремы препятствуют обнаружению хлорида лития , который может стать токсичным при высоких концентрациях. Поскольку крысы не могли обнаружить это химическое вещество, они не могли использовать психологическую процедуру, известную как кондиционирование отвращения к вкусу , заставляющую крысу постоянно поглощать раствор сахарина с литием. Эти данные показывают, что крысы с поражениями области постремы не приобретают нормальных условных вкусовых отвращений, когда в качестве безусловного раздражителя используется хлорид лития. Помимо простых вкусовых отвращений, у крыс с поражением области постремы отсутствовали другие поведенческие и физиологические реакции, связанные с введением токсина и присутствующие в контрольной группе, такие как лежание на животе, задержка опорожнения желудка и гипотермия. [13] Такие эксперименты подчеркивают значение постремной области не только для выявления токсичных веществ в организме, но и для многих физических реакций на токсин.
Эффект дофамина
[ редактировать ]Постремная область также играет важную роль в обсуждении болезни Паркинсона . Препараты, лечащие болезнь Паркинсона с помощью дофамина, оказывают сильное воздействие на постремную область. Эти препараты стимулируют передачу дофамина и пытаются нормализовать двигательные функции, пораженные болезнью Паркинсона. Это работает, поскольку нервные клетки, в частности, в базальных ганглиях , которые играют решающую роль в регуляции движения и являются основным местом патологии Паркинсона, используют дофамин в качестве своего нейромедиатора и активируются лекарствами, повышающими концентрацию дофамина или работать над стимуляцией дофаминовых рецепторов. Дофамину также удается стимулировать область пострема, поскольку эта часть мозга содержит высокую плотность дофаминовых рецепторов. Область пострема очень чувствительна к изменениям токсичности крови и ощущает наличие в крови ядовитых или опасных веществ. В качестве защитного механизма постремная область вызывает рвоту, чтобы предотвратить дальнейшую интоксикацию. Высокая плотность дофаминовых рецепторов в постремной области делает ее очень чувствительной к препаратам, усиливающим дофамин. Стимуляция дофаминовых рецепторов в области пострема активирует эти рвотные центры мозга; вот почему тошнота является одним из наиболее распространенных побочных эффектов противопаркинсонические препараты . [14]
История
[ редактировать ]Область пострема была впервые названа и расположена в общей анатомии мозга Магнусом Густавом Ретциусом , шведским анатомом, антропологом и профессором гистологии . В 1896 году он опубликовал двухтомную монографию по общей анатомии человеческого мозга, в которой упоминается постремная область. [ нужна ссылка ] В 1975 году были опубликованы данные о наличии нейронов в области пострема у нескольких видов млекопитающих. [15]
Ученые стали все больше интересоваться исследованием рвоты в 1950-х годах, возможно, отчасти из-за повышенной осведомленности общества о лучевой болезни - состоянии, при котором многие пациенты, у которых была рвота после радиационного воздействия, умирали. Исследования показали наличие в мозге двух областей, связанных с рвотой: одна — хемосенсор рвоты без координирующей функции, расположенная в четвертом желудочке, и вторая — координатор рвоты без хемосенсорной функции, расположенная в латеральной ретикулярной формации продолговатый мозг. [ нужна ссылка ]
В 1953 году Борисон и Ван определили, что область хемосенсора действует как триггерная зона рвоты в стволе мозга, которую они назвали триггерной зоной хеморецептора (CTZ) рвоты. Используя кошек и собак в качестве модельных организмов, они обнаружили, что удаление этой триггерной зоны из мозга позволяет предотвратить рвоту у животных непосредственно после инъекции определенных химических веществ в кровь, демонстрируя существование взаимосвязи между триггерной зоной и акт рвоты. КТЗ анатомически располагалась в области пострема продолговатого мозга. Постремная область была анатомически идентифицирована и названа почти 60 лет назад, но ее функция оставалась неизвестной до тех пор, пока позже не была подтверждена ее роль в рвоте. [16]
Текущие исследования
[ редактировать ]Сегодня во всем мире продолжаются исследования функций области пострема. Помимо своей роли в рвоте, которую интенсивно изучали исследователи в середине 1900-х годов, активность постремной области была тесно связана с другими вегетативными функциями, такими как регуляция приема пищи, гомеостаз жидкости в организме и сердечно-сосудистая регуляция, посредством поведенческих исследований и электрофизиологические исследования. В 2007 г. в Японии были проведены исследования механизма возбудимости нейронов области постремы внеклеточной АТФ. с фиксированием напряжения На срезах мозга крыс использовались методы записи целых клеток . Результаты показали, что большинство ответов на АТФ являются возбуждающими и опосредуются определенными пуринорецепторами P2, обнаруженными в постремной области. [17] Роль области пострема в отвращении и предпочтении, обусловленном вкусом, изучалась в 2001 году исследователями из Бруклинского колледжа Городского университета Нью-Йорка . В эксперименте проверялось влияние поражений области постремы у крыс на их способность обучаться обусловленному вкусом отвращению к ароматам в сочетании с лечением токсичными лекарствами, что действительно показало, что поражение области постремы приводит к нарушению обучения отвращению к вкусу. [18] Исследование 2009 года следило за развитием постремной области с использованием модели макаки в попытке идентифицировать и охарактеризовать нейротрансмиссию в этой области, а также разрешить существенные несоответствия в ходе исследований. В заключение эти ученые обнаружили, что предыдущие исследования предполагают, что норадреналин и/или дофамин вызывают флуоресценцию СА в области пострема макака-СА, что означает катехоламинергический или производный от амина и функционирующий как нейротрансмиттер или гормон, или и то, и другое. Исследование, однако, обнаружило доказательства секреции нейромедиаторов, а не их высвобождения в везикулах. Кроме того, их результаты пришли к выводу, что ГАМК основным нейромедиатором в постремной области является , а не глутамат . Продолжающиеся исследования продолжают раскрывать расхождения между различными моделями исследований на крысах, кошках, а теперь и на макаках. [19]
Возможные методы лечения
[ редактировать ]В ходе исследования 2002 года в Японии был протестирован препарат, который может быть полезен для сдерживания рвотной реакции на препараты, повышающие концентрацию дофамина. В ходе исследования изучалась рвота, вызванная морфином, у хорьков, и объяснялось, что воздействие морфина вызывало высвобождение дофамина в продолговатом мозге и в постремной области путем активации опиатных рецепторов, что, в свою очередь, вызывало рвоту у хорьков. Тем не менее, предварительное лечение 6-гидроксидофамином, дофаминергическим нейротоксином , значительно снизило количество эпизодов рвоты у хорьков после воздействия морфина. Этот нейротоксин снижает уровень дофамина, норадреналина и гомованилиновой кислоты, метаболита дофамина, и, как известно, разрушает норадренергические и дофаминергические нейроны. В данном случае 6-гидроксидофамин вводили непосредственно в продолговатый мозг, а не в другие части мозга. Это исследование показывает, как можно манипулировать дофаминергическими путями в продолговатом мозге, чтобы уменьшить тошнотворные побочные эффекты, связанные с приемом многих препаратов, повышающих уровень дофамина. [20]
Продолжающиеся патологические исследования
[ редактировать ]Область пострема также показана при лечении инсулином диабета 1 и 2 типа. Особый механизм, используемый препаратом прамлинтидом , действует главным образом на постремную область и приводит к снижению секреции глюкагона , что, в свою очередь, замедляет опорожнение желудка и эффект насыщения. Такое воздействие на постремную область позволяет улучшить гликемический контроль, не вызывая увеличения веса. Поскольку препарат действует на постремную область, дозу следует титровать медленно, чтобы не вызвать у пациента тошноту. [21]
В настоящее время также продолжаются исследования по определению влияния абляции постремной области на гипертонию и сердечно-сосудистую функцию. Например, исследования на крысах и кроликах показывают, что ангиотензин II -зависимая гипертензия устраняется за счет поражения постремной области. [22] [23] Механизм этой физиологической реакции до сих пор не до конца понятен, но способность постремной области регулировать сердечно-сосудистую функцию представляет собой очень интересное направление для нейроэндокринологии.
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и Мирза М., Дас Дж.М. (8 июля 2019 г.). Нейроанатомия, область пострема . СтатПерлс; Остров сокровищ (Флорида): StatPearls Publishing. ПМИД 31334969 . Проверено 4 декабря 2019 г.
- ^ Jump up to: а б с д и ж г Гросс, П.М.; Уолл, К.М.; Панг, Джей Джей; Шейвер, С.В.; Уэйнман, Д.С. (1990). «Микрососудистые специализации, способствующие быстрой интерстициальной дисперсии растворенных веществ в одиночном ядре». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 259 (6, часть 2): R1131–8. дои : 10.1152/ajpregu.1990.259.6.R1131 . ПМИД 2260724 .
- ^ Jump up to: а б Прайс, СиДжей; Хойда, Т.Д.; Фергюсон, А.В. (2007). «Область пострема: монитор мозга и интегратор системного вегетативного состояния». Нейробиолог . 14 (2): 182–194. дои : 10.1177/1073858407311100 . ПМИД 18079557 . S2CID 42334122 .
- ^ Jump up to: а б с Гросс, П.М. (1991). «Морфология и физиология капиллярных систем субобластей подскладчатого органа и постремной области». Канадский журнал физиологии и фармакологии . 69 (7): 1010–25. дои : 10.1139/y91-152 . ПМИД 1954559 .
- ^ Jump up to: а б с д МакКинли, Майкл Дж.; Дентон, Дерек А.; Райан, Филип Дж.; Яо, Сун Т.; Стефанидис, Анета; Олдфилд, Брайан Дж. (14 марта 2019 г.). «От сенсорных околожелудочковых органов к коре головного мозга: нервные пути, контролирующие жажду и голод». Журнал нейроэндокринологии . 31 (3): e12689. дои : 10.1111/jne.12689 . hdl : 11343/285537 . ISSN 0953-8194 . ПМИД 30672620 . S2CID 58947441 .
- ^ Jump up to: а б с д и Шейвер, Стивен В.; Панг, Джуди Дж.; Уолл, Кэтрин М.; Спозито, Надин М.; Гросс, Пол М. (1 марта 1991 г.). «Субрегиональная топография капилляров дорсального блуждающего комплекса крыс: I. Морфометрические свойства». Журнал сравнительной неврологии . 306 (1): 73–82. дои : 10.1002/cne.903060106 . ISSN 0021-9967 . ПМИД 2040730 . S2CID 25603661 .
- ^ Уильямс, председатель редколлегии Питер Л., изд. (1995). Анатомия Грея: анатомические основы медицины и хирургии (38-е изд.). Нью-Йорк: Черчилль Ливингстон . ISBN 978-0-443-04560-8 .
- ^ Jump up to: а б Гросс, Пол М.; Уолл, Кэтрин М.; Уэйнман, Дэн С.; Шейвер, Стивен В. (1 марта 1991 г.). «Субрегиональная топография капилляров дорсального вагусного комплекса крыс: II. Физиологические свойства». Журнал сравнительной неврологии . 306 (1): 83–94. дои : 10.1002/cne.903060107 . ISSN 0021-9967 . ПМИД 2040731 . S2CID 21792860 .
- ^ Миллер, AD; Лесли, РА (1994). «Область Пострема и Рвоты». Границы нейроэндокринологии . 15 (4): 301–320. дои : 10.1006/frne.1994.1012 . ПМИД 7895890 . S2CID 28944289 .
- ^ Ганонг, ВФ (2000). «Циркумвентрикулярные органы: определение и роль в регуляции эндокринной и вегетативной функции». Клиническая и экспериментальная фармакология и физиология . 27 (5–6): 422–427. дои : 10.1046/j.1440-1681.2000.03259.x . ПМИД 10831247 . S2CID 23652492 .
- ^ Велькович, С.; Йованович-Мичич, Д.; Япундзич, Н.; Самарджич, Р.; Белеслин, Д.Б. (1989). «Область пострема и гипертензивный эффект ангиотензина». Метаболические заболевания головного мозга . 4 (1): 61–65. дои : 10.1007/BF00999495 . ПМИД 2704347 . S2CID 12032912 .
- ^ Словарь биологической психологии Филипа Винна (2001) Routledge
- ^ Бернштейн, Иллинойс; Чавес, М.; Аллен, Д.; Тейлор, Э.М. (1992). «Area postrema опосредование физиологических и поведенческих эффектов хлорида лития у крыс». Исследования мозга . 575 (1): 132–137. дои : 10.1016/0006-8993(92)90432-9 . ПМИД 1324085 . S2CID 21897766 .
- ^ Дэвид Э. Голан и Армен Х. Ташджян «Принципы фармакологии: патофизиологические основы лекарственной терапии»
- ^ Клара, ПМ; Бриззи, К. (1975). «Ультраструктурная морфология области постремы беличьей обезьяны». Исследования клеток и тканей . 160 (3): 315–26. дои : 10.1007/BF00222042 . ПМИД 807331 . S2CID 950178 .
- ^ Джон Кухарчик; Дэвид Дж. Стюарт; Алан Д. Миллер (1991). Тошнота и рвота: последние исследования и клинические достижения . Бока-Ратон: CRC Press. ISBN 0-8493-6781-6 . OCLC 24066772 .
- ^ Кодама, Н.; Фунахаси, М.; Мито, Ю.; Минаги, С.; Мацуо, Р. (2007). «Пуринэргическая модуляция возбудимости нейронов области постремы в срезах мозга крыс» . Исследования мозга . 1165 : 50–59. дои : 10.1016/j.brainres.2007.06.003 . ПМИД 17658494 . S2CID 6422014 .
- ^ Тузани, К.; Склафани, А. (2002). «Поражения области постремы ухудшают обучение отвращению к вкусовым токсинам, но не ухудшают обучение предпочтениям в отношении вкусовых и питательных веществ». Поведенческая нейронаука . 116 (2): 256–266. дои : 10.1037/0735-7044.116.2.256 . ПМИД 11996311 .
- ^ Пангестинингсих, ТВ; Хендриксон, А.; Сигит, К.; Саджути, Д.; Нурхидаят; Боуден, DM (2009). «Развитие области пострема: иммуногистохимическое исследование на макаках» . Исследования мозга . 1280 : 23–32. дои : 10.1016/j.brainres.2009.05.028 . ПМЦ 8850980 . ПМИД 19460361 .
- ^ Ёсикава, Т.; Ёсида, Н. (2002). «Влияние лечения 6-гидроксидофамином в постремной области на рвоту, вызванную морфином, у хорьков» . Японский журнал фармакологии . 89 (4): 422–425. дои : 10.1254/jjp.89.422 . ПМИД 12233822 .
- ^ Дэй, К. (2005). «Аналог амилина как противодиабетическое средство». Британский журнал диабета и сосудистых заболеваний . 5 (3): 151–154. дои : 10.1177/14746514050050030701 .
- ^ Аверилл, Д.Б.; Мацумура, К.; Гантен, Д.; Феррарио, CM (1996). «Роль области постремы в трансгенной гипертензии». Гипертония . 27 (3 ч. 2): 591–597. дои : 10.1161/01.HYP.27.3.591 . ПМИД 8613209 .
- ^ Бишоп, В.С.; Хэй, М. (1993). «Участие области Postrema в регуляции симпатического оттока в сердечно-сосудистой системе». Границы нейроэндокринологии . 14 (2): 57–75. дои : 10.1006/frne.1993.1003 . ПМИД 8486207 . S2CID 36991707 .