Клеммная колодка
| Клеммная колодка | |
|---|---|
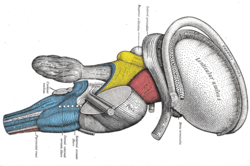 Рассечение ствола головного мозга. Вид сбоку (концевая полоска отмечена вверху справа). | |
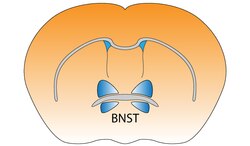 Ядро ложа терминальной полоски мозга мыши | |
| Подробности | |
| Идентификаторы | |
| латинский | клеммная колодка |
| Нейроимена | 286 |
| НейроЛекс ID | бирнлекс_937 |
| ТА98 | A14.1.09.275 |
| ТА2 | 5592 |
| ФМА | 61974 |
| Анатомические термины нейроанатомии | |
( Конечная полоска или терминальная полоска ) — структура головного мозга, состоящая из пучка волокон, идущих вдоль латерального края желудочковой поверхности таламуса . Служа основным выходным путем миндалевидного тела , терминальная полоска проходит от ее центромедиального отдела к вентромедиальному ядру гипоталамуса .
Анатомия
[ редактировать ]Конечная полоска покрывает верхнюю таламостриатную вену , отмечая линию разделения между таламусом и хвостатым ядром , как видно при грубом рассечении желудочков мозга , если смотреть сверху.
Конечная полоска простирается от области межжелудочковых отверстий до височных рогов бокового желудочка , неся волокна от миндалевидного тела к ядрам перегородки , гипоталамическому и таламическому областям мозга. Он также переносит волокна, отходящие от этих областей, обратно к миндалевидному телу.
Ядро ложа терминальной полоски
[ редактировать ]Ядро ложа терминальной полоски (BST) является продолжением ростральной миндалины. [1] Это коррелирует с тревогой в ответ на мониторинг угроз. [2] Считается, что он действует как релейный участок внутри гипоталамо-гипофизарно-надпочечниковой оси и регулирует ее активность в ответ на острый стресс . [3] Однако реакция на стресс зависит от времени, и BNST не активируется из-за контекстуального страха. Это означает, что внезапная пугающая ситуация продолжительностью менее десяти минут не активирует BNST. [4] Также считается, что он способствует поведенческому торможению в ответ на незнакомых людей за счет воздействия орбитофронтальной коры . [5] Было показано, что двустороннее нарушение этого пути ослабляет восстановление поведения, связанного с поиском лекарств, у грызунов. [6]
Известно, что это ядро проецирует тормозные волокна в латеральный гипоталамус и участвует в контроле питания грызунов. Оптогенетическая активация этого ингибирующего пути быстро вызывала ненасытное пищевое поведение у сытых мышей, а оптогенетическое ингибирование этого пути снижает потребление пищи даже у голодающих животных. [7]
Нарушение передачи опиоидных сигналов в BNST, по-видимому, опосредует хронические вызванные алкоголем изменения реакции на стресс.
Половой диморфизм
[ редактировать ]Центральный отдел ядра ложа терминальной полоски (BSTc) имеет половой диморфизм . В среднем BSTc у мужчин в два раза больше, чем у женщин, и содержит в два раза больше соматостатиновых нейронов. [8] прошедших посмертную длительную заместительную гормональную терапию (ЗГТ), Было обнаружено, что в образце шести транс-женщин (от мужчины к женщине), было типичное для женщин количество клеток в BSTc, тогда как у транс-мужчины (от женщины к женщине ) было обнаружено типичное для женщин количество клеток в BSTc. -мужчина) имело типичное для мужчин число. [9] [10] Авторы (Цзян-Нин Чжоу, Фрэнк П.М. Круйвер, Дик Свааб ) также исследовали субъектов с гормонально-зависимыми расстройствами и не обнаружили никакой закономерности между этими расстройствами и BSTc, в то время как у одного нелеченного транссексуала от мужчины к женщине было типичное для женщины количество клетки. Они пришли к выводу, что BSTc предоставляет доказательства нейробиологической основы гендерной идентичности , и предположили, что она определяется еще до рождения.
заместительная гормональная терапия влияет на размер гипоталамуса, Было показано, что [11] хотя в исследование пытались сделать это, включив контрольную группу нетранссексуальных мужчин и женщин, у которых по ряду медицинских причин произошла реверсия гормонов. [9] Утверждение о нейробиологической основе с рождения позже было поставлено под сомнение, хотя и не опровергнуто, последующим исследованием той же группы, которое обнаружило, что половой диморфизм BSTc не присутствует до взрослой жизни (приблизительно 22 года). хотя транссексуалы сообщают, что осознают свою гендерную идентичность с детства. [12]
Поскольку нейроны, экспрессирующие соматостатин, обычно блокируют дендритные входы к постсинаптическому нейрону, тем самым подавляя сигналы, проходящие через ассоциированные структуры, считается, что более крупное ядро ложа терминальной полоски, обнаруженное у мужчин (включая мужчин-трансгендеров), уменьшает реакцию испуга у мужчин и может быть ответственным за более высокую частоту специфических фобий у женщин и возможным источником стереотипа о том, что женщины боятся мышей. [13]
Активность рецептора окситоцина в BNST важна для социального признания у крыс. Как самцы, так и самки крыс, получившие микроинъекцию антагониста рецептора окситоцина, имели более низкие показатели социального распознавания, чем крысы, получившие инъекцию носителя, а микроинъекции окситоцина в BNST улучшали социальную память у самцов, но не самок крыс. [14]
наблюдалось уменьшение размера ядра ложа терминальной полоски У мужчин-преступников -педофилов , а также уменьшение размеров правой миндалевидного тела, гипоталамуса и аномалий в связанных с ними структурах. Авторы предполагают, что дефицит BNST и медиальной миндалины в детстве может вызывать задержку полового созревания. [15]
Ссылки
[ редактировать ]- ^ «Ядро терминальной полоски lm — Медицинский словарь Медицинской Академии» . www.academie-medecine.fr . Проверено 24 мая 2024 г.
- ^ Сомервилл Л., Уэлен П., Келли В. (2010). «Ядро человеческой кровати терминальной полоски индексирует сверхбдительный мониторинг угроз» . Биологическая психиатрия . 68 (5): 416–424. doi : 10.1016/j.biopsych.2010.04.002 . ПМК 2921460 . ПМИД 20497902 .
- ^ Чой Д., Фюрей А., Эвансон Н., Острандер М., Ульрих-Лай Ю., Герман Дж. (2007). «Ядро ложа субрегионов конечной полоски дифференциально регулирует активность гипоталамо-гипофизарно-надпочечниковой оси: значение для интеграции лимбических входов» . Дж. Нейроски . 27 (8): 2025–34. doi : 10.1523/JNEUROSCI.4301-06.2007 . ПМЦ 6673539 . ПМИД 17314298 .
- ^ Хаммак С.Е., Тодд Т.П., Кочо-Шелленберг М., Бутон М.Э. (2015). «Роль ложного ядра конечной полоски в приобретении контекстуального страха в длинных или коротких интервалах контекстного шока» . Поведенческая нейронаука . 129 (5): 673–678. дои : 10.1037/bne0000088 . ПМЦ 4586907 . ПМИД 26348716 .
- ^ Фокс А., Шелтон С., Оукс Т., Конверс А., Дэвидсон Р., Калин Н. (2010). «Поражения орбитофронтальной коры изменяют активность, связанную с тревогой, в ядре ложа приматов конечной полоски» . Дж. Нейроски . 30 (20): 7023–27. doi : 10.1523/JNEUROSCI.5952-09.2010 . ПМЦ 2915894 . ПМИД 20484644 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сюзанна Эрб · Наталина Сальмасо · Деметра Родарос Джейн Стюарт (2001). «Роль пути, содержащего CRF, от центрального ядра миндалины к ядру ложа конечной полоски в вызванном стрессом возобновлении поиска кокаина у крыс». Психофармакология . 158 (4): 360–65. дои : 10.1007/s002130000642 . ПМИД 11797056 . S2CID 23284158 .
- ^ Дженнингс, Джошуа Х.; Рицци, Джорджио; Стаматакис, Алиса М.; Унг, Рэндалл Л.; Стубер, Гаррет Д. (27 сентября 2013 г.). «Архитектура тормозного контура латерального гипоталамуса управляет питанием» . Наука . 341 (6153): 1517–1521. Бибкод : 2013Sci...341.1517J . дои : 10.1126/science.1241812 . ISSN 0036-8075 . ПМЦ 4131546 . ПМИД 24072922 .
- ^ Свааб Д. (2007). «Половая дифференциация мозга и поведения». Передовая практика Res Clin Endocrinol Metab . 21 (3): 431–44. дои : 10.1016/j.beem.2007.04.003 . ПМИД 17875490 .
- ^ Перейти обратно: а б Чжоу Дж., Хофман М., Гурен Л., Свааб Д. (1995). «Половые различия в человеческом мозге и их связь с транссексуальностью» . Природа . 378 (6552): 68–70. Бибкод : 1995Natur.378...68Z . дои : 10.1038/378068a0 . hdl : 20.500.11755/9da6a0a1-f622-44f3-ac4f-fec297a7c6c2 . ПМИД 7477289 . S2CID 4344570 .
- ^ Круйвер Ф., Чжоу Дж., Пул С., Хофман М., Гурен Л., Свааб Д. (2000). «Транссексуалы, переходящие от мужчины к женщине, имеют женское количество нейронов в лимбическом ядре» . Дж. Клин. Эндокринол. Метаб . 85 (5): 2034–41. дои : 10.1210/jcem.85.5.6564 . ПМИД 10843193 .
- ^ Хулшофф Пол Х.Э., Коэн-Кеттенис П.Т., Ван Харен Н.Е., Пепер Дж.С., Бранс Р.Г., Кан В. и др. (2006). «Смена пола меняет ваш мозг: влияние тестостерона и эстрогена на структуру мозга взрослого человека». Европейский журнал эндокринологии . 155 (Приложение 1): С107–С114. дои : 10.1530/eje.1.02248 .
- ^ Чунг В., Де Врис Г., Свааб Д. (2002). «Половая дифференциация ядра ложа терминальной полоски у человека может распространяться и во взрослом возрасте» . Дж. Нейроски . 22 (3): 1027–33. doi : 10.1523/JNEUROSCI.22-03-01027.2002 . ПМК 6758506 . ПМИД 11826131 .
- ^ Кэмерон, Аласдер (2004). Ускоренный курс психиатрии . ООО "Эльзевир" ISBN 978-0-7234-3340-8 .
- ^ Дюмэ, К.М.; Алонсо, АГ; Иммормино, Массачусетс; Бредеволд, Р; Винема, штат Ага (февраль 2016 г.). «Участие системы окситоцина в ядре ложа терминальной полоски в половой регуляции социального признания» . Психонейроэндокринология . 64 : 79–88. дои : 10.1016/j.psyneuen.2015.11.007 . ПМЦ 4698213 . ПМИД 26630388 .
- ^ Шильц К., Витцель Дж., Нортофф Г., Зирхут К., Губка У., Феллман Х., Кауфманн Дж., Темпельманн С., Вибкинг С., Богертс Б. (2007). «Патология головного мозга у преступников-педофилов: свидетельства уменьшения объема правой миндалины и связанных с ней диэнцефальных структур». Архив общей психиатрии . 64 (6): 737–746. дои : 10.1001/archpsyc.64.6.737 . ПМИД 17548755 .