Барьер
| Барьер | |
|---|---|
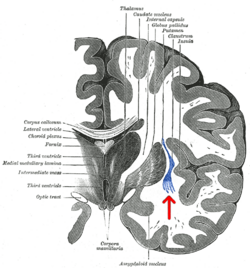 Корональный срез человека головного мозга . Клауструм указан стрелкой. | |
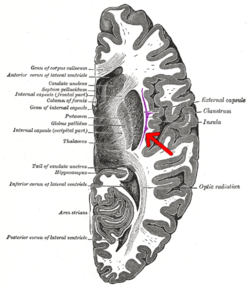 Поперечный срез головного мозга человека. Клауструм указан стрелкой. | |
| Подробности | |
| Часть | Мозг |
| Артерия | Средняя мозговая артерия |
| Идентификаторы | |
| МеШ | D000079482 |
| Нейроимена | 252 |
| НейроЛекс ID | бирнлекс_1522 |
| ТА98 | A14.1.09.421 |
| ТА2 | 5535 |
| ФМА | 67440 |
| Анатомические термины нейроанатомии | |
Клауструм ( лат . «закрывать» или «закрывать») представляет собой тонкий слой нейронов и поддерживающих глиальных клеток , который соединяется с корой головного мозга и подкорковыми областями, включая миндалевидное тело , гиппокамп и таламус головного мозга . [1] [2] Он расположен между островковой корой латерально и скорлупой медиально и окружен крайней и наружной капсулами соответственно. [3] [1] [4] Кровь к Клауструму снабжает средняя мозговая артерия . [1] Считается, что это наиболее плотно связанная структура в мозге, и поэтому предполагается, что она позволяет интегрировать различные корковые сигналы, такие как зрение, звук и осязание, в один опыт . [4] [5] Другие гипотезы предполагают, что клауструм играет роль в обработке значимости , направляя внимание на наиболее поведенчески значимые стимулы среди фонового шума. [6] Клауструм трудно изучать, учитывая ограниченное число людей с клаустральными поражениями и плохое разрешение нейровизуализации .
Клауструм состоит из различных типов клеток, различающихся по размеру, форме и нейрохимическому составу. Существует пять типов клеток, и большинство из этих клеток напоминают пирамидальные нейроны коры головного мозга. [7] [8] Внутри клауструма нет ламинарной организации типов клеток, как в корковых слоях , а тела клеток могут быть пирамидальными, веретенообразными или круглыми. [1] Основным типом клеток, обнаруженным в клауструме, является нейрон Гольджи типа I , который представляет собой большую клетку с дендритами , покрытыми шипами . [9] [8]
Считается, что благодаря межполушарным связям клауструм играет роль в синхронизации активности в широко разделенных, но функционально связанных частях мозга (например, лобных полях глаз и зрительной коре ). [1] [10] [11] Таким образом, считается, что клауструм играет роль в объединении различных информационных модальностей, потенциально поддерживающих само сознание. [10] [12] Другая предлагаемая функция клауструма — различать релевантную и нерелевантную информацию, чтобы последнюю можно было игнорировать. [5] [12] [13] Корковые компоненты сознания включают лобно-теменную кору, поясную извилину и предклинье . Из-за широкой связи клауструма с этими областями предполагается, что он может играть роль как во внимании, так и в сознании . Нейронные сети, которые обеспечивают устойчивое внимание и сознание, посылают сигналы в клауструм, и один отчет о случае у людей предполагает, что электрическая стимуляция вблизи клауструма обратимо нарушила состояние сознания пациента. [14]
Структура
[ редактировать ]Клауструм представляет собой небольшую двустороннюю структуру серого вещества (составляющую примерно 0,25% коры головного мозга), расположенную глубоко в островковой коре и крайней капсуле и поверхностно по отношению к наружной капсуле и базальным ганглиям. [1] [15]
Как уже упоминалось, его название означает «скрытый или запертый» и впервые было обнаружено в 1672 году, а более подробные описания появились позже, в 19 веке. [1] [15] Несмотря на то, что региональные нейроанатомические границы клауструма были определены, в литературе по-прежнему отсутствует консенсус относительно определения его точных границ. [13] [16] [17] хотя встреча экспертов в 2019 году определила рамки для обозначения структур разных видов. [3]
Соединения
[ редактировать ]В первых сводках отчетов 20-го века особое внимание уделялось корковым входам и выходам. [18] Однако более поздние исследования показали, что клауструм имеет обширные связи с корковыми и подкорковыми областями. [19] В частности, электрофизиологические исследования показывают обширные связи с ядрами таламуса и базальными ганглиями , в то время как изотопологические отчеты связывают Клауструм с префронтальной, лобной, теменной, височной и затылочной корой. [20] [21] Дополнительные исследования также изучали связь клауструма с хорошо описанными подкорковыми трактами белого вещества. Такие структуры, как лучистая венечка , затылочно-лобный пучок и крючковидный пучок, выступают на Клауструм из лобной, перицентральной, теменной и затылочной областей. [22] Реципрокные связи существуют также с двигательными, соматосенсорными , слуховыми и зрительными областями коры. [13] В целом, эти результаты делают клауструм наиболее тесно связанной структурой в региональном объеме мозга и предполагают, что она может служить центром координации активности мозговых цепей. [23] [24] Даже при такой обширной связи большинство проекций к клауструму и от него являются ипсилатеральными (хотя все еще существуют контралатеральные проекции), и существует мало данных для описания его афферентных или эфферентных связей со стволом головного мозга и спинным мозгом. [13] [18] [25] Таким образом, корковая и подкорковая связь клаустра подразумевает, что она больше всего участвует в обработке сенсорной информации, а также в физическом и эмоциональном состоянии животного.
Микроанатомия
[ редактировать ]Входные данные в клауструм организованы по модальности, которые включают префронтальную, зрительную, слуховую и соматомоторную области обработки. Точно так же, как морфология нейронов в пластинках Рексада спинного мозга указывает на функцию, зрительные, слуховые и соматомоторные области внутри клауструма имеют сходные нейроны со специфическими функциональными характеристиками. Например, часть клауструма, которая обрабатывает зрительную информацию ( в первую очередь синтезируя афферентные волокна, связанные с нашим периферическим зрительным полем ), состоит из большинства бинокулярных клеток, которые имеют «удлиненные рецептивные поля и не имеют ориентационной избирательности». [26] [27] Этот фокус на периферической сенсорной системе не является изолированным явлением, поскольку большинство сенсорных афферентов, попадающих в клауструм, несут периферическую сенсорную информацию. Более того, клауструм обладает четкой топологической организацией для каждой сенсорной модальности, а также плотными связями, которые он разделяет с лобной корой. [28] [29] Например, существует ретинотопическая организация в области зрительной обработки клауструма, которая отражает организацию зрительных ассоциативных кортикальных слоев и V1, сходным (но менее сложным) способом с ретинотопической консервацией в латеральном коленчатом ядре. [13]
Внутри клауструма в локальной связности преобладает прямое дисинаптическое торможение, при котором интернейроны, экспрессирующие парвальбумин, подавляют активность близлежащих проекционных нейронов. [30] Сами локальные интернейроны связаны как через химические, так и через электрические синапсы , что обеспечивает широкомасштабное и синхронное торможение локальных цепей клауструма. В недавних исследованиях клауструма на мышах [28] и летучие мыши , [31] Было обнаружено, что кортикально выступающие нейроны возбуждающего клаустра образуют синапсы поперек переднезадней оси и смещаются в сторону нейронов, которые не имеют общих проекционных целей, с возможной функцией объединения активности различных афферентных модулей. [29] В совокупности эти две схемы позволяют предположить, что клауструм способен выполнять локальные преобразования разнообразной входной информации, поступающей со всего мозга.
Типы ячеек
[ редактировать ]Клауструм состоит из различных типов клеток, которые различаются по размеру, форме и нейрохимическому составу. [4] Типы возбуждающих клеток в клауструме состоят из двух основных классов, которые по-разному проецируются в корковые и подкорковые области мозга. [ нужна ссылка ] . Тормозные нейроны составляют лишь 10–15% нейронов в клауструме и состоят из трех типов, экспрессирующих парвальбумин , соматостатин или вазоактивный кишечный пептид , подобно тормозным нейронам в коре головного мозга. [32] Наконец, многие исследования показывают, что клауструм лучше всего отличается структурно благодаря выступающему сплетению парвальбумин-положительных волокон, образованных типами ингибирующих экспрессию парвальбумина клеток. [5] использование основного белка миелина (ОБМ) и ретроградно перемещающегося холерного токсина . В недавних исследованиях в качестве эффективных методов идентификации клауструма дополнительно использовалось [29] [33]
несколько подходов , включая электрофизиологические, морфологические, генетические и коннектомные подходы. Для оценки типов клеток клауструма на мышах было использовано [28] [29] [34] [35] [36] Хотя до сих пор не достигнуто четкого консенсуса относительно точного количества типов возбуждающих клеток, недавние исследования показали, что кортикально- и субкортикально выступающие нейроны клауструма, вероятно, различаются и различаются по нескольким показателям, таким как их внутренние электрофизиологические профили, афферентные проекции и нейромодулирующий профиль. [28] [34] [37]
Функция
[ редактировать ]Было показано, что клауструм оказывает широкое воздействие на многочисленные корковые компоненты, каждый из которых связан с наличием компонентов сознания и устойчивого внимания. Это связано с широким распространением связей с лобно-теменными областями, поясной извилиной коры и таламусами. Устойчивое внимание обеспечивается связями с поясной корой, височной корой и таламусом.
Крик и Кох предполагают, что клауструм выполняет роль, аналогичную роли дирижера в оркестре, поскольку он пытается координировать функции всех связей. [1] Эту аналогию с «проводником» можно также поддержать через связи между клаустральной, сенсорной и лобной областями. Было подтверждено, что клауструм взаимно связан с префронтальной корой, зрительной, слуховой, сенсорной и моторной областями соответственно. Связь с этими модальностями дает представление о функциональности клауструма. Здесь предполагается, что клауструм выполняет функцию шлюза избирательного внимания. Посредством этого процесса шлюзования клауструм может выборочно контролировать входные данные от этих модальностей, чтобы облегчить процесс «фокусировки». Также было высказано предположение, что он действует в противоположном контексте; что посредством вызывающей разногласия нормализации клауструм может реализовывать сопротивление определенным модальностям ввода, чтобы предотвратить «отвлечение».
Потенциальная функция
[ редактировать ]Клауструм, чтобы облегчить сознание, должен будет интегрировать различные сенсорные и моторные модальности из разных частей коры. Анатомические связи клауструма наблюдались с помощью диффузионно-тензорной визуализации (DTI). Функциональная магнитно-резонансная томография (фМРТ) изучает уровень насыщения крови кислородом в мозге как способ наблюдения за активностью определенных областей коры головного мозга. Сканирование фМРТ демонстрирует снижение активности у крыс под наркозом по сравнению с бодрствованием, в частности, соединения клауструма с медиальной префронтальной корой (мПФК) и медиодорсальным таламусом (MD таламус). Клауструм связан с клауструмом контралатерального полушария прочными функциональными связями. Также существуют связи с таламусом MD, mPFC, а также окружающими и отдаленными областями коры. [10]
Электрическая стимуляция спинного щита кошек вызывает возбуждающие реакции в зрительной коре. Клауструм анатомически расположен в месте слияния большого количества трактов белого вещества, соединяющих различные части коры. Это также предполагает роль интеграционного центра для этих различных модальностей, таких как сенсорная и моторная. Было показано, что между ашиповыми интернейронами клауструма существуют щелевые соединения, что указывает на роль в его способности синхронизировать эти модальности при получении входных данных. [1]
Дополнительные исследования указывают на участие в пространственной навигации. [38] и медленноволновой сон . [39] [40]
Внимание
[ редактировать ]Клауструм обладает дифференциальной способностью выбирать между информацией, относящейся к задаче, и информацией, не относящейся к задаче, для обеспечения направленного внимания. Он содержит самую высокую плотность соединительных участков белого вещества коры. Это подтверждает идею взаимодействия и координации между различными областями мозга. [12] Клауструм имеет региональную специфику; Информация, поступающая из зрительных центров, проецируется на определенные участки нейронов серого вещества в структурах и слуховой коре. [1] Неожиданные стимулы также активируют клауструм, вызывая немедленную фокусировку или распределение функций. У низших млекопитающих (например, крыс) клаустральные области получают информацию от соматосенсорных модальностей, таких как двигательный контроль усов, из-за его сенсорного и дискриминационного использования у этих млекопитающих. [13]
Функционально предлагается разделить внимание между этими модальностями. Само внимание рассматривалось как обработка сверху вниз или обработка снизу вверх; оба контекстуально соответствуют тому, что наблюдается в клауструме структурно и функционально, подтверждая представление о том, что взаимодействия происходят с сенсорными областями высокого порядка, участвующими в кодировании объектов и функций. Например, входные данные из префронтальной коры будут определять внимание на основе поведения, ориентированного на более высокие когнитивные задачи. Более того, было показано, что индукция электрической стимуляции клауструма вызывает заторможенность чтения, пустой взгляд и отсутствие реакции. Сообщалось, что клауструм имеет базальную частоту, которая модулируется для увеличения или уменьшения в зависимости от направленного внимания. Например, проекции на двигательные и глазодвигательные области помогут с помощью движения взгляда направить внимание на новые стимулы за счет увеличения частоты срабатывания клаустральных нейронов. [13]
Сальвинорин А, активное галлюциногенное соединение, обнаруженное в Salvia Divinorum, способно вызывать потерю сознания. Потребление сальвинорина А может вызвать синестезию, при которой разные сенсорные модальности интерпретируются разными сенсорными кортикальными слоями. (Например: видеть звуки, ощущать вкус цветов.) Это подтверждает идею внутриталамической сегрегации и проводимости (внимания). Клауструм имеет каппа-опиоидные рецепторы, с которыми связывается сальвинорин А, вызывая этот эффект. [4] [13]
Эмпирические данные
[ редактировать ]Высокочастотная стимуляция (ВЧС) в клауструме кошек способна вызывать вегетативные изменения и вызывать «синдром инактивации». Этот синдром описывается как снижение осознанности, что указывает на связь между клауструмом и сознанием. [41] У людей можно наблюдать тот же эффект. Стимуляция левой Клауструма у людей вызывала «полную остановку волевого поведения, отсутствие реакции и амнезию без негативных двигательных симптомов или простой афазии», что указывает на вовлечение сознания. [14] Кроме того, исследования МРТ показали, что повышенная интенсивность сигнала в клауструме связана с эпилептическим статусом – состоянием, при котором эпилептические припадки следуют друг за другом без восстановления сознания между событиями. [42] [43] Кроме того, повышенная интенсивность сигнала связана с фокальными дискогнитивными припадками, которые представляют собой припадки, вызывающие нарушение осознанности или сознания без судорог. Человек перестает осознавать свое окружение, и припадок проявляется в виде пустого или пустого взгляда на какое-то окно времени.
Использование оперантной задачи кондиционирования в сочетании с HFS клауструма привело к значительным изменениям в поведении крыс; это включало модулированные двигательные реакции, бездействие и снижение реактивности. (добавить ссылку Koubessi) Помимо этого, исследования также показали, что клауструм активен во время быстрого сна, наряду с другими структурами, такими как зубчатая извилина. Они играют ассоциативную роль в пространственной памяти, что позволяет предположить, что в этих областях имеет место некоторая форма консолидации памяти. [5]
Поражения и сознание
[ редактировать ]Функционально клауструм интегрирует различные корковые входы через свои связи в сознание. На основании его структуры и связности предполагается, что его функция связана с координацией различных функций мозга; т.е. аналогия с проводником. Сознание функционально можно разделить на два компонента: (i) бодрствование, которое представляет собой возбуждение и бдительность; (ii) содержание сознания, которое представляет собой обработку содержания. Для лучшего понимания функциональной роли клауструма было предпринято исследование черепно-мозговых травм у ветеранов войны. Повреждение клауструма было связано с длительностью потери сознания, а не с ее частотой. Размер поражения коррелировал с большей продолжительностью событий LOC. Никаких последствий для ослабления когнитивной обработки обнаружено не было. [4]
В одном тематическом исследовании было показано, что сознание нарушалось при стимуляции крайней капсулы мозга, которая находится в непосредственной близости от клауструма, так что после прекращения стимуляции сознание восстанавливалось. [14] Другое исследование, посвященное симптоматологии шизофрении, установило, что тяжесть бреда связана с уменьшением объема серого вещества левой клаустры; постулируя, что существуют корреляции между структурой и положительными симптомами, наблюдаемыми при этом психическом расстройстве. Дополнительным подтверждением этой корреляции между шизофренией и клауструмом является увеличение объема белого вещества, попадающего в клауструм. [44] Подтверждена обратная корреляция между объемом серого вещества и выраженностью галлюцинаций в контексте слуховых галлюцинаций при шизофрении. [45] Кроме того, чтобы увидеть полную потерю функции клауструма, должно произойти поражение обеих клаустр в каждом полушарии. [1]
Однако исследование 2019 года, состоящее из электрической стимуляции клауструма, не выявило нарушений сознания ни у одного из пяти пациентов, подвергшихся анализу. Испытуемые пациенты сообщали о субъективных ощущениях в различных сенсорных областях и демонстрировали рефлекторные движения, но ни у одного из них не наблюдалось потери сознания, что ставит под сомнение способность клауструма нарушать сознание при электрической стимуляции. [46]
Клиническое значение
[ редактировать ]Шизофрения
[ редактировать ]Повреждения Клауструма могут имитировать различные распространенные заболевания или психические расстройства; задержка развития этой структуры, по-видимому, связана с аутизмом . Клауструм может быть вовлечен в развитие шизофрении, поскольку результаты показывают усиление положительных симптомов, таких как бред, когда объем серого вещества левой клауструма и правой островковой доли уменьшается. [45]
Эпилепсия
[ редактировать ]Также считается, что Клауструм играет роль в эпилепсии; МРТ выявила повышенную интенсивность клаустрального сигнала у людей, у которых диагностирована эпилепсия. В некоторых случаях судороги, как правило, возникают из клауструма, когда они участвуют в ранних судорогах, вызванных каиновой кислотой .
Сознание
[ редактировать ]Единственное тематическое исследование показало, что сознание нарушалось при электрической стимуляции области между островком и клауструмом; сознание восстанавливалось, когда стимуляция прекращалась. [4] [14] Пациенты с поражением левой клауструма с большей вероятностью испытывали потерю сознания по сравнению с теми, у кого были поражения за пределами клауструма. [4] Например, пациент, подвергшийся электродной стимуляции клауструма, перестал читать, тупо смотрел и не реагировал. Как только электрод был удален, пациент возобновил чтение и не мог вспомнить события, связанные с ошеломлением. [13]
Исследование 2019 года, состоящее из электрической стимуляции клауструма, не выявило нарушений сознания ни у одного из пяти пациентов, подвергшихся анализу. Испытуемые пациенты сообщали о субъективных ощущениях в различных сенсорных областях и демонстрировали рефлекторные движения, но ни у одного из них не наблюдалось потери сознания, что ставит под сомнение способность клауструма нарушать сознание при электрической стимуляции. [46]
Исследование 2020 года, включавшее искусственную активацию клаустра с помощью оптогенетической световой стимуляции, заставило замолчать активность мозга в коре головного мозга - феномен, известный как «состояние сна», которое можно наблюдать, когда мыши спят или отдыхают бодрствуя (тихое бодрствование). [40] Авторы заявляют, что «клауструм является координатором глобальной медленноволновой активности, и это настолько захватывающе, что мы приближаемся к связыванию конкретных связей и действий мозга с окончательной загадкой сознания».
Однако учёный-когнитивист Стеван Харнад не считает клауструм «переключателем» сознания, а просто переключателем бодрствования. Он утверждает, что если бы клауструм действительно был переключателем сознания, искусственная активация клауструма не привела бы к потере сознания пациента, но вместо этого пациент продолжал бы вести себя нормально, а позже сообщал, что не испытывал никаких ощущений в течение определенного периода времени. когда их клауструм стимулировался. [47]
Псилоцибин
[ редактировать ]Клауструм имеет высокую плотность рецепторов 5HT2A , что означает, что на него существенно влияют серотонинергические психоделики, такие как псилоцибин . Псилоцибин, по-видимому, влияет на функциональную связь клауструма с сетью режима по умолчанию (DMN) и лобно-теменной сетью управления задачами (FPTC). Было обнаружено, что псилоцибин значительно снижает функциональную связь правой клаустры с сетью режима по умолчанию и увеличивает связь правой клаустры с сетью управления лобно-теменной задачей. [48]
паркинсонизм
[ редактировать ]Группа исследователей под руководством нейробиологов из Медицинского центра Бет Исраэль Диаконесса определила поражения в клауструме как вероятную причину паркинсонизма при различных состояниях. Команда использовала новую методологию, называемую картированием сети поражений, чтобы обнаружить причины паркинсонизма у 29 пациентов, чьи симптомы не были результатом болезни Паркинсона, а скорее были связаны с поражением головного мозга – аномалией или повреждением мозга, видимыми при визуализации мозга. Картирование 29 поражений, расположенных в разных областях мозга, показало, что связь с клауструмом является единственным наиболее чувствительным и специфичным маркером паркинсонизма, вызванного поражением. [49]
Тревога и стресс
[ редактировать ]У мышей подавление клауструма, по-видимому, ослабляет тревогу / стресс и повышает хроническую стрессоустойчивость. [50]
Другие животные
[ редактировать ]У животных с помощью отслеживания путей результаты показали, что клауструм имеет обширные связи по всей коре с сенсорными и моторными областями, а также с гиппокампом. Были использованы различные модели животных, такие как кошки, грызуны и обезьяны.

Кошки
[ редактировать ]Этот раздел может быть слишком техническим для понимания большинства читателей . ( Апрель 2019 г. ) |
У кошек высокочастотная стимуляция (HFS) клаустры может изменить двигательную активность, вызвать вегетативные изменения и спровоцировать «синдром инактивации», описываемый как «снижение осознанности». Записи, в основном у кошек и приматов, показывают, что клаустральные нейроны реагируют на сенсорных раздражителей и во время произвольных движений. [5] Картирование от зрительной коры до клауструма включает всего одну карту, которая включает V1 и три другие зрительные области. Клетки V1 являются частью слоя 6, который отличается от клеток, идущих в латеральное коленчатое ядро ; эти клетки используют глутамат в качестве нейромедиатора . Клауструм кошки имеет 3 определенные зоны: (1) передняя дорсальная зона, которая соединяется с моторной и соматосенсорной корой, (2) задняя дорсальная зона, которая связана со зрительной корой и (3) третья зона, которая находится вентральнее зрительный и соединяется со слуховыми областями. [1]
Сенсорная информация разделяется в зависимости от модальностей, при этом предпочтение отдается периферической сенсорной информации. У кошки входной сигнал поступает из различных зрительных областей коры и проецируется обратно в эту область. Эти петли являются ретинотопическими, то есть области, получающие визуальную информацию, отвечают за ту же область зрительного поля, что и область коры, проецирующуюся на клауструм. Зрительная клауструм представляет собой единую карту контралатерального зрительного полуполя, получающую информацию на основе движения на периферии зрительного поля и не обладающую реальной избирательностью. [51] [52] Что касается соматочувствительности, клауструм кошки получает плотные входные сигналы от первичной соматосенсорной коры (S1), но более слабые входные сигналы от вторичной соматосенсорной коры (S2). Входные сигналы от S1 перекрываются с входными сигналами первичной моторной коры (по крайней мере, входными сигналами от передних лап обоих). [53] Клауструм грызунов не получает входных данных от S1 или S2 и в основном управляется моторной корой.
Грызуны
[ редактировать ]У крыс области двигательных усов получают входные сигналы от ипсилатеральной клауструма, но затем проецируются на контралатеральную клауструм. [5] Сенсорная бочкообразная кора и первичная зрительная кора также получают входные данные от ипсилатеральной клауструма, но отправляют очень мало проекций обратно в клауструм. Таким образом, исследования указывают на четкую структуру связей клауструма с различными областями коры. Это предполагает, что они играют не диффузную роль, а специализированную роль в корковой обработке. [5]
У мышей волокна парвальбумина тесно связаны между собой химическими и электрическими синапсами. Кроме того, они тесно связаны с клаустрокортикальными нейронами, что позволяет предположить, что эти тормозные интернейроны сильно модулируют их активность. [5] Эти локальные сети предполагают синхронизировать активность клаустрокортикальных проекций, чтобы, таким образом, влиять на мозговые ритмы и скоординированную деятельность различных областей коры головного мозга. Существуют дополнительные классы тормозных интернейронов с локальными связями внутри клаустрокортикальных нейронов. [5]
Эксперименты на мышах, отслеживающие активность клаустрокортикальных аксонов в зависимости от изменяющихся зрительных стимулов, позволяют предположить, что клауструм сигнализирует об изменении стимулов. [5] Хотя клаустрокортикальный входной сигнал в зрительные области коры был задействован, самые сильные реакции наблюдались в областях коры более высокого порядка, включая переднюю поясную извилину, которая плотно иннервируется клаустральной проекцией. [5]
Обезьяны
[ редактировать ]У обезьян широко распространены связи Клауструма с аллокортикальными и неокортикальными областями. Эти связи проецируются, среди прочего, на лобную долю, зрительные области коры, височную кору, теменно-затылочную кору и соматосенсорные области. [1] Подкорковыми областями, принимающими проекции, являются миндалевидное тело, хвостатое ядро и гиппокамп. Неизвестно, существуют ли области коры, которые не получают входных данных от клауструма. Кроме того, в мозге обезьяны наблюдаются большие и малые типы аспид, которые классифицируются как «нейроны локальной цепи».
Дорсальная клауструм имеет двунаправленные связи с двигательными структурами коры. [1] Взаимосвязь между движением животного и поведением нейронов дорсокаудальной клауструма следующая: 70% двигательных нейронов неизбирательны и могут активировать любые толчковые, тянущие или поворотные движения в передней конечности, остальные были более проницательными и выполняли только одно из трех движений, перечисленных выше. [1]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот Крик ФК, Кох С (июнь 2005 г.). «Какова функция клауструма?» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 360 (1458): 1271–9. дои : 10.1098/rstb.2005.1661 . ПМК 1569501 . ПМИД 16147522 .
- ^ Смит Дж.Б., Ли А.К., Джексон Дж. (2020). «Клауструм» . Современная биология . 30 (23): Р1401–Р1406. дои : 10.1016/j.cub.2020.09.069 . ПМИД 33290700 . S2CID 227507231 .
- ^ Jump up to: а б Смит Дж.Б., Аллоуэй К.Д., Хоф П.Р., Орман Р., Резер Д.Х., Ватакабе А., Уотсон ГДР (февраль 2019 г.). «Отношения между клауструмом и эндопириформным ядром: взгляд на консенсус по межвидовой гомологии» . Джей Комп Нейрол . 527 (2): 476–499. дои : 10.1002/cne.24537 . ПМК 6421118 . ПМИД 30225888 .
- ^ Jump up to: а б с д и ж г Чау А., Салазар А.М., Крюгер Ф., Кристофори И., Графман Дж. (ноябрь 2015 г.). «Влияние поражения клауструма на сознание человека и восстановление функций». Сознание и познание . 36 : 256–64. дои : 10.1016/j.concog.2015.06.017 . ПМИД 26186439 . S2CID 46139982 .
- ^ Jump up to: а б с д и ж г час я дж к Браун С.П., Матур Б.Н., Олсен С.Р., Луппи П.Х., Бикфорд М.Э., Цитри А. (ноябрь 2017 г.). «Новые прорывы в понимании роли функциональных взаимодействий между неокортексом и клауструмом» . Журнал неврологии . 37 (45): 10877–10881. doi : 10.1523/JNEUROSCI.1837-17.2017 . ПМК 5678020 . ПМИД 29118217 .
- ^ Смит Дж.Б., Уотсон Дж., Лян З., Лю Ю., Чжан Н., Аллоуэй К.Д. (2020). «Роль Клауструма в обработке значимости?» . Границы нейроанатомии . 13 (64): Р1401–Р1406. дои : 10.3389/fnana.2019.00064 . ПМК 6594418 . ПМИД 31275119 .
- ^ Браак Х, Браак Э (1982). «Типы нейронов в Клауструме человека». Анатомия и эмбриология . 163 (4): 447–60. дои : 10.1007/BF00305558 . ПМИД 7091711 . S2CID 7566723 .
- ^ Jump up to: а б Николенко В.Н., Ризаева Н.А., Берака Н.М., Оганесян М.В., Кудряшова В.А., Дубовец А.А., Борминская И.Д., Булыгин К.В., Синельников М.Ю., Алиев Г (июль 2021 г.). «Тайна клаустральных нейронных цепей и последние новости о ее роли в нейродегенеративной патологии» . Поведенческие функции мозга . 17 (1): 8. дои : 10.1186/s12993-021-00181-1 . ПМЦ 8261917 . ПМИД 34233707 .
- ^ Матур Б.Н. (2014). «Клауструм в обзоре» . Передняя система нейронов . 8 : 48. дои : 10.3389/fnsys.2014.00048 . ПМЦ 3983483 . ПМИД 24772070 .
- ^ Jump up to: а б с Смит Дж.Б., Лян З., Уотсон Г.Д., Аллоуэй К.Д., Чжан Н. (июль 2017 г.). «Межполушарная функциональная связность клауструма в состоянии покоя в бодрствующем и наркотическом состояниях» . Структура и функции мозга . 222 (5): 2041–2058. дои : 10.1007/s00429-016-1323-9 . ПМЦ 5382132 . ПМИД 27714529 .
- ^ Стивенс CF (июнь 2005 г.). «Сознание: Крик и клауструм». Природа . 435 (7045): 1040–1. Бибкод : 2005Natur.435.1040S . дои : 10.1038/4351040a . ПМИД 15973394 . S2CID 5402518 .
- ^ Jump up to: а б с Торгерсон CM, Иримия А., Го С.Ю., Ван Хорн JD (март 2015 г.). «Связность DTI клауструма человека» . Картирование человеческого мозга . 36 (3): 827–38. дои : 10.1002/hbm.22667 . ПМК 4324054 . ПМИД 25339630 .
- ^ Jump up to: а б с д и ж г час я Голл Ю., Атлан Дж., Ситри А. (август 2015 г.). «Внимание: клауструм». Тенденции в нейронауках . 38 (8): 486–95. дои : 10.1016/j.tins.2015.05.006 . ПМИД 26116988 . S2CID 38353825 .
- ^ Jump up to: а б с д Кубейси М.З., Бартоломей Ф., Бельтаги А., Пикард Ф. (август 2014 г.). «Электрическая стимуляция небольшого участка мозга обратимо нарушает сознание». Эпилепсия и поведение . 37 : 32–5. дои : 10.1016/j.yebeh.2014.05.027 . ПМИД 24967698 . S2CID 8368944 .
- ^ Jump up to: а б Байер, SA; Альтман, Дж. (январь 1991 г.). «Развитие эндопириформного ядра и Клауструма в мозге крысы». Нейронаука . 45 (2): 391–412. дои : 10.1016/0306-4522(91)90236-h . ПМИД 1762685 . S2CID 14720827 .
- ^ Байзер Дж.С., Шервуд CC, Нунан М., Хоф PR (2014). «Сравнительная организация клауструма: что структура говорит нам о функции?» . Границы системной нейронауки . 8 : 117. дои : 10.3389/fnsys.2014.00117 . ПМК 4079070 . ПМИД 25071474 .
- ^ Матур Б.Н. (2014). «Клауструм в обзоре» . Границы системной нейронауки . 8 : 48. дои : 10.3389/fnsys.2014.00048 . ПМЦ 3983483 . ПМИД 24772070 .
- ^ Jump up to: а б Эдельштейн Л.Р., Денаро Ф.Дж. (сентябрь 2004 г.). «Клауструм: исторический обзор его анатомии, физиологии, цитохимии и функционального значения». Клеточная и молекулярная биология . 50 (6): 675–702. ПМИД 15643691 .
- ^ Бьюкенен К.Дж., Джонсон Дж.И. (май 2011 г.). «Разнообразие пространственных взаимоотношений ворот и островка в ветвях радиации млекопитающих». Анналы Нью-Йоркской академии наук . 1225 Приложение 1 (S1): E30-63. Бибкод : 2011NYASA1225E..30B . дои : 10.1111/j.1749-6632.2011.06022.x . ПМИД 21599698 . S2CID 2245096 .
- ^ Шерк, Хелен (2014). «Физиология клостридий». Монастырь стр. 177–191. дои : 10.1016/B978-0-12-404566-8.00005-2 . ISBN 978-0-12-404566-8 .
- ^ Смитис-младший, Эдельштейн Л.Р., Рамачандран В.С. (2014). Клауструм: структурная, функциональная и клиническая нейробиология . Академическая пресса. ISBN 978-0-12-404566-8 . OCLC 861211388 . [ нужна страница ]
- ^ Фернандес-Миранда Х.К., Патхак С., Энг Дж., Джарбо К., Верстинен Т., Йе Ф.К., Ван Ю., Минц А., Боада Ф., Шнайдер В., Фридлендер Р. (август 2012 г.). «Волоконная трактография головного мозга высокого разрешения: нейроанатомическая проверка и нейрохирургическое применение». Нейрохирургия . 71 (2): 430–53. doi : 10.1227/NEU.0b013e3182592faa . ПМИД 22513841 . S2CID 12867524 .
- ^ ЛеВэй С. (декабрь 1986 г.). «Синаптическая организация клаустральных и коленчатых афферентов зрительной коры кошки» . Журнал неврологии . 6 (12): 3564–75. doi : 10.1523/JNEUROSCI.06-12-03564.1986 . ПМК 6568649 . ПМИД 2432202 .
- ^ Зингг Б., Хинтирян Х., Гоу Л., Сонг М.И., Бэй М., Биенковски М.С., Фостер Н.Н., Ямасита С., Боуман И., Тога А.В., Донг Х.В. (февраль 2014 г.). «Нейронные сети неокортекса мыши» . Клетка . 156 (5): 1096–111. дои : 10.1016/j.cell.2014.02.023 . ПМК 4169118 . ПМИД 24581503 .
- ^ Маркович Х.Дж., Ирле Э., Банг-Олсен Р., Флиндт-Эгебак П. (июнь 1984 г.). «Клаустральные эфференты лимбической коры кошки, изученные с помощью методов ретроградного и антероградного отслеживания». Нейронаука . 12 (2): 409–25. дои : 10.1016/0306-4522(84)90062-9 . ПМИД 6462456 . S2CID 21613309 .
- ^ Смит Дж.Б., Аллоуэй К.Д. (декабрь 2010 г.). «Функциональная специфика межполушарных связей у крысы: межполушарная связь между отдельными участками моторной коры» . Журнал неврологии . 30 (50): 16832–44. doi : 10.1523/JNEUROSCI.4438-10.2010 . ПМК 3010244 . ПМИД 21159954 .
- ^ Смит Дж.Б., Аллоуэй К.Д. (2014). «Межполушарные клаустральные цепи координируют сенсорные и моторные области коры, которые регулируют исследовательское поведение» . Границы системной нейронауки . 8 : 93. дои : 10.3389/fnsys.2014.00093 . ПМК 4032913 . ПМИД 24904315 .
- ^ Jump up to: а б с д Шелтон, Эндрю; Оливер, Дэвид; Гримстведт, Иоахим; Лазарт, Иван; Капур, Ишаан; Клиффорд, Кентрос; Виттер, Менно; Батт, Саймон; Пакер, Адам (2022). «Отдельные нейроны и сети в клауструме интегрируют входные данные из широко распространённых корковых источников» . биоRxiv . дои : 10.1101/2022.05.06.490864 . S2CID 248672084 .
- ^ Jump up to: а б с д Марриотт, Брайан А.; Делай, Элисон Д.; Захаси, Райан; Джексон, Джесси (2021). «Топографические градиенты определяют характер проекций ядра и оболочки клауструма у мышей» . Журнал сравнительной неврологии . 529 (7): 1607–1627. дои : 10.1002/cne.25043 . ISSN 0021-9967 . ПМЦ 8048916 . ПМИД 32975316 .
- ^ Ким, Джухён; Мэтни, Шанель Дж.; Рот, Ричард Х.; Браун, Соланж П. (20 января 2016 г.). «Синапсическая организация нейрональных цепей клауструма» . Журнал неврологии . 36 (3): 773–784. doi : 10.1523/JNEUROSCI.3643-15.2016 . ISSN 0270-6474 . ПМК 4719014 . ПМИД 26791208 .
- ^ Орман, Рена (01 ноября 2015 г.). «Клауструм: случай направленной, возбуждающей внутренней связи у крысы» . Журнал физиологических наук . 65 (6): 533–544. дои : 10.1007/s12576-015-0391-6 . ISSN 1880-6562 . ПМЦ 10717944 . ПМИД 26329935 . S2CID 255605784 .
- ^ Трамбле, Робин; Ли, Сухён; Руди, Бернардо (20 июля 2016 г.). «ГАМКергические интернейроны в неокортексе: от клеточных свойств к цепям» . Нейрон . 91 (2): 260–292. дои : 10.1016/j.neuron.2016.06.033 . ПМЦ 4980915 . ПМИД 27477017 .
- ^ Ван, Цюаньсинь; Ван, Юн; Куо, Сянь-Чи; Се, Пэн; Куанг, Сюли; Хирокава, Карла Э.; Наэми, Мейтам; Яо, Шэньцинь; Мэллори, Мэтт; Уэллетт, Бен; Леснар, Фил; Ли, Яояо; Да, Мин; Чен, Чао; Сюн, Вэй (28 февраля 2023 г.). «Региональные и специфичные для каждого типа клеток афферентные и эфферентные проекции клауструма мыши». Отчеты по ячейкам . 42 (2): 112118. doi : 10.1016/j.celrep.2023.112118 . ISSN 2211-1247 . PMC 10415534. PMID 36774552 .
- ^ Jump up to: а б Кадир, Хоуман; Стюарт, Брент В.; ВанРизин, Джонатан В.; Ву, Цион; Чен, Шуо; Семинович, Дэвид А.; Матур, Брайан Н. (20 декабря 2022 г.). «Клауструм мыши синаптически соединяет мотивы корковой сети» . Отчеты по ячейкам . 41 (12): 111860. doi : 10.1016/j.celrep.2022.111860 . ISSN 2211-1247 . ПМЦ 9838879 . ПМИД 36543121 .
- ^ Граф, Мартин; Наир, Адитья; Вонг, Келли Л.Л.; Тан, Янься; Августин, Джордж Дж. (01 июля 2020 г.). «Идентификация типов клаустральных нейронов мыши на основе их внутренних электрических свойств» . эНейро . 7 (4). doi : 10.1523/ENEURO.0216-20.2020 . ISSN 2373-2822 . ПМК 7405070 . ПМИД 32527746 .
- ^ Эрвин, Сара Р.; Бристоу, Брианна Н; Салливан, Кейтлин Э; Кендрик, Ренни М; Марриотт, Брайан; Ван, Лихуа; Клементс, Джоди; Лемир, Эндрю Л; Джексон, Джесси; Цембровский, Марк С (16 августа 2021 г.). Мао, Тяньи; Уэстбрук, Гэри Л; Чжан, Ли И (ред.). «Пространственно структурированные подтипы возбуждающих нейронов и проекции клауструма» . электронная жизнь . 10 : е68967. doi : 10.7554/eLife.68967 . ISSN 2050-084X . ПМЦ 8367382 . ПМИД 34397382 .
- ^ Наир, Адитья; Тео, Юэ Ян; Августин, Джордж Дж.; Граф, Мартин (11 июля 2023 г.). «Функциональная логика высвобождения нейромедиатора в холинергическом пути переднего мозга» . Труды Национальной академии наук . 120 (28): e2218830120. Бибкод : 2023PNAS..12018830N . дои : 10.1073/pnas.2218830120 . ISSN 0027-8424 . ПМЦ 10334726 . ПМИД 37399414 .
- ^ Грасби К., Разговор A (март 2013 г.). «Передняя клауструм и обучение изменению пространственного положения у крыс». Исследования мозга . 1499 : 43–52. дои : 10.1016/j.brainres.2013.01.014 . ПМИД 23318254 . S2CID 19605350 .
- ^ Норимото и др. (2020). «Клауструм у рептилий и ее роль в медленноволновом сне» . Природа . 578 (7795): 413–418. Бибкод : 2020Natur.578..413N . дои : 10.1038/s41586-020-1993-6 . hdl : 21.11116/0000-0008-09DB-8 . ПМИД 32051589 . S2CID 256820426 .
- ^ Jump up to: а б Нарикё и др. (2020). «Клауструм координирует корковую медленноволновую активность». Природная неврология . 23 (6): 741–753. дои : 10.1038/s41593-020-0625-7 . ПМИД 32393895 . S2CID 256840965 .
- ^ Габор, Эндрю Дж.; Пил, Талмейдж Л. (ноябрь 1964 г.). «Изменения поведения после стимуляции Клауструма кошки». Электроэнцефалография и клиническая нейрофизиология . 17 (5): 513–519. дои : 10.1016/0013-4694(64)90181-6 . ПМИД 14229851 .
- ^ Сильва Дж., Джейкоб С., Мело К., Алвес Д., Коста Д. (июнь 2018 г.). «Признак Клауструма у ребенка с рефрактерным эпилептическим статусом после лихорадочного заболевания: почему это происходит?». Acta Neurologica Belgica . 118 (2): 303–305. дои : 10.1007/s13760-017-0820-9 . ПМИД 28741106 . S2CID 32771124 .
- ^ Мелетти С, Слонкова Дж, Марецкова И, Монти Г, Спеккио Н, Хон П, Джованнини Г, Марсиан В, Кьяри А, Крупа П, Пьетрафуза Н, Беранкова Д, Бар М (октябрь 2015 г.). «Повреждение Клауструма и рефрактерный эпилептический статус после лихорадочного заболевания» . Неврология . 85 (14): 1224–32. дои : 10.1212/WNL.0000000000001996 . ПМЦ 4607596 . ПМИД 26341869 .
- ^ Шаплеске Дж., Росселл С.Л., Читнис XA, Саклин Дж., Симмонс А., Буллмор Э.Т., Вудрафф П.В., Дэвид А.С. (декабрь 2002 г.). «Вычислительная морфометрическая МРТ-исследование шизофрении: эффекты галлюцинаций» . Кора головного мозга . 12 (12): 1331–41. дои : 10.1093/cercor/12.12.1331 . ПМИД 12427683 . S2CID 33360335 .
- ^ Jump up to: а б Каселла Н.Г., Гернер Г.Дж., Филдстоун СК, Сава А., Шретлен DJ (декабрь 2011 г.). «Область островка-клаустра и бред при шизофрении». Исследования шизофрении . 133 (1–3): 77–81. doi : 10.1016/j.schres.2011.08.004 . ПМИД 21875780 . S2CID 45564142 .
- ^ Jump up to: а б Бикель, Стефан; Парвизи, Йозеф (август 2019 г.). «Электрическая стимуляция Клауструма человека» . Эпилепсия и поведение . 97 : 296–303. дои : 10.1016/j.yebeh.2019.03.051 . ПМИД 31196825 . S2CID 182952015 .
- ^ «Claustrum Nostrum: нет переключателя включения/выключения сознания - Skywritings» . generic.wordpress.soton.ac.uk . Проверено 2 декабря 2023 г.
- ^ Барретт, Фредерик С.; Криммель, Сэмюэл Р.; Гриффитс, Роланд Р.; Семинович, Дэвид А.; Матур, Брайан Н. (сентябрь 2020 г.). «Псилоцибин резко изменяет функциональные связи Клауструма с мозговыми сетями, которые поддерживают восприятие, память и внимание» . НейроИмидж . 218 : 116980. doi : 10.1016/j.neuroimage.2020.116980 . ISSN 1095-9572 . ПМЦ 10792549 . ПМИД 32454209 .
- ^ Ютса Дж., Хорн А., Сюй Дж., Фокс, доктор медицинских наук (август 2018 г.). «Локализация паркинсонизма по очаговым поражениям головного мозга» . Мозг . 141 (8): 2445–2456. дои : 10.1093/brain/awy161 . ПМК 6061866 . ПМИД 29982424 .
- ^ Ню, М.; Касаи, А.; Танума, М.; Сейрики, К.; Игараси, Х.; Куваки, Т.; Нагаясу, К.; Мияджи, К.; Уэно, Х.; Танабэ, В.; Со, К.; Ёкояма, Р.; Окубо, Дж.; Назад, Ю.; Хаяшида, М.; Иноуэ, К.И.; Такада, М.; Ямагучи, С.; Наказава, Т.; Канеко, С.; Окуно, Х.; Яманака, А.; Хашимото, Х. (2022). «Клауструм обеспечивает двунаправленный и обратимый контроль реакций тревоги, вызванных стрессом» . Достижения науки . 8 (11): eabi6375. Бибкод : 2022SciA....8I6375N . дои : 10.1126/sciadv.abi6375 . ПМЦ 8932664 . ПМИД 35302853 .
Новостная статья: « Отключение определенных клеток мозга может усилить стресс и реакцию тревоги, показывают исследования» . УПИ . Проверено 27 апреля 2022 г. - ^ Олсон Ч.Р., Грейбил А.М. (1980). «Сенсорные карты Клауструма кошки». Природа . 288 (5790): 479–481. Бибкод : 1980Natur.288..479O . дои : 10.1038/288479a0 . ПМИД 7442793 . S2CID 52530 .
- ^ Шерк Х., ЛеВэй С. (1981). «Зрительная клауструм: топография и свойства рецептивного поля у кошки». Наука . 212 (4490): 87–89. Бибкод : 1981Sci...212...87S . дои : 10.1126/science.7209525 . ПМИД 7209525 .
- ^ Смит Дж.Б., Чакрабарти С., Мауэри Т.М., Аллоуэй К.Д. (2022). «Конвергенция соматосенсорных и моторных корковых проекций передней лапы в полосатом теле, клауструме, таламусе и ядрах моста кошек». Структура и функции мозга . 227 (1): 361–379. дои : 10.1007/s00429-021-02405-6 . ПМИД 34665323 . S2CID 253984860 .