головной мозг
| головной мозг | |
|---|---|
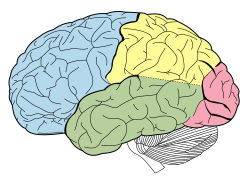
Доли коры головного мозга включают лобную (синюю), височную (зеленую), затылочную (красную) и теменную (желтую) доли. Мозжечок . (без обозначения) не является частью конечного мозга | |
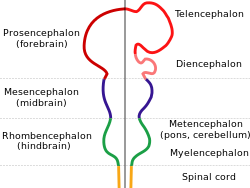 Схема, изображающая основные подразделения эмбрионального мозга позвоночных. | |
| Подробности | |
| Произношение | / ˈ s ɛ r ɪ b r ə m / , / s ɪ ˈ r iː b r ə m / |
| Артерия | Передний мозговой , средний мозговой , задний мозговой отделы |
| вена | Церебральные вены |
| Идентификаторы | |
| латинский | головной мозг |
| МеШ | D054022 |
| НейроЛекс ID | бирнлекс_1042 |
| ТА98 | A14.1.03.008 A14.1.09.001 |
| ТА2 | 5416 |
| ТД | Х3.11.03.6.00001 |
| ТО | Е5.14.1.0.2.0.12 |
| ФМА | 62000 |
| Анатомические термины нейроанатомии | |
Головной мозг ( мн .: мозг ), конечный мозг или конечный мозг. [1] — самый крупный отдел головного мозга, содержащий кору головного мозга (из двух полушарий головного мозга ), а также несколько подкорковых структур, включая гиппокамп , базальные ганглии и обонятельную луковицу . В человеческом мозге большой мозг является самой верхней областью центральной нервной системы . Головной мозг развивается внутриутробно из переднего мозга (прозэнцефалона). У млекопитающих дорсальный конечный мозг, или паллий , развивается в кору головного мозга, а вентральный конечный мозг, или субпаллий , становится базальными ганглиями . Головной мозг также разделен на примерно симметричные левое и правое полушария головного мозга.
При содействии мозжечка большой мозг управляет всеми произвольными действиями в организме человека.
Структура [ править ]

Головной мозг – самая большая часть головного мозга . В зависимости от положения животного он лежит либо спереди, либо сверху ствола мозга . У человека большой мозг является самым крупным и наиболее развитым из пяти основных отделов мозга.
Головной мозг состоит из двух полушарий головного мозга и их коры головного мозга (наружных слоев серого вещества ) и нижележащих областей белого вещества . [2] Его подкорковые структуры включают гиппокамп, базальные ганглии и обонятельную луковицу. Головной мозг состоит из двух полушарий головного мозга С-образной формы, отделенных друг от друга глубокой щелью, называемой продольной щелью .
Кора головного мозга [ править ]

Кора головного мозга , внешний слой серого вещества головного мозга, встречается только у млекопитающих. У более крупных млекопитающих, включая человека, поверхность коры головного мозга складывается, образуя извилины (гребни) и бороздки (борозды), которые увеличивают площадь поверхности . [3]
Кору головного мозга обычно разделяют на четыре доли : лобную , теменную , затылочную и височную . Доли классифицируются на основе лежащих над ними нейрокраниальных костей . [4] Меньшая доля — островковая доля , часть коры головного мозга, сложенная глубоко внутри латеральной борозды , которая отделяет височную долю от теменной и лобной долей, расположена в каждом полушарии мозга млекопитающих.
Полушария головного мозга [ править ]
Головной мозг разделен медиальной продольной щелью на два полушария головного мозга — правое и левое. Головной мозг организован контралатерально , т. е. правое полушарие контролирует и обрабатывает сигналы (преимущественно) с левой стороны тела, тогда как левое полушарие контролирует и обрабатывает сигналы (преимущественно) с правой стороны тела. [4] Согласно современным знаниям, это происходит из-за осевого перекручивания, которое происходит у раннего эмбриона . [5] существует сильная, но не полная двусторонняя симметрия , а латерализация имеет тенденцию увеличиваться с увеличением размера мозга. Между полушариями [6] Латерализация функций мозга рассматривает известные и возможные различия между ними.
Развитие [ править ]
У развивающегося эмбриона позвоночных нервная трубка подразделяется на четыре неразделенных отдела, которые затем развиваются в отдельные области центральной нервной системы ; это проэнцефалон ( передний мозг ), средний мозг ( средний мозг ), ромбовидный мозг ( задний мозг ) и спинной мозг . [7] Прозэнцефалон далее развивается в конечный мозг и промежуточный мозг . Дорсальный конечный мозг дает начало паллиуму (коре головного мозга у млекопитающих и рептилий), а вентральный конечный мозг образует базальные ганглии . Из промежуточного мозга развиваются таламус и гипоталамус , включая зрительные пузырьки (будущая сетчатка ). [8] Дорсальный конечный мозг затем образует два латеральных конечноцефальных пузырька, разделенных средней линией, которые развиваются в левое и правое полушария головного мозга. У птиц и рыб есть дорсальный конечный мозг, как и у всех позвоночных, но он обычно не имеет слоев и поэтому не считается корой головного мозга. Только слоистая цитоархитектура может считаться корой.
Функции [ править ]
Примечание . Поскольку головной мозг представляет собой крупный отдел со множеством подразделений и субрегионов, важно отметить, что в этом разделе перечислены функции, которым служит головной мозг в целом . Для получения дополнительной информации см. основные статьи о коре головного мозга и базальных ганглиях . Головной мозг — это основная часть мозга, контролирующая эмоции, слух, зрение, личность и многое другое. Он контролирует всю точность произвольных действий.
Верхние мотонейроны первичной моторной коры посылают свои аксоны в ствол мозга и спинной мозг для образования синапсов на нижних мотонейронах , которые иннервируют мышцы. Повреждение двигательных областей корой головного мозга может привести к определенным типам заболеваний двигательных нейронов . Повреждения такого рода приводят к потере мышечной силы и точности, а не к полному параличу .
Он функционирует как центр чувственного восприятия, памяти, мыслей и суждений; головной мозг также функционирует как центр произвольной двигательной активности.
Сенсорная обработка [ править ]
Первичные сенсорные области коры головного мозга получают и обрабатывают зрительную , слуховую , соматосенсорную , вкусовую и обонятельную информацию. Вместе с ассоциативными областями коры эти области мозга синтезируют сенсорную информацию в наше восприятие мира.
Обоняние [ править ]
Обонятельная луковица , отвечающая за обоняние, у большинства позвоночных занимает большую площадь головного мозга. Однако у людей эта часть мозга намного меньше и находится под лобной долей. Обонятельная сенсорная система уникальна, поскольку нейроны обонятельной луковицы посылают свои аксоны непосредственно в обонятельную кору головного мозга , а не сначала в таламус . Обоняние также является единственным чувством, которое представлено ипсилатеральной стороной мозга. Повреждение обонятельной луковицы приводит к потере обоняния (обоняния).
Язык и общение [ править ]
Речь и язык в основном относятся к отделам коры головного мозга. Двигательные части языка относятся к зоне Брока в лобной доле. Понимание речи осуществляется за счет зоны Вернике , расположенной на стыке височно-теменной доли. Эти две области соединены между собой большим трактом белого вещества — дугообразным пучком . Повреждение зоны Брока приводит к выраженной афазии (неплавной афазии), тогда как повреждение зоны Вернике приводит к рецептивной афазии (также называемой беглой афазией).
Обучение и память [ править ]
Формирование эксплицитной или декларативной (фактической) памяти происходит за счет гиппокампа и связанных с ним областей медиальной височной доли . Эта ассоциация была первоначально описана после того, как пациенту, известному как ХМ, хирургическим путем удалили левый и правый гиппокамп для лечения хронической височной эпилепсии. После операции у ХМ возникла антероградная амнезия , или неспособность формировать новые воспоминания.
Неявная или процедурная память , например, сложное двигательное поведение, затрагивает базальные ганглии.
Кратковременная или рабочая память задействует ассоциативные области коры головного мозга, особенно дорсолатеральную префронтальную кору , а также гиппокамп.
Другие животные [ править ]
У наиболее примитивных позвоночных — миксин и миног — головной мозг представляет собой сравнительно простую структуру, получающую нервные импульсы от обонятельной луковицы . У хрящевых и лопастных рыб , а также у земноводных имеется более сложное строение: головной мозг разделен на три отдельные области. Самая нижняя (или вентральная) область образует базальные ядра и содержит волокна, соединяющие остальную часть головного мозга с таламусом. Над ним, образуя латеральную часть головного мозга, находится палеопаллий , а самая верхняя (или дорсальная) часть называется архипаллием . У этих животных головной мозг по-прежнему в основном отвечает за обонятельные ощущения, в отличие от гораздо более широкого диапазона его функций у амниот . [9]
У лучепёрых рыб строение несколько иное. Внутренние поверхности латеральных и вентральных отделов головного мозга выбухают в желудочки ; они включают как базальные ядра, так и различные части мантия и могут иметь сложную структуру, особенно у костистых рыб . Дорсальная поверхность головного мозга мембранозная и не содержит нервной ткани . [9]
У амниот головной мозг становится все более крупным и сложным. У рептилий палеопаллий намного крупнее, чем у земноводных, и его рост привел к смещению базальных ядер в центральные области головного мозга. Как и у низших позвоночных, серое вещество обычно расположено под белым веществом , но у некоторых рептилий оно распространяется на поверхность, образуя примитивную кору, особенно в передней части мозга. [9]
У млекопитающих это развитие идет дальше, так что кора охватывает почти все полушария головного мозга, особенно у более развитых видов, например приматов . Палеопаллий выталкивается на вентральную поверхность мозга, где он становится обонятельными долями, а архипаллий переворачивается на медиальном дорсальном крае, образуя гиппокамп . У плацентарных млекопитающих , также развивается мозолистое тело дополнительно соединяющее два полушария. Сложные извилины мозговой поверхности (см. извилины , гирификация ) встречаются также только у высших млекопитающих. [9] Хотя у некоторых крупных млекопитающих (например, слонов) головной мозг особенно большой, дельфины — единственные виды (кроме человека), у которых головной мозг составляет до 2 процентов массы тела. [10]
Мозг птиц по сравнению с рептилиями увеличен так же, как и у млекопитающих. Увеличенный размер мозга птиц классически объяснялся увеличением базальных ганглиев , тогда как другие области оставались примитивными, но от этой точки зрения в значительной степени отказались. [11] Птицы, по-видимому, подверглись альтернативному процессу энцефализации . [12] поскольку они отличались от других архозавров , с немногими явными параллелями с тем, что испытывали млекопитающие и их предки -терапсиды .
Дополнительные изображения [ править ]
 Головной мозг. Боковое лицо. Глубокое рассечение.
Головной мозг. Боковое лицо. Глубокое рассечение. Головной мозг. Медиальное лицо. Глубокое рассечение.
Головной мозг. Медиальное лицо. Глубокое рассечение.


См. также [ править ]
Ссылки [ править ]
- ^ «БрейнИнфо» . braininfo.rprc.washington.edu .
- ^ Арнулд-Тейлор, Уильям (1998). Учебник анатомии и физиологии . Нельсон Торнс. п. 52. ИСБН 9780748736348 . Проверено 27 января 2015 г.
- ^ Анжуйн, Дж.; Котман, К. (1981). Принципы нейроанатомии . Нью-Йорк: Издательство Оксфордского университета. ISBN 9780195028850 . Проверено 25 января 2015 г.
- ↑ Перейти обратно: Перейти обратно: а б Росдал, Кэролайн; Ковальски, Мэри (2008). Учебник по основам сестринского дела (9-е изд.). Липпинкотт Уильямс и Уилкинс. п. 189. ИСБН 9780781765213 . Проверено 28 января 2015 г.
- ^ де Люссане, MHE; Оссе, JWM (2012). «Наследственное осевое скручивание объясняет контралатеральную переднюю часть и перекрест зрительных нервов у позвоночных». Биология животных . 62 (2): 193–216. arXiv : 1003.1872 . дои : 10.1163/157075611X617102 . S2CID 7399128 .
- ^ Эббесон, Свен О.Э.; Ито, Хиронобу (1980). «Двусторонние проекции сетчатки у черной пираны (Serrasalmus niger)». Ресурсы клеточных тканей . 213 (3): 483–495. дои : 10.1007/BF00237893 . ПМИД 7448850 . S2CID 2406618 .
- ^ Гилберт, Скотт Ф. (2014). Биология развития (10-е изд.). Сандерленд, Массачусетс: Синауэр. ISBN 978-0-87893-978-7 .
- ^ Кандел, Эрик Р., изд. (2006). Принципы нейронауки (5-е изд.). Эпплтон и Ланге: МакГроу-Хилл. ISBN 978-0-07-139011-8 .
- ↑ Перейти обратно: Перейти обратно: а б с д Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 536–543. ISBN 0-03-910284-Х .
- ^ Т.Л. Бринк (2008). «Блок 4: Нервная система». Психология: подход, дружественный к студентам (PDF) . п. 62.
- ^ Джарвис Э.Д., Гюнтюркюн О., Брюс Л. и др. (2005). «Птичий мозг и новое понимание эволюции мозга позвоночных» . Нат. Преподобный Нейроски . 6 (2): 151–9. дои : 10.1038/nrn1606 . ПМК 2507884 . ПМИД 15685220 .
- ^ Эмери, Нью-Джерси (29 января 2006 г.). «Когнитивная орнитология: эволюция птичьего интеллекта» . Филос. Пер. Р. Сок. Лонд. Б Биол. Наука . 361 (1465): 23–43. дои : 10.1098/rstb.2005.1736 . ПМК 1626540 . ПМИД 16553307 .