Нервная трубка
Эта статья нуждается в дополнительных цитатах для проверки . ( ноябрь 2016 г. ) |
| Нервная трубка | |
|---|---|
 Поперечный срез половины куриного эмбриона сорокапятичасовой инкубации. Дорсальная (задняя) поверхность эмбриона находится вверху этой страницы, а вентральная (передняя) поверхность — внизу. (Нервная трубка выделена зеленым цветом.) | |
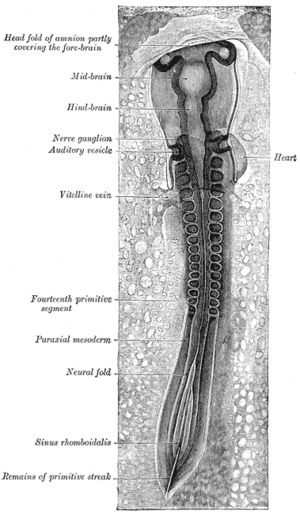 Куриный эмбрион после тридцатитрехчасовой инкубации, вид со спины (30-кратное увеличение) | |
| Подробности | |
| Этап Карнеги | 10 |
| Предшественник | Нейронная канавка |
| Дает начало | Центральная нервная система ( головной и спинной мозг ) |
| Идентификаторы | |
| латинский | невральная трубка, невральная трубка |
| МеШ | D054259 |
| ТО | Tube_by_E5.14.1.0.0.0.1 E5.14.1.0.0.0.1 |
| Анатомическая терминология | |
У развивающихся хордовых (включая позвоночных ) нервная трубка является эмбриональным предшественником центральной нервной системы , которая состоит из головного и спинного мозга . Нервная борозда постепенно углубляется по мере того, как нервная складка поднимается, и в конечном итоге складки встречаются и сливаются по средней линии и превращают борозду в закрытую нервную трубку. У человека закрытие нервной трубки обычно происходит к четвертой неделе беременности (28-й день после зачатия).

Развитие [ править ]
Нервная трубка развивается двумя способами: первичная нейруляция и вторичная нейруляция .
Первичная нейруляция делит эктодерму на три типа клеток:
- Внутренне расположенная нервная трубка
- Наружно расположенный эпидермис
- Клетки нервного гребня , которые развиваются в области между нервной трубкой и эпидермисом, но затем мигрируют в новые места.
- Первичная нейруляция начинается после формирования нервной пластинки. Края нервной пластинки начинают утолщаться и подниматься вверх, образуя нервные складки. Центр нервной пластинки остается заземленным, что позволяет сформировать U-образную нервную борозду. Эта нервная борозда устанавливает границу между правой и левой сторонами эмбриона. Нервные складки сжимаются к средней линии эмбриона и сливаются вместе, образуя нервную трубку. [1]
- При вторичной нейруляции клетки нервной пластинки образуют шнуровидную структуру, которая мигрирует внутри эмбриона и впадает, образуя трубку.
Каждый организм в разной степени использует первичную и вторичную нейруляцию.
- Нейруляция у рыб протекает только через вторичную форму.
- У видов птиц задние участки трубки развиваются за счет вторичной нейруляции, а передние участки - за счет первичной нейруляции.
- У млекопитающих вторичная нейруляция начинается около 35-го сомита .
Нервные трубки млекопитающих закрываются в голове в порядке, противоположном тому, который закрывается в туловище.
- В голове:
- Клетки нервного гребня мигрируют
- Нервная трубка закрывается
- Вышележащая эктодерма закрывается
- В багажнике:
- Вышележащая эктодерма закрывается
- Нервная трубка закрывается
- Клетки нервного гребня мигрируют
Структура [ править ]

развивается в отдельные области центральной нервной системы Каждый из четырех подразделений нервной трубки в конечном итоге в результате деления нейроэпителиальных клеток : передний мозг (передний мозг), средний мозг (мезенцефалон), задний мозг (ромбэнцефалон) и спинной мозг .
- Прозэнцефалон ( далее развивается в конечный мозг мозг ) и промежуточный мозг ( зрительные пузырьки и гипоталамус ).
- остается Средний мозг средним мозгом.
- Ромбэнцефалон продолговатый развивается в задний мозг ( мост и мозжечок ) и мозг ( продолговатый мозг ).
На короткое время нервная трубка открывается как краниально , так и каудально . Эти отверстия, называемые нейропорами , у человека закрываются на четвертой неделе. Неправильное закрытие нейропор может привести к дефектам нервной трубки, таким как анэнцефалия или расщелина позвоночника .
В дорсальной части нервной трубки находится крыловая пластинка , связанная преимущественно с ощущением . Вентральная мышечным часть нервной трубки содержит базальную пластинку , которая в первую очередь связана с моторным (т. е. ) контролем.
Спинной мозг развивается из задней нервной трубки. По мере развития спинного мозга клетки, составляющие стенку нервной трубки, пролиферируют и дифференцируются в нейроны и глию спинного мозга. Дорсальные ткани будут связаны с сенсорными функциями, а вентральные ткани — с двигательными функциями. [2]
Дорсально-вентральный рисунок [ править ]
Структура нервной трубки вдоль дорсально-вентральной оси образует определенные компартменты нервных клеток-предшественников, которые приводят к различным классам нейронов. [3] Согласно French flag модели морфогенеза , этот паттерн возникает на ранних стадиях развития и является результатом активности нескольких секретируемых сигнальных молекул. Sonic hedgehog (Shh) является ключевым игроком в формировании паттерна вентральной оси, тогда как костные морфогенные белки (BMPs) и члены семейства Wnt играют важную роль в формировании паттерна дорсальной оси. [4] Другие факторы, которые, как показано, предоставляют информацию о положении нервным клеткам-предшественникам, включают факторы роста фибробластов (FGF) и ретиноевую кислоту . Ретиноевая кислота необходима вентрально вместе с Shh для индукции Pax6 и Olig2 во время дифференцировки мотонейронов. [5]
Во время раннего развития нервной трубки устанавливаются три основных типа вентральных клеток: пластинки дна клетки , которые формируются на вентральной средней линии на стадии нервной складки; а также более дорсально расположенные мотонейроны и интернейроны . [3] Эти типы клеток определяются секрецией Shh из хорды (расположенной вентрально от нервной трубки), а затем из клеток пластинки дна. [6] Shh действует как морфоген, что означает, что он действует в зависимости от концентрации, определяя типы клеток по мере их удаления от источника. [7]
Ниже приводится предполагаемый механизм того, как Shh паттернирует вентральную нервную трубку: создается градиент Shh, который контролирует экспрессию группы гомеодомена (HD) и основных транскрипционных факторов Helix-Loop-Helix (bHLH). Эти факторы транскрипции сгруппированы в два класса белков в зависимости от того, как на них влияет Shh. Класс I ингибируется Shh, тогда как класс II активируется Shh. Эти два класса белков затем перекрестно регулируют друг друга, создавая более четкие границы экспрессии. Различные комбинации экспрессии этих факторов транскрипции вдоль дорсально-вентральной оси нервной трубки ответственны за создание идентичности нейрональных клеток-предшественников. [4] Из этих нейрональных клеток-предшественников in vitro формируются пять молекулярно различных групп вентральных нейронов. Кроме того, положение, в котором эти группы нейронов генерируются in vivo, можно предсказать по концентрации Shh, необходимой для их индукции in vitro. [8] Исследования показали, что нейрональные предшественники могут вызывать разные реакции в зависимости от продолжительности воздействия Shh, причем более длительное время воздействия приводит к образованию большего количества вентральных типов клеток. [9] [10]
На дорсальном конце нервной трубки BMP отвечают за формирование нейронного паттерна. BMP первоначально секретируется из вышележащей эктодермы. Вторичный сигнальный центр затем устанавливается в верхней пластинке, самой дорсальной структуре нервной трубки. [1] BMP из дорсального конца нервной трубки, по-видимому, действует таким же зависимым от концентрации образом, как Shh в вентральном конце. [11] Это было показано с использованием мутантов рыбок данио, которые обладали разной степенью сигнальной активности BMP. Исследователи наблюдали изменения в дорсально-вентральном паттерне: например, у рыбок данио с дефицитом определенных BMP наблюдалась потеря дорсальных сенсорных нейронов и расширение интернейронов. [12]

См. также [ править ]
Ссылки [ править ]
![]() текст В эту статью включен общедоступный со страницы 50 20-го издания «Анатомии Грея» (1918 г.).
текст В эту статью включен общедоступный со страницы 50 20-го издания «Анатомии Грея» (1918 г.).
- ↑ Перейти обратно: Перейти обратно: а б Гилберт, Скотт Ф. Биология развития, восьмое издание. Сандерленд, Массачусетс: Sinauer Associates, Inc., 2006.
- ^
 В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (8 июня 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 13.1. Эмбриологическая перспектива. ISBN 978-1-947172-04-3 .
В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (8 июня 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 13.1. Эмбриологическая перспектива. ISBN 978-1-947172-04-3 . - ↑ Перейти обратно: Перейти обратно: а б Джесселл ТМ (2000). «Спецификация нейронов спинного мозга: индуктивные сигналы и транскрипционные коды». Нат преподобный Жене . 1 (1): 20–9. дои : 10.1038/35049541 . ПМИД 11262869 . S2CID 205012382 .
- ↑ Перейти обратно: Перейти обратно: а б Уллоа Ф, Марти Э (2010). «Wnt выиграл войну: антагонистическая роль Wnt по отношению к Shh контролирует дорсо-вентральное паттернирование нервной трубки позвоночных» . Динамика развития . 239 (1): 69–76. дои : 10.1002/dvdy.22058 . ПМИД 19681160 . S2CID 205766310 .
- ^ Дустер Дж. (2008). «Синтез ретиноевой кислоты и передача сигналов на раннем органогенезе» . Клетка . 134 (6): 921–931. дои : 10.1016/j.cell.2008.09.002 . ПМК 2632951 . ПМИД 18805086 .
- ^ Паттен И, Плачек М (2000). «Роль ежа Соника в формировании рисунка нервной трубки» . Cell Mol Life Sci . 57 (12): 1695–708. дои : 10.1007/pl00000652 . ПМЦ 11146859 . ПМИД 11130176 . S2CID 20950575 .
- ^ Дессо Э., МакМахон А.П., Бриско Дж. (2008). «Формирование паттерна в нервной трубке позвоночных: транскрипционная сеть, регулируемая морфогеном звукового ежа» . Разработка . 135 (15): 2489–2503. дои : 10.1242/dev.009324 . ПМИД 18621990 .
- ^ Эриксон Дж., Бриско Дж., Рашбасс П., ван Хейнинген В., Джесселл Т.М. (1997). «Градуированная передача сигналов Sonic hedgehog и спецификация судьбы клеток в вентральной части нервной трубки». Холодный источник Harb Symp Quant Biol . 62 : 451–466. дои : 10.1101/SQB.1997.062.01.053 . ПМИД 9598380 .
- ^ Стаматаки Д., Уллоа Ф., Цони С.В., Минетт А., Бриско Дж. (2005). «Градиент активности Gli опосредует ступенчатую передачу сигналов Sonic hedgehog в нервной трубке» . Генс Дев . 19 (5): 626–641. дои : 10.1101/gad.325905 . ПМК 551582 . ПМИД 15741323 .
- ^ Дессо Э., Ян Л.Л., Хилл К. и др. (ноябрь 2007 г.). «Интерпретация градиента морфогена звукового ежа с помощью механизма временной адаптации» (PDF) . Природа . 450 (7170): 717–20. Бибкод : 2007Natur.450..717D . дои : 10.1038/nature06347 . hdl : 2027.42/62511 . ПМИД 18046410 . S2CID 4419025 .
- ^ Уилсон Л., Мэден М. (2005). «Механизмы формирования дорсовентрального паттерна в нервной трубке позвоночных» . Биология развития . 282 (1): 1–13. дои : 10.1016/j.ydbio.2005.02.027 . ПМИД 15936325 .
- ^ Нгуен В.Х., Траут Дж., Коннорс С.А., Андерманн П., Вайнберг Э., Маллинз MC (2000). «Типы дорсальных и промежуточных нейрональных клеток спинного мозга определяются сигнальным путем BMP» . Разработка . 127 (6): 1209–1220. дои : 10.1242/dev.127.6.1209 . ПМИД 10683174 .
Внешние ссылки [ править ]
- Швейцарская эмбриология (из UL , UB и UF ) i periodembry/carnegie03
- Эмбриология в UNSW Notes/week3_5
- Схема на сайте Embryology.med.unsw.edu.au.
- Схема на сайте Brainviews.com.
| Базы данных органов управления : Национальные |
|---|