Напольная плита
Эта статья нуждается в дополнительных цитатах для проверки . ( декабрь 2009 г. ) |
| Напольная плита | |
|---|---|
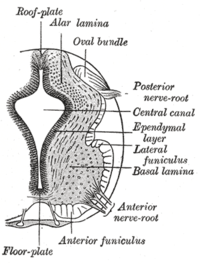 Пластинка дна разделяет левую и правую базальные пластинки развивающейся нервной трубки . | |
| Подробности | |
| Предшественник | Нотохорда |
| Система | Нервная система |
| Анатомическая терминология | |
Донная пластинка представляет собой структуру, неотъемлемую часть развивающейся нервной системы позвоночных организмов. Пластинка дна , расположенная на вентральной срединной линии эмбриональной нервной трубки , представляет собой специализированную глиальную структуру, которая охватывает переднезаднюю ось от среднего мозга до хвостовых отделов. Было показано, что пластинка дна консервативна среди позвоночных, таких как рыбки данио и мыши, с гомологичными структурами у беспозвоночных, таких как плодовая мушка Drosophila и нематода C. elegans . Функционально эта структура служит организатором вентрализации тканей эмбриона, а также направляет позиционирование и дифференцировку нейронов вдоль дорсовентральной оси нервной трубки. [1] [2] [3]
Индукция
[ редактировать ]Индукция пластинки дна во время эмбриогенеза эмбрионов позвоночных широко изучалась у кур и рыбок данио и происходит в результате сложной сигнальной сети между тканями, детали которой еще предстоит полностью уточнить. В настоящее время существует несколько конкурирующих направлений мысли. Во-первых, дифференцировка пластинки дна может быть опосредована индуктивной передачей сигналов от подлежащей хорды , аксиальной мезодермальной сигнальной структуры. Это подтверждается экспериментально на курах, у которых индукция пластинки дна, а также ассоциативная дифференцировка вентральной нервной ткани опосредованы секретируемой сигнальной молекулой sonic hedgehog (Shh). Shh выражается в градиенте с самой высокой концентрацией, локализованной в хорде и пластинке дна. Эксперименты по трансплантации тканей in vitro показывают, что удаление этой молекулы предотвращает дифференцировку пластинки дна, тогда как ее эктопическая экспрессия индуцирует дифференцировку клеток пластинки дна. [4] Альтернативная точка зрения предполагает, что клетки пластинки дна нервной трубки происходят из клеток-предшественников, которые мигрируют непосредственно из осевой мезодермы. Благодаря экспериментам с гибридами кур и перепелов, а также экспериментам по генетическому взаимодействию на рыбках данио выяснилось, что клетки хорды и пластинки дна происходят от общего предшественника. Более того, у рыбок данио передача сигналов Nodal необходима для дифференцировки клеток медиальной пластинки дна, тогда как Shh является расходным материалом. Эти данные могут указывать на то, что механизм индукции пластинки дна у амниот и анамниот различен. [5] Чтобы примирить эти различия, у цыплят была предложена двухрежимная модель индукции. В этой модели исключительно эктодермальные клетки индуцируются превращением в медиальную пластинку дна во время гаструляции с помощью прехордальной мезодермы, возможно, посредством передачи сигналов Nodal. Позже в развитии во время нейруляции расширенный контакт и взаимодействие между хордой и обреченными клетками пластинки дна вызывает дифференцировку, указывая на кооперативный эффект между передачей сигналов Nodal и Shh. [6]
Наведение аксона
[ редактировать ]В развитии центральной нервной системы решающее значение имеет решение нейрона пересекать или не пересекать срединную линию. У позвоночных этот выбор опосредован пластинкой дна и позволяет эмбриону успешно развивать левую и правую половины тела по отношению к нервной ткани. Например, хотя ипсилатеральные нейроны не пересекают срединную линию, комиссуральные нейроны пересекают срединную линию, образуя единственную спайку. Эти конкретные нейроны развиваются в дорсальной области нервной трубки и перемещаются вентрально к пластинке дна. Достигнув пластинки дна, комиссуральные нейроны пересекают структуру и выходят на противоположную сторону нервной трубки, после чего они выступают вперед или назад внутри трубки. [7]

- Нетрины: Нетрины представляют собой белки, экспрессируемые и секретируемые клетками пластинки дна. Эксперименты с использованием экстрактов пластинки дна и комиссуральных нейронов, встроенных в коллагеновый матрикс, показывают притяжение нейронов к пластинке дна in vitro . [8] Более того, выделение и трансфекция нетрина-1 и нетрина-2, двух секретируемых белков, в клетки Cos имеет аналогичные эффекты. [9] Дальнейшие исследования подтвердили, что нетрины действуют как белки-аттрактанты в дополнение к Shh, направляя комиссуральные аксоны к пластинке дна. [10] Нетрины секретируются клетками пластинки дна и функционируют, связываясь с аксонным рецептором DCC хемотаксическим способом. Недавние исследования поставили под сомнение эту модель и показали, что Netrin-1, происходящий из пластинки дна, необязателен для пересечения средней линии и что Netrin-1 преимущественно происходит из предшественников желудочковой зоны. [11]
- Slit: Slit представляет собой секретируемый лиганд, экспрессирующийся в пластинке дна и ингибирующий пересечение аксонов нервной трубки. В то время как нетрины привлекают комиссуральные нейроны к средней линии, щелевые белки отталкивают и вытесняют нейроны из средней линии. Поскольку аксоны, которым не суждено пересечь срединную линию, проходят через нервную трубку, они отталкиваются лигандной щелью, которая экспрессируется в клетках пластинки дна. Slit действует через свои рецепторы Roundabout (Robo) 1 и 2. Это взаимодействие ингибирует хемотаксис, обеспечиваемый путем Netrin/DCC. Однако Robo-3 (Rig-1) активируется во время роста комиссуральных аксонов во время миграции к пластинке дна, что изолирует Robo-1/2 внутри клетки внутри везикул. Следовательно, путь притяжения Netrin/DCC доминирует над путем отталкивания Slit/Robo , и аксон может расти к средней линии и входить в пластинку дна. При входе, посредством еще не до конца понятного механизма, Робо-3 становится подавленным, что освобождает и активирует Робо-1/2, эффективно отталкивая нейрон от средней линии пластинки дна. Благодаря этому сложному перекрестному взаимодействию Slit, Robo-1/2 и Robo-3 комиссуральные аксоны направляются к средней линии, чтобы пересечь нервную трубку и не допускаются обратного пересечения. [12]
Сигнальные молекулы, управляющие ростом и проекцией комиссуральных нейронов, имеют хорошо изученные гомологи у беспозвоночных. В пути хемоаттрактции нетрина/DCC гомологами C. elegans являются Unc-6/Unc-40 и Unc-5, тогда как гомологами дрозофилы являются Netrin-A и Netrin-B/Frazzled и Dunc5. В хемоотталкивающем пути Slit/Robo гомологами C. elegans являются Slt-1/Sax-3, тогда как гомологи Drosophila также известны как Slit/Robo(1-3). [7]
Картирование глиальной судьбы
[ редактировать ]В центральной нервной системе (ЦНС) общее картирование судеб клеток обычно направляется сигнальным путем морфогена sonic hedgehog (Shh). В спинном мозге Shh направляется как хордой, так и областями пластинки дна, что в конечном итоге управляет организацией популяций нейрональных и глиальных предшественников. Специфические глиальные популяции, на которые влияет Shh в этих двух регионах, включают клетки-предшественники олигодендроцитов (OPC), олигодендроциты , NG2+ клетки, микроглию и астроциты . [13] Область пластинки дна (FP) спинного мозга индивидуально способствует глиогенезу или образованию глиальных клеток. Традиционно клетки-предшественники переходят от фазы экспансии своих предшественников к нейрогенной фазе и, в конечном счете, к глиогенной фазе. На глиогенной фазе бывшие клетки-предшественники могут затем стать астроцитами, олигодендроцитами или другими более специализированными типами глиальных клеток. Недавно были предприняты попытки использовать условный мутагенез для избирательной инактивации пути Shh, особенно в области FP, чтобы идентифицировать различные роли молекул, участвующих в судьбе клеток олигодендроцитов. Олигодендроциты — это клетки, ответственные за миелинизирование аксонов в ЦНС.
Shh регулирует процессинг Gli посредством двух белков, Ptch1 и Smo . [14] Когда Shh не активен, Ptch1 отвечает за подавление этого пути посредством ингибирования Smo. Smo имеет решающее значение для общей передачи сигнала по пути Shh. Если Smo ингибируется, путь Shh также неактивен, что в конечном итоге подавляет глиогенез. Специфические факторы, такие как Gli3, необходимы для судьбы клеток олигодендроцитов. Поскольку Shh регулирует процессинг Gli, если Smo нарушается или ингибируется Ptch1, это инактивирует путь Shh и предотвращает процессинг Gli, что нарушает картирование судеб глиальных клеток. Передача сигналов Shh в области FP очень важна, поскольку она должна быть активной для того, чтобы произошел глиогенез. Если Shh инактивирован в области FP и активирован в других регионах спинного мозга, таких как домены Dbx или pMN, глиогенез нарушается. Но когда Shh активен в области FP, активируется глиогенез и глиальные клетки начинают мигрировать к месту назначения для функционирования.
Травма спинного мозга и регенерация аксонов
[ редактировать ]Область пластинки дна помогает в наведении аксонов, картировании судеб глии и эмбриогенезе . Если эта область спинного мозга повреждается, могут возникнуть серьезные осложнения для всех функций этой области, а именно ограничение пролиферации и производства глиальных клеток, ответственных за миелинизацию и фагоцитоз в ЦНС. Травма спинного мозга (ТСМ) также чаще всего приводит к оголению или разрыву аксонов. Передача сигналов Wnt является распространенным сигнальным путем, участвующим в случаях травм. Передача сигналов Wnt регулирует регенерацию после травмы спинного мозга. Сразу после травмы экспрессия Wnt резко возрастает. [15] Наведение аксона осуществляется Netrin-1. [8] в ФП-области спинного мозга. Во время случаев повреждения, особенно случаев разрыва аксонов, передача сигналов Wnt активируется, и аксоны начинают инициировать регенерацию, и аксоны перенаправляются через области FP с использованием сигнальных путей Shh и Wnt.
Эпендимальные клетки спинного мозга также находятся в FP-области спинного мозга. Эти клетки представляют собой популяцию нервных стволовых клеток, ответственную за репопуляцию потерянных клеток во время травмы. Эти клетки обладают способностью дифференцироваться в глиальные популяции-предшественники. При травме в FP-области секретируется фактор Ахирин. Во время развития спинного мозга ахирин экспрессируется исключительно на эпендимальных стволовых клетках со скрытыми свойствами стволовых клеток и играет ключевую роль в развитии спинного мозга. В отсутствие ахирина стволовость этих эпендимальных клеток не регулируется. [16] Повреждение нарушает экспрессию и регуляцию ахирина, и клетки FP-области не могут быть должным образом восстановлены с помощью популяций эпендимальных стволовых клеток.
Ссылки
[ редактировать ]- ^ «Вулперт, Льюис. Принципы развития: 3-е издание. Oxford University Press, 2007».
- ^ «Гилберт, Скотт Ф. Принципы развития: 8-е издание. Sinauer Associates, Inc. 2006».
- ^ "Джесселл, Томас М. Спецификация нейронов в спинном мозге: индуктивные сигналы и транскрипционные коды. Nature Reviews Genetics. Октябрь 2000 г. (1)"
- ^ «Ямада, Т. Контроль структуры клеток в развивающейся нервной системе: поляризующая активность пластинки дна и хорды. Клетка, том 64, 635-647, 8 февраля 1991 г.»
- ^ «Страле, Уве и др. Спецификация пластины пола позвоночных: вариации на общие темы. Тенденции в генетике, том 20, № 3, март 2004 г.»
- ^ «Паттен, Иэн и др. Различные способы индукции пластинки дна у куриного эмбриона. 19 июня 2003 г. Развитие 130, 4809-4821»
- ^ Jump up to: а б «Гуань К.Л. и Рао Ю. Механизмы передачи сигналов, опосредующие реакцию нейронов на сигналы управления. Nat Rev Neurosci. Декабрь 2003 г.; 4 (12): 941-56».
- ^ Jump up to: а б «Серафини, Тито и др. Нетрины определяют семейство белков, способствующих росту аксонов, гомологичных C. elegans UNC-6. Cell, Vol. 79, 409-424. 12 августа 1994 г.»
- ^ «Кеннеди, Тимоти Э. и др. Нетрины являются диффузионными хемотропными факторами для комиссуральных аксонов в эмбриональном спинном мозге. Клетка, том 79, 425-435, 12 августа 1994 г.»
- ^ «Чаррон, Ф. и др. Морфоген sonic hedgehog представляет собой аксональный хемоаттрактант, который сотрудничает с нетрином-1 в наведении аксонов по средней линии. Cell. 4 апреля 2003 г.; 113 (1): 11-23».
- ^ Доминичи, К; Морено-Браво, Дж.А.; Пуиггрос, СР; Раппено, Q; Рама, Н; Вьег, П; Бернет , А; Мелен, П; Чедоталь, А. (18 мая 2017 г.). «Нетрин-1, полученный из пластинки дна, не требуется для проведения комиссуральных аксонов» . Природа . 545 (7654): 350–354. дои : 10.1038/nature22331 . ПМЦ 5438598 . ПМИД 28445456 .
- ^ «Лонг, Хуа и др. Консервативные роли щелевых и робо-белков в наведении срединных комиссуральных аксонов. Neuron, Vol. 42, 213–223, 22 апреля 2004 г.»
- ^ Ю, Кванха; МакГлинн, Шон; Матисе, Майкл П. (01 апреля 2013 г.). «Звуковой еж, полученный из напольной пластинки, регулирует судьбы глиальных и эпендимальных клеток в развивающемся спинном мозге» . Разработка . 140 (7): 1594–1604. дои : 10.1242/dev.090845 . ISSN 0950-1991 . ПМЦ 3596997 . ПМИД 23482494 .
- ^ Бай, К. Брайан; Ауэрбах, Войтек; Ли, Джун С.; Стивен, Дэниел; Джойнер, Александра Л. (октябрь 2002 г.). «Gli2, но не Gli1, необходим для начальной передачи сигналов Shh и эктопической активации пути Shh». Разработка . 129 (20): 4753–4761. ISSN 0950-1991 . ПМИД 12361967 .
- ^ Цзоу, Имин (2015). «Передача сигналов Wnt при травме спинного мозга». Передача сигналов Wnt при травме спинного мозга - Регенерация нервов - Глава 15 . стр. 237–244. дои : 10.1016/B978-0-12-801732-6.00015-X . ISBN 9780128017326 .
- ^ Абдулхалим М., Фелембан Атари; Сун, Сяохун; Кавано, Рие; Уэзоно, Наохиро; Ито, Аяко; Ахмед, Гиасуддин; Хоссейн, Махмуд; Накашима, Киничи; Танака, Хидеаки (01 мая 2015 г.). «Ахирин регулирует пролиферацию и дифференцировку нервных стволовых клеток в интактном и поврежденном спинном мозге мыши». Развивающая нейробиология . 75 (5): 494–504. дои : 10.1002/dneu.22238 . ISSN 1932-846X . ПМИД 25331329 .