нейруляция
| нейруляция | |
|---|---|
 Поперечные срезы, показывающие продвижение нервной пластинки к нервной борозде снизу вверх. | |
| Идентификаторы | |
| МеШ | D054261 |
| Анатомическая терминология | |
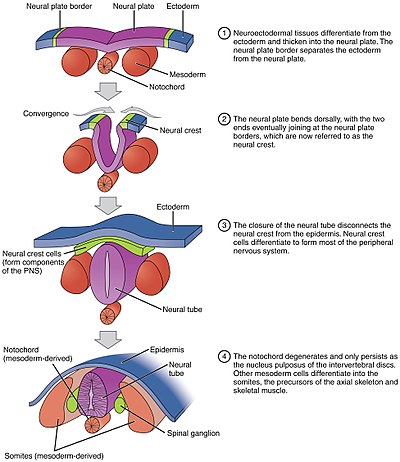
Нейруляция относится к процессу складывания у позвоночных эмбрионов , который включает трансформацию нервной пластинки в нервную трубку . [1] Эмбрион на этой стадии называется нейрулой .
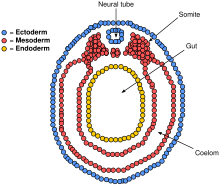
Процесс начинается, когда хорда индуцирует формирование центральной нервной системы (ЦНС), посылая сигнал зародышевому слою эктодермы над ней о формировании толстой и плоской нервной пластинки . Нервная пластинка складывается сама по себе, образуя нервную трубку , которая позже дифференцируется в спинной и головной мозг , образуя в конечном итоге центральную нервную систему. [2] Компьютерное моделирование показало, что заклинивание клеток и дифференциальная пролиферация достаточны для нейруляции млекопитающих. [3]
Различные части нервной трубки у разных видов формируются в результате двух разных процессов, называемых первичной и вторичной нейруляцией. [4]
- При первичной нейруляции нервная пластинка сгибается внутрь до тех пор, пока ее края не соприкоснутся и не срастутся.
- При вторичной нейруляции трубка образуется путем выдалбливания внутренней части твердого предшественника.
Первичная нейруляция [ править ]
нейронная Первичная индукция
Концепция индукции возникла в работе Пандора в 1817 году. [5] Первые эксперименты, подтверждающие индукцию, были приписаны Виктору Гамбургеру. [6] независимым открытиям Ганса Шпемана из Германии в 1901 году. [7] и Уоррен Льюис из США в 1904 году. [8] Ганс Спеманн первым популяризировал термин «первичная нейрональная индукция» в отношении первой дифференцировки эктодермы в нервную ткань во время нейруляции. [9] [10] Его назвали «первичным», потому что считалось, что это первое событие индукции в эмбриогенезе. Эксперимент, получивший Нобелевскую премию, провела его ученица Хильда Мангольд . [9] Эктодерму из области дорсальной губы бластопора развивающегося эмбриона саламандры трансплантировали в другой эмбрион, и эта ткань-организатор «индуцировала» формирование полноценной вторичной оси, меняющей окружающую ткань в исходном эмбрионе с эктодермальной на нервную ткань. Поэтому ткань донорского эмбриона называли индуктором, поскольку она вызывала изменения. [9] Важно отметить, что хотя организатором является дорсальная губа бластопора, это не один набор клеток, а скорее постоянно меняющаяся группа клеток, которые мигрируют по дорсальной губе бластопора, образуя апикально суженные бутылочные клетки. В любой момент времени во время гаструляции органайзер образуют разные клетки. [11]
Последующие работы ученых над индукторами в течение 20-го века показали, что не только спинная губа бластопора может действовать как индуктор, но и огромное количество других, казалось бы, не связанных друг с другом элементов. обнаружил, что вареная эктодерма все еще способна индуцировать Это началось, когда Йоханнес Холтфретер . [12] Такие разнообразные факторы, как низкий уровень pH, циклический АМФ и даже напольная пыль, могут действовать как индукторы, приводящие к серьезному ужасу. [13] Даже ткань, которая не могла активироваться при жизни, могла активироваться при кипячении. [14] Другие предметы, такие как сало, воск, банановая кожура и свернувшаяся лягушачья кровь, не вызывали эффекта. [15] Охотой за химически обоснованной молекулой-индуктором занялись молекулярные биологи, занимающиеся вопросами развития, и обширная литература, посвященная объектам, обладающим индукторными способностями, продолжала расти. [16] [17] Совсем недавно молекулу-индуктор стали приписывать генам, а в 1995 году прозвучал призыв к каталогизации всех генов, участвующих в первичной нейронной индукции, и всех их взаимодействий, чтобы определить «молекулярную природу организатора Спемана». [18] Некоторые другие белки и факторы роста также использовались в качестве индукторов, включая растворимые факторы роста , такие как костный морфогенетический белок , а также потребность в «ингибирующих сигналах», таких как ноггин и фоллистатин .
Еще до того, как термин «индукция» стал популяризирован, несколько авторов, начиная с Ганса Дриша в 1894 г., [19] предположил, что первичная нейронная индукция может иметь механическую природу. Механохимическая модель первичной нейронной индукции была предложена в 1985 году Г. В. Бродландом и Р. Гордоном . [20] Было показано, что настоящая физическая волна сокращения возникает в точном месте расположения организатора Спемана, который затем пересекает предполагаемый нервный эпителий. [21] и полная рабочая модель того, как первичная нейронная индукция была предложена в 2006 году. [22] [23] В этой области долгое время существовало общее нежелание рассматривать возможность того, что первичная нейронная индукция может быть инициирована механическими эффектами. [24] Полное объяснение первичной нейронной индукции еще предстоит найти.
Изменение формы [ править ]
Поскольку нейруляция продолжается после индукции, клетки нервной пластинки становятся высокостолбчатыми и могут быть идентифицированы с помощью микроскопии как отличающиеся от окружающей презумптивной эпителиальной эктодермы ( эпибластической энтодермы у амниот). Клетки движутся латерально и в сторону от центральной оси и принимают форму усеченной пирамиды. Эта форма пирамиды достигается за счет тубулина и актина в апикальной части клетки, которые сжимаются при движении. Изменение формы клеток частично определяется расположением ядра внутри клетки, что приводит к выпучиванию участков клетки, вызывая изменение высоты и формы клетки. Этот процесс известен как апикальное сужение . [25] [26] В результате происходит уплощение дифференцирующейся нервной пластинки, что особенно заметно у саламандр, когда ранее круглая гаструла становится округлым шаром с плоской вершиной. [27] См. Нервная пластинка .
Складной [ править ]
Процесс сворачивания плоской нервной пластинки в цилиндрическую нервную трубку называется первичной нейруляцией . В результате изменения формы клеток нервная пластинка образует медиальную шарнирную точку (МПТ). Расширяющийся эпидермис оказывает давление на MHP и заставляет нервную пластинку сгибаться, что приводит к образованию нервных складок и образованию нервной борозды . Нервные складки образуют дорсолатеральные шарнирные точки (DLHP), и давление на этот шарнир заставляет нервные складки встречаться и сливаться по средней линии. Слияние требует регуляции молекул клеточной адгезии. Нервная пластинка переключается с экспрессии E-кадгерина на экспрессию N-кадгерина и N-CAM, чтобы распознать друг друга как одну и ту же ткань и закрыть трубку. Это изменение экспрессии останавливает связывание нервной трубки с эпидермисом.
Хорда играет важную роль в развитии нервной трубки. До нейруляции, во время миграции клеток эпибластической энтодермы к гипобластической энтодерме, нотохордальный отросток открывается в дугу, называемую нотохордальной пластинкой , и прикрепляет вышележащий нейроэпителий нервной пластинки. Затем нотохордальная пластинка служит якорем для нервной пластинки и толкает два края пластинки вверх, сохраняя при этом среднюю часть закрепленной. Некоторые из нотоходральных клеток включаются в центральную часть нервной пластинки, чтобы позже сформировать пластинку дна нервной трубки. Пластинка хорды отделяется и образует твердую хорду. [4]
Складывание нервной трубки в настоящую трубку не происходит сразу. Вместо этого он начинается примерно на уровне четвертого сомита на 9-й стадии Карнеги (около 20-го дня эмбрионального развития у человека ). Латеральные края нервной пластинки соприкасаются по средней линии и соединяются. Это продолжается как краниально (по направлению к голове), так и каудально (по направлению к хвосту). Отверстия, образующиеся в краниальной и каудальной областях, называются краниальными и каудальными нейропорами . У человека эмбрионов краниальный нейропор закрывается примерно на 24-й день, а каудальный — на 28-й день. [28] Неспособность краниального (верхнего) и каудального (нижнего) закрытия нейропор приводит к состояниям, называемым анэнцефалией и расщелиной позвоночника соответственно. Кроме того, нарушение закрытия нервной трубки по всей длине тела приводит к состоянию, называемому рахишизисом . [29]
Узоры [ править ]
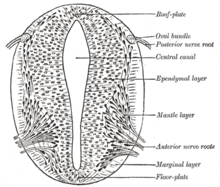
Согласно модели French Flag, где стадии развития управляются градиентами продуктов генов, некоторые гены считаются важными для индукции паттернов в открытой нервной пластинке, особенно для развития нейрогенных плакод . Эти плакоды впервые обнаруживаются гистологически в открытой нервной пластинке. После того, как передача сигнала sonic hedgehog (SHH) из хорды индуцирует его формирование, пластинка дна зарождающейся нервной трубки также секретирует SHH. После закрытия нервная трубка образует базальную или нижнюю пластинку и крышу или крыловую пластинку в ответ на комбинированные эффекты SHH и факторов, включая BMP4, секретируемых верхней пластинкой. Базальная пластинка образует большую часть вентральной части нервной системы, включая двигательную часть спинного мозга и ствола головного мозга; Крылья пластинка образует дорсальную часть, предназначенную в основном для сенсорной обработки. [30]
Дорсальный эпидермис экспрессирует BMP4 и BMP7 . Верхняя пластинка нервной трубки реагирует на эти сигналы, экспрессируя больше сигналов BMP4 и других трансформирующих факторов роста бета (TGF-β), образуя дорсальный/вентральный градиент среди нервной трубки. Хорда выражает SHH. Пластина пола реагирует на SHH, производя собственный SHH и образуя градиент. Эти градиенты позволяют дифференциальную экспрессию факторов транскрипции. [30]
Сложности модели [ править ]
Закрытие нервной трубки до конца не изучено. Закрытие нервной трубки зависит от вида. У млекопитающих закрытие происходит путем встречи в нескольких точках, которые затем закрываются вверх и вниз. У птиц закрытие нервной трубки начинается в одной точке среднего мозга и продвигается вперед и назад. [31] [32]
Вторичная нейруляция [ править ]
Первичная нейруляция развивается во вторичную нейруляцию, когда каудальный нейропор подвергается окончательному закрытию. Полость спинного мозга переходит в нервный мозг. [33] При вторичной нейруляции нейральная эктодерма и некоторые клетки энтодермы образуют мозговой тяж . Медуллярный тяж уплотняется, разделяется и затем образует полости. [34] Затем эти полости сливаются, образуя единую трубку. Вторичная нейруляция встречается в заднем отделе большинства животных, но лучше выражена у птиц. Трубочки как первичной, так и вторичной нейруляции в конечном итоге соединяются примерно на шестой неделе развития. [35]
У человека механизмы вторичной нейруляции играют важную роль, поскольку они влияют на правильное формирование заднего отдела спинного мозга человека. Ошибки на любом этапе процесса могут привести к проблемам. Например, сохранение медуллярного тяжа происходит из-за частичной или полной остановки вторичной нейруляции, в результате чего на рудиментарном конце образуется нефункциональная часть. [36]
Раннее развитие мозга [ править ]
Передняя часть нервной трубки образует три основные части головного мозга: передний мозг ( prosencephalon ), средний мозг ( mesencephalon ) и задний мозг ( ромбенцефалон ). [37] Эти структуры первоначально появляются сразу после закрытия нервной трубки в виде выпуклостей, называемых мозговыми пузырьками , по образцу, определяемому генами передне-заднего формирования паттерна, включая гены Hox , другими факторами транскрипции, такими как гены Emx, Otx и Pax, а также секретируемыми сигнальными факторами, такими как рост фибробластов. факторы (FGFs) и Wnts . [38] Эти мозговые пузырьки далее делятся на субрегионы. Прозэнцефалон дает начало телэнцефалону и промежуточному мозгу , а ромбэнцефалон порождает метэнцефалон и промежуточный мозг . Задний мозг, который является эволюционно самой древней частью мозга хордовых , также делится на различные сегменты, называемые ромбомерами . Ромбомеры генерируют множество наиболее важных нервных цепей, необходимых для жизни, в том числе те, которые контролируют дыхание и частоту сердечных сокращений, а также производят большую часть черепных нервов . [37] Клетки нервного гребня образуют ганглии над каждым ромбомером. Ранняя нервная трубка в основном состоит из зародышевого нейроэпителия , позже названного желудочковой зоной , который содержит первичные нервные стволовые клетки, называемые радиальными глиальными клетками , и служит основным источником нейронов , образующихся во время развития мозга в процессе нейрогенеза . [39] [40]
Ненейральная эктодермы ткань
Параксиальная мезодерма, окружающая хорду по бокам, разовьется в сомиты (будущие мышцы, кости и способствует формированию конечностей позвоночных ) . [41]
Клетки нервного гребня [ править ]
Массы ткани, называемые нервным гребнем , которые расположены на самых краях боковых пластинок складчатой нервной трубки, отделяются от нервной трубки и мигрируют, образуя множество различных, но важных клеток. [ нужна ссылка ]
Клетки нервного гребня будут мигрировать через эмбрион и давать начало нескольким популяциям клеток, включая пигментные клетки и клетки периферической нервной системы. [ нужна ссылка ]
Дефекты нервной трубки [ править ]
Нарушение нейруляции, особенно нарушение закрытия нервной трубки, являются одними из наиболее распространенных и инвалидизирующих врожденных дефектов у человека, встречаясь примерно у 1 из каждых 500 живорождений. [42] Неспособность рострального конца нервной трубки закрыться приводит к анэнцефалии или отсутствию развития мозга и чаще всего приводит к летальному исходу. [43] Неспособность каудального конца нервной трубки закрыться вызывает состояние, известное как расщелина позвоночника , при котором спинной мозг не закрывается. [44]
См. также [ править ]
Ссылки [ править ]
- ^ Ларсен В.Дж. Эмбриология человека. Третье изд. 2001. С 86. ISBN 0-443-06583-7
- ^ «Глава 14. Гаструляция и нейруляция» . biology.kenyon.edu . Проверено 2 февраля 2016 г.
- ^ Нильсен, Бьярке Фрост; Ниссен, Сайлас Бойе; Снеппен, Ким; Матисен, Иоахим; Трусина Аля (21 февраля 2020 г.). «Модель, связывающая форму и полярность клеток с органогенезом» . iScience . 23 (2): 100830. Бибкод : 2020iSci...23j0830N . дои : 10.1016/j.isci.2020.100830 . ПМК 6994644 . ПМИД 31986479 .
- ↑ Перейти обратно: Перейти обратно: а б Вулперт, Льюис; Пощекочи, Шерил; Ариас, Альфонсо Мартинес (2015). Принципы развития (Пятое изд.). Оксфорд, Соединенное Королевство: Издательство Оксфордского университета. п. 393. ИСБН 9780198709886 .
- ^ Тидеманн, Х. Химический подход к индуцирующим агентам. В: О. Накамура и С. Тойвонен (ред.), Организатор – веха полувека от Спеманна, Амстердам: Elsevier/North Holland Biomedical Press, стр. 91- 117. 1978 г.
- ^ Гамбургер, В.. Наследие экспериментальной эмбриологии: Ганс Спеманн и организатор. Нью-Йорк: Издательство Оксфордского университета. 1988 год
- ^ Спеманн, Х. О корреляциях в развитии глаза. Брак анат. Йена 15, 61-79. 1901 год
- ^ Льюис, WH Экспериментальные исследования развития глаза у амфибий. I. О происхождении хрусталика у Rana palustris. амер. Дж. Анат. 3, 505–536. 1904 год
- ↑ Перейти обратно: Перейти обратно: а б с Спеманн, Х. и Х. Мангольд, Об индукции зачатка эмбриона путем имплантации организаторов других видов. Архивный микроскоп. Анат. 100, 599–638 1924 г.
- ^ Спеманн, Х. и Х. Мангольд 1924: Индукция эмбриональных зачатков путем имплантации организаторов от другого вида. В: BH Willier & JM Oppenheimer (ред.), Foundations of Experimental Embryology (переведено изд. 1964 г.), Englewood Cliffs, New Jersey: Prentice-Hall, p. 144-184
- ^ Гордон, Р., Н.К. Бьорклунд и П.Д. Ньюкооп. Диалог об эмбриональных волнах индукции и дифференцировки. Межд. Преподобный Цитол. 150, 373–420. 1994 г.
- ^ Холтфретер, Дж. Характеристики и распространение индуцирующих веществ. Естественные науки 21, 766–770. 1933 год
- ^ Твитти, ВК, Об ученых и саламандрах Фриман, Сан-Франциско, Калифорния, 1966 г.
- ^ Спеманн, Х., Ф.Г. Фишер и Э. Вемайер. Продолжающиеся попытки анализа причин индукции в эмбриональном развитии. Естественные науки 21, 505–506. 1933 год
- ^ Вайс, Пенсильвания. Так называемый организатор и проблема организации развития земноводных. Физиол. Откр. 15(4), 639-674. 1935 год
- ^ Де Робертис, Э.М., М. Блюм, К. Ниерс и Х. Штайнбейссер, гусекоид и организатор. Развитие (Приложение), 167-171. 1992 год
- ^ Хан, М. и Х. Джекль Гусекоид дрозофилы участвует в развитии нервной системы, но не в формировании оси тела. ЭМБО Дж. 15(12), 3077-3084. 1996 год
- ^ Де Робертис, Э.М. Демонтаж органайзера. Природа 374(6521), 407-408. 1995 год
- ^ Дриш, ХАЕ. Аналитическая теория органического развития. Лейпциг: Издательство Вильгельма Энгельмана. 1984 г.
- ^ Гордон, Р. Бродланд, GW. Цитоскелетная механика морфогенеза головного мозга: расщепители состояний клеток вызывают первичную нервную индукцию. Гелл Биофиз. 11: 177-238. (1987)
- ^ Бродланд, Г.В.» Гордон, Р., Скотт М.Дж., Бьорклунд Н.К., Лучка К.Б., Мартин, К.С., Матуга, К., Глобус, М., Ветамани-Глобус С. и Шу, Д. Волна сокращения поверхности бороздок, совпадающая с первичной нейронной волной индукция у эмбрионов амфибий. Дж Морфол. 219: 131-142. 1994 г.
- ^ Гордон, Н.К., Гордон Р. Органелла дифференцировки эмбрионов: расщепитель клеточного состояния Theor Biol Med Model (2016) 13: 11. https://doi.org/10.1186/s12976-016-0037-2
- ^ Бьёрклунд, Н.К., Гордон, Р.А. Гипотеза, связывающая низкое потребление фолиевой кислоты с дефектами нервной трубки из-за нарушения посттрансляционного метилирования цитоскелета. Международный журнал биологии развития 50 (2-3), 135-141.
- ^ Иерархический геном и волны дифференциации . Серия по математической биологии и медицине. Том. 3. Всемирная научная издательская компания. 1999. дои : 10.1142/2755 . ISBN 978-981-02-2268-0 .
- ^ Бернсайд. М.Б. Микрорубулы и микрофиламенты в нейруляции амфибий. Алий. Зоол. 13, 989-1006 1973 г.
- ^ Джейкобсон, АГ и Р. Гордон. Изменения вформа развивающейся нервной системы позвоночных анализируется экспериментально, математически и с помощью компьютерного моделирования. Дж. Эксп. Зоол. 197, 191–246. 1973 год
- ^ Бордзиловская, Н.П., Т.А. Детлафф, С.Т. Духон и Г.М. Малачински (1989). Серия эмбрионов аксолотля на стадии развития [Ошибка: Таблица стадий 19-1 предназначена для 20°C, а не 29°C]. В: Дж. Б. Армстронг и Г. М. Малачински (ред.), Биология развития аксолотля, Нью-Йорк: Oxford University Press, стр. 201-219.
- ^ Неврологическая хирургия Юмана, Х. Ричард Винн, 6-е изд. Том 1, стр. 81, 2011 г. Эльзевир Сондерс, Филадельфия, Пенсильвания.
- ^ Гилберт, Сан-Франциско (2000). «12: Формирование нервной трубки» . Биология развития (6-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-243-6 . Проверено 30 ноября 2011 г.
- ↑ Перейти обратно: Перейти обратно: а б Гилберт, Сан-Франциско (2013). «10: Возникновение эктодермы» . Биология развития (10-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-978-7 . Проверено 22 марта 2015 г.
- ^ Голден Дж. А., Чернов Г. Ф. Прерывистый характер закрытия нервной трубки у двух линий мышей. Тератология. 1993;47:73–80.
- ^ Ван Аллен М.И., 15 других. Доказательства множественного закрытия нервной трубки у человека. Являюсь. Дж. Мед. Жене. 1993;47:723–743.
- ^ Шепард, Томас Х. (1989). «Стадии развития человеческих эмбрионов. Р. О'Рахилли и Ф. Мюллер (редакторы), Институт Карнеги в Вашингтоне, Вашингтон, округ Колумбия, 1987, 306 стр., 52 доллара» . Тератология . 40:85 . дои : 10.1002/тера.1420400111 .
- ^ Формирование книжной полки NCBI по биологии развития нервной трубки
- ^ Симокита, Э; Такахаши, Ю. (апрель 2011 г.). «Вторичная нейруляция: картирование судьбы и манипуляция генами нервной трубки хвостовой зачатки» . Развитие, рост и дифференциация . 53 (3): 401–10. дои : 10.1111/j.1440-169X.2011.01260.x . ПМИД 21492152 .
- ^ Панг, Дахлинг; Зовикян, Джон (2011). " "Сохраненный мозговой канатик у человека: поздняя остановка вторичной нейруляции" " . Нейрохирургия . 68 (6): 1500–19. дои : 10.1227/NEU.0b013e31820ee282 . ПМИД 21336222 . S2CID 25638763 . Проверено 19 ноября 2020 г.
- ↑ Перейти обратно: Перейти обратно: а б Гилберт, Скотт Ф.; Колледж, Суортмор; Хельсинкский университет (2014). Биология развития (Десятое изд.). Сандерленд, Массачусетс: Синауэр. ISBN 978-0878939787 .
- ^ Эрик Р. Кандел, изд. (2006). Принципы нейронауки (5-е изд.). Эпплтон и Ланге: МакГроу Хилл. ISBN 978-0071390118 .
- ^ Ракич, П. (октябрь 2009 г.). «Эволюция неокортекса: взгляд из биологии развития» . Обзоры природы. Нейронаука . 10 (10): 724–35. дои : 10.1038/nrn2719 . ПМЦ 2913577 . ПМИД 19763105 .
- ^ Деэй, К; Кеннеди, Х. (июнь 2007 г.). «Контроль клеточного цикла и развитие коры». Обзоры природы. Нейронаука . 8 (6): 438–50. дои : 10.1038/nrn2097 . ПМИД 17514197 . S2CID 1851646 .
- ^ Параксиальная мезодерма: сомиты и их производные. Книжная полка NCBI, Биология развития, 6-е издание. По состоянию на 29 ноября 2017 г.
- ^ Дейли, Даррел. Формирование нервной системы. Архивировано 3 января 2008 г. в Wayback Machine . Последний доступ: 29 октября 2007 г.
- ^ Справочник, Дом генетики. «Анэнцефалия» . Домашний справочник по генетике . Проверено 02 марта 2020 г.
- ^ Центр по контролю и профилактике заболеваний (31 августа 2018 г.). «Факты о расщелине позвоночника | CDC» . Центры по контролю и профилактике заболеваний . Проверено 02 марта 2020 г.
Дальнейшее чтение [ править ]
- Алмейда, Карла Л.; и др. (2010). «Нейронная индукция» . В Хеннинге, Ульрихе (ред.). Перспективы стволовых клеток: от инструментов изучения механизмов дифференцировки нейронов к терапии . Спрингер. ISBN 978-90-481-3374-1 .
- Баш, Мартин Л.; Боннер-Фрейзер, Марианна (2006). «Сигналы, индуцирующие нервный гребень» . В Сен-Женне, Жан-Пьер (ред.). Индукция и дифференцировка нервного гребня . Спрингер. ISBN 978-0-387-35136-0 .
- Харланд, Ричард М. (1997). «Нейронная индукция у Xenopus » . В Коуэне, У. Максвелл (ред.). Молекулярный и клеточный подходы к развитию нейронов . Издательство Оксфордского университета. ISBN 978-0-19-511166-8 .
- Ладер, Радж; Шенвольф, Гэри К. (2004). «Изготовление нервной трубки» . В Джейкобсоне, Маркус; Рао, Махендра С. (ред.). Нейробиология развития . Спрингер. ISBN 978-0-306-48330-1 .
- Тянь, Цзин; Сампат, Каруна (2004). «Формирование и функции пластинки пола». В Гонге, Чжиюань; Корж, Владимир (ред.). Развитие и генетика рыб: модели данио и медака . Всемирная научная. стр. 123 , 139–140 . ISBN 978-981-238-821-6 .
- Чжан, Су-Чун (2005). «Нейральная спецификация эмбриональных стволовых клеток человека» . В Одорико, Джон С.; и др. (ред.). Эмбриональные стволовые клетки человека . Гирляндная наука. ISBN 978-1-85996-278-7 .
Внешние ссылки [ править ]
- Обзор на uvm.edu
- Анимация нейруляции, заархивировано 3 марта 2016 г. в Wayback Machine.