Бластоцель
| Бластоцель | |
|---|---|
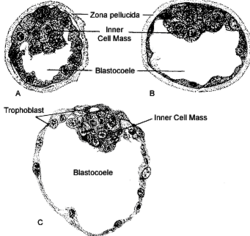 Бластоцель млекопитающих | |
 Схематическая диаграмма, показывающая бластоцисту с ее эмбриобластом (внутренней клеточной массой) и слоем трофобласта , расположенными рядом с поверхностью эндометрия . | |
| Подробности | |
| Этап Карнеги | 3 |
| Дни | 5 |
| Предшественник | Морула |
| Дает начало | Гаструла , примитивный желточный мешок |
| Анатомическая терминология | |
Бластоцель s ( / ˈ b l æ s t ə ˌ iː l бластоцеле / ), также пишется . и бластоцеле , а также называется полостью расщепления или полостью сегментации [1] представляет собой полость, заполненную жидкостью или желтком, которая образуется в бластуле на очень ранних стадиях эмбрионального развития . На этой стадии у млекопитающих бластула называется бластоцистой и состоит из наружного эпителия — трофэктодермы , окутывающего внутреннюю клеточную массу и бластоцель .
Он развивается дробления зиготы оплодотворения после . после [2] [3] Это первая заполненная жидкостью полость или просвет, образующаяся по мере увеличения эмбриона. [4] и является важным предшественником дифференцированной гаструлы . [5] [ нужна страница ] У Xenopus на двухклеточной стадии развития описана очень маленькая полость. [6]
У млекопитающих
[ редактировать ]После оплодотворения зигота претерпевает несколько циклов деления , образуя дочерние клетки, известные как бластомеры . На стадии 8 или 16 клеток зародыш уплотняется и образует морулу . В конце концов, морула представляет собой сплошной шар клеток, в котором есть небольшая группа внутренних клеток, окруженная большей группой внешних клеток. Затем бластомеры подвергаются клеточной дифференцировке , при этом внутренние клетки принимают судьбу внутренней клеточной массы , а внешний слой становится трофэктодермой . Внутренняя клеточная масса впоследствии станет настоящим эмбрионом. Внешние, окружающие клетки развиваются в клетки трофобласта , которые вносят вклад только во внеэмбриональные ткани. нет просвета На этой стадии внутри эмбриона . В процессе, называемом кавитацией , клетки трофэктодермы транспортируют жидкость в эмбрион, создавая бластоцель, заполненный жидкостью просвет . Мембраны клеток трофэктодермы содержат натрий (Na + ) насосы, Na + /К + - АТФаза и Na + /ЧАС + обменники, которые перекачивают натрий в эмбрион. Клетки яйцевода стимулируют натриевые насосы трофобласта, когда оплодотворенная яйцеклетка перемещается по фаллопиевой трубе к матке. [7] Накопление натрия вытягивает воду посредством осмоса . [2] Накопление воды разрывает межклеточные контакты посредством гидроразрыва . [8] Чтобы сформировать единый просвет, жидкость из нескольких водных карманов собирается в единое целое, что похоже на созревание Оствальда . [9] Бластоцель далее расширяется, и внутренняя клеточная масса оказывается на одной стороне клеток трофобласта, образуя бластулу млекопитающих, называемую бластоцистой . Ось, образованная внутренней клеточной массой и бластоцелем, является первой осью симметрии эмбриона млекопитающих и определяет точку его прикрепления к матке.
У амфибий
[ редактировать ]Эмбрион амфибии на стадии 128 клеток считается бластулой, поскольку на этой стадии у эмбриона становится очевидным бластоцель. образуется полость, заполненная жидкостью В анималистическом полушарии лягушки . Однако раннее формирование бластоцеля было прослежено до самой первой борозды дробления . На эмбрионе лягушки было показано, что первая борозда дробления расширяется в полушарии животного, образуя небольшую межклеточную полость, изолированную плотными соединениями. [3] По мере продолжения дробления полость расширяется и превращается в развитый бластоцель. Бластоцель является важнейшим компонентом развития эмбриона амфибий. Он обеспечивает миграцию клеток во время гаструляции и предотвращает преждевременное взаимодействие клеток под бластоцелем с клетками над бластоцелем. Например, бластоцель предотвращает вегетативных клеток, которым суждено стать энтодермой контакт , с теми клетками эктодермы, которым суждено дать начало коже и нервам. [10]
Повреждение бластоцеля
[ редактировать ]Бластоцель может быть поврежден и уничтожен, если адгезия между бластомерами, обеспечиваемая молекулами клеточной адгезии, такими как EP-кадгерин, разрушается в виде мРНК олигонуклеотидами . Если мРНК разрушена, EP-кадгерина нет, адгезия бластомеров практически отсутствует, а бластоцель отсутствует. [4] На следующей стадии эмбрионального развития, гаструляции амфибий , бластоцель смещается за счет образования архентерона в середине гаструляции. В конце гаструляции бластоцель уничтожается. [11]
У морских ежей
[ редактировать ]На стадии 120 клеток эмбрион морского ежа считается бластулой из-за развитого бластоцеля, который окружает и соприкасается каждая эмбриональная клетка. Каждая клетка изнутри контактирует с белковой жидкостью бластоцеля, а снаружи соприкасается с гиалиновым слоем. Слабо связанные бластомеры теперь плотно соединены благодаря плотным соединениям , которые создают бесшовный эпителий, полностью окружающий бластоцель. [12] Даже когда бластомеры продолжают делиться, бластула остается толщиной в одну клетку и истончается по мере расширения эмбриона наружу. Частично это достигается за счет притока воды, которая расширяет бластоцель и выталкивает окружающие его клетки наружу. К этому моменту клетки конкретизируются и покрываются реснитками на противоположной стороне бластоцеля. Вегетативная пластинка и животное полушарие развивают и выделяют фермент вылупления, который переваривает оболочку оплодотворения и позволяет эмбриону теперь стать свободно плавающей вылупившейся бластулой. [13]
Развитие первичной мезенхимы
[ редактировать ]Для бластулы морского ежа важным является проникновение первичной мезенхимы . После того, как бластула вылупляется из оболочки оплодотворения, вегетативная сторона бластулы начинает уплощаться и утолщаться, поскольку в небольшом скоплении этих клеток развиваются длинные тонкие отростки, называемые филоподиями . Эти клетки затем диссоциируют и проникают в бластоцель и называются первичной мезенхимой. Клетки хаотично перемещаются внутри бластоцеля, пока не локализуются в вентролатеральной области бластоцеля. [13] [12]
У птиц
[ редактировать ]Как и у млекопитающих, оплодотворение яйцеклетки птиц происходит в яйцеводе. Оттуда бластодиск , небольшое скопление клеток в анимальном полюсе яйца, затем подвергается дискоидальному меробластическому расщеплению. Бластодерма развивается в эпибласт и гипобласт, и между этими слоями формируется бластоцель. Форма и формирование бластодиска птиц отличаются от бластул земноводных, рыб и иглокожих, но общее пространственное соотношение бластоцеля остается тем же. [5] [ нужна страница ]
Формирование примитивной полоски
[ редактировать ]Птичий бластоцель важен во время развития примитивной полоски. Ингрессия энтодермальных клеток-предшественников формирует эпибласт в бластоцель, а миграция латеральных клеток заднего эпибласта к центру образует раннюю примитивную полоску . Когда эти клетки сходятся внутрь, образуется углубление, называемое примитивной бороздкой , которое функционирует как отверстие, через которое клетки попадают в бластоцель. По мере миграции клеток в бластоцель они подвергаются эпителиально-мезенхимальной трансформации . [5] [ нужна страница ]
У рыбок данио
[ редактировать ]В отличие от эмбрионов амфибий, иглокожих, млекопитающих и птиц, у рыбок данио нет четко выраженного бластоцеля. Скорее, у них есть небольшие внеклеточные пространства неправильной формы, которые образуются между клетками бластодиска, расположенными над желтком. [14]
Ссылки
[ редактировать ]- ^ «Определение БЛАСТОЦЕЛЬ» . Мерриам-Вебстер .
- ^ Jump up to: а б Биггерс, Джей Ди; Борланд, РМ; Пауэрс, Р.Д. (1977). «Механизмы транспорта в преимплантационном эмбрионе млекопитающих». Симпозиум 52 Фонда Ciba – Замораживание эмбрионов млекопитающих . Симпозиумы Фонда Новартис. Том. 52. С. 129–53. дои : 10.1002/9780470720332.ch7 . ISBN 9780470720332 . ПМИД 145938 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Jump up to: а б Кальт, Марвин Р. (1971). «Взаимосвязь между дроблением и образованием бластоцеля у Xenopus laevis. I. Световые микроскопические наблюдения» . Журнал эмбриологии и экспериментальной морфологии . 26 (1): 37–49. ПМИД 5565077 .
- ^ Jump up to: а б Хисман, Джанет; Кроуфорд, Аарон; Голдстоун, Ким; Гарнер-Хэмрик, Пегги; Гамбинер, Барри; МакКри, Пьер; Кинтнер, Крис; Норо, Чикако Ёсида; Уайли, Крис (1994). «Сверхэкспрессия кадгеринов и недостаточная экспрессия β-катенина ингибируют индукцию дорсальной мезодермы у ранних эмбрионов Xenopus». Клетка . 79 (5): 791–803. дои : 10.1016/0092-8674(94)90069-8 . ПМИД 7528101 . S2CID 33403560 .
- ^ Jump up to: а б с Гилберт, Скотт Ф. (2010). Биология развития (9-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-384-6 .
- ^ Вулперт, Льюис (2015). Принципы развития (Пятое изд.). Оксфорд, Великобритания. п. 375. ИСБН 9780199678143 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Уайли, Линн М. (1984). «Кавитация в предимплантационном эмбрионе мыши: и происхождение зарождающейся жидкости бластоцеле». Биология развития . 105 (2): 330–42. дои : 10.1016/0012-1606(84)90290-2 . ПМИД 6090240 .
- ^ Дюмортье, Жюльен Г.; Ле Верж-Серандур, Матье; Торторелли, Анна Франческа; Мильке, Аннетт; Де Платер, Людмила; Тюрлье, Эрве; Мэтр, Жан-Леон (2019). «Гидравлический разрыв и активное огрубление позиционируют просвет бластоцисты мыши» . Наука . 365 (6452): 465–468. дои : 10.1126/science.aaw7709 .
- ^ Дюмортье, Жюльен Г.; Ле Верж-Серандур, Матье; Торторелли, Анна Франческа; Мильке, Аннетт; Де Платер, Людмила; Тюрлье, Эрве; Мэтр, Жан-Леон (2019). «Гидравлический разрыв и активное огрубление позиционируют просвет бластоцисты мыши» . Наука . 365 (6452): 465–468. дои : 10.1126/science.aaw7709 .
- ^ Ньюкооп, П.Д. (1973). «Организационный центр зародыша амфибии: его происхождение, пространственная организация и морфогенетическое действие». Достижения в области морфогенеза . 10 :1–39. дои : 10.1016/b978-0-12-028610-2.50005-8 . ISBN 9780120286102 . ПМИД 4581327 .
- ^ Перселл, С.М.; Келлер, Р. (январь 1993 г.). «Другой тип морфогенеза мезодермы амфибий у Ceratophrys ornata». Разработка . 117 (1): 307–17. дои : 10.1242/dev.117.1.307 . ПМИД 8223254 .
- ^ Jump up to: а б Галилей, Дени С.; Моррилл, Джон Б. (1985). «Схемы клеток и внеклеточного материала эмбриона морского ежа Lytechinus variegatus (Echinodermata; Echinoidea), от вылупившейся бластулы до поздней гаструлы». Журнал морфологии . 185 (3): 387–402. дои : 10.1002/jmor.1051850310 . ПМИД 29991195 . S2CID 51615081 .
- ^ Jump up to: а б Черр, Дж.Н.; Саммерс, Р.Г.; Болдуин, доктор медицинских наук; Моррилл, Дж. Б. (15 июня 1992 г.). «Сохранение и визуализация бластоцельного внеклеточного матрикса эмбриона морского ежа». Микроскопические исследования и техника . 22 (1): 11–22. дои : 10.1002/jemt.1070220104 . ПМИД 1617206 . S2CID 32044141 .
- ^ Киммел, Чарльз Б.; Баллард, Уильям В.; Киммел, Сет Р.; Ульманн, Бонни; Шиллинг, Томас Ф. (1995). «Этапы эмбрионального развития рыбок данио». Динамика развития . 203 (3): 253–310. дои : 10.1002/aja.1002030302 . ПМИД 8589427 . S2CID 19327966 .
Дальнейшее чтение
[ редактировать ]- Персонал Дорландса (2004). «бластоцель [словарная статья]». Иллюстрированный медицинский словарь Дорланда (онлайн). Амстердам, NDE: Эльзевир-Сондерс. Проверено 30 января 2016 г. «бластоцель...[blaso- + -coele] заполненная жидкостью полость массы клеток (бластулы), образующаяся в результате расщепления оплодотворенной яйцеклетки. Иногда пишется...[c]alled...» Также «бластоцелия...относящаяся к бластоцеле».; Дорландс.ком
- Гилберт, Скотт Ф. (2000). «Раннее развитие млекопитающих» . Биология развития (6-е изд.). Сандерленд: Sinauer Associates. ISBN 0-87893-243-7 .
- Гилберт, Скотт Ф. (2000). «Раннее развитие амфибий» . Биология развития (6-е изд.). Сандерленд: Sinauer Associates. ISBN 0-87893-243-7 .
- Гилберт, Скотт Ф. (2000). «Раннее развитие морских ежей» . Биология развития (6-е изд.). Сандерленд: Sinauer Associates. ISBN 0-87893-243-7 .