Взрыв
| Взрыв | |
|---|---|
 Бластуляция: от 1 . морула до 2 . бластула | |
| Подробности | |
| Дни | 4 |
| Предшественник | Морула |
| Дает начало | Гаструла |
| Анатомическая терминология | |
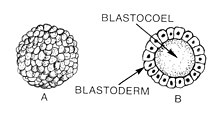
Бластуляция — это стадия раннего животных эмбрионального развития , на которой образуется бластула . В развитии млекопитающих бластула развивается в бластоцисты с дифференцированной внутренней клеточной массой и внешней трофэктодермой . Бластула (от греческого βλαστός ( бластос означает росток )) представляет собой полую сферу клеток, известных как бластомеры, окружающих внутреннюю заполненную жидкостью полость, называемую бластоцель . [1] [2] Эмбриональное развитие начинается с того, что , образуя зиготу сперматозоид оплодотворяет яйцеклетку , которая подвергается многочисленным расщеплениям и превращается в клубок клеток, называемый морулой . Только когда формируется бластоцель, ранний зародыш становится бластулой. Бластула предшествует образованию гаструлы , в которой формируются зародышевые листки зародыша. [3]
Общей особенностью бластулы позвоночных является то, что она состоит из слоя бластомеров, известного как бластодерма , который окружает бластоцель. [4] [5] У млекопитающих бластоциста содержит эмбриобласт (или внутреннюю клеточную массу), который в конечном итоге дает начало дефинитивным структурам плода , и трофобласт , который в дальнейшем формирует внеэмбриональные ткани. [3] [6]
Во время бластуляции внутри раннего эмбриона происходит значительная активность по установлению полярности клеток , спецификации клеток , формированию осей и регуляции экспрессии генов . [7] У многих животных, таких как Drosophila и Xenopus , переход средней бластулы (MBT) является решающим этапом развития, во время которого материнская мРНК деградирует и контроль над развитием передается эмбриону. [8] Многие взаимодействия между бластомерами зависят от экспрессии кадгерина , особенно E-кадгерина у млекопитающих и EP-кадгерина у амфибий . [7]
Изучение бластулы и клеточной спецификации имеет большое значение для исследований стволовых клеток и вспомогательных репродуктивных технологий . [6] У Xenopus бластомеры ведут себя как плюрипотентные стволовые клетки, которые могут мигрировать по нескольким путям, в зависимости от передачи сигналов клетками . [9] Манипулируя клеточными сигналами на стадии развития бластулы, различные ткани можно формировать . Этот потенциал может сыграть важную роль в регенеративной медицине при заболеваниях и травмах. Экстракорпоральное оплодотворение предполагает перенос эмбриона в матку для имплантации . [10]
Разработка
[ редактировать ]Стадия бластулы раннего развития эмбриона начинается с появления бластоцеля. что бластоцель у Xenopus Было показано, берет начало из первой борозды расщепления , которая расширена и запечатана плотными соединениями , образуя полость . [11]
У многих организмов развитие эмбриона до этого момента и на ранней стадии стадии бластулы контролируется материнской мРНК, названной так потому, что она вырабатывается в яйцеклетке до оплодотворения и, следовательно, поступает исключительно от матери. [12] [13]
Переход мидбластулы
[ редактировать ]У многих организмов, включая Xenopus и Drosophila , переход средней бластулы обычно происходит после определенного количества делений клеток для данного вида и определяется окончанием синхронных циклов деления клеток раннего развития бластулы и удлинением клеточных циклов. путем добавления фаз G1 и G2 . До этого перехода расщепление происходит только с фазами синтеза и митоза клеточного цикла. [13] Добавление двух фаз роста в клеточный цикл позволяет клеткам увеличиваться в размерах, поскольку до этого момента бластомеры подвергаются редукционным делениям, при которых общий размер эмбриона не увеличивается, но создается больше клеток. С этого перехода начинается рост организма в размерах. [3]
Переход средней бластулы также характеризуется заметным увеличением транскрипции новой, нематеринской мРНК, транскрибируемой из генома организма. В этот момент большое количество материнской мРНК разрушается либо такими белками, как SMAUG, у дрозофилы. [14] или микроРНК . [15] Эти два процесса смещают контроль над эмбрионом от материнской мРНК к ядрам.
Структура
[ редактировать ]Бластула ( бластоциста у млекопитающих ) представляет собой сферу клеток, окружающую заполненную жидкостью полость, называемую бластоцель . Бластоцель содержит аминокислоты , белки , факторы роста , сахара, ионы и другие компоненты, необходимые для клеточной дифференциации . Бластоцель также позволяет бластомерам двигаться в процессе гаструляции . [16]
У эмбрионов Xenopus бластула состоит из трех разных областей. Животная шапочка образует крышу бластоцеля и в первую очередь образует эктодермальные производные. Экваториальная или маргинальная зона, составляющая стенки бластоцеля, дифференцируется преимущественно в мезодермальную ткань. Вегетативная масса состоит из дна бластоцеля и в первую очередь развивается в энтодермальную ткань. [7]
В бластоцисте млекопитающих есть три линии, которые дают начало более позднему развитию тканей. Эпибласт дает начало самому плоду , тогда как трофобласт развивается в часть плаценты , а примитивная энтодерма становится желточным мешком . [6] У эмбриона мыши формирование бластоцеля начинается на стадии 32 клеток. Во время этого процесса вода поступает в эмбрион благодаря осмотическому градиенту, который является результатом работы натрий-калиевых насосов , которые создают высокий градиент натрия на базолатеральной стороне трофэктодермы. Этому движению воды способствуют аквапорины . Уплотнение создается плотными соединениями эпителиальных клеток , выстилающих бластоцель. [6]
Клеточная адгезия
[ редактировать ]Плотные соединения очень важны для развития эмбриона. В бластуле эти кадгерин-опосредованные клеточные взаимодействия необходимы для развития эпителия, который наиболее важен для параклеточного транспорта , поддержания полярности клеток и создания проницаемого уплотнения для регуляции образования бластоцеля. Эти плотные контакты возникают после того, как устанавливается полярность эпителиальных клеток, что закладывает основу для дальнейшего развития и спецификации. Внутри бластулы внутренние бластомеры обычно неполярны, тогда как эпителиальные клетки демонстрируют полярность. [16]
Эмбрионы млекопитающих подвергаются уплотнению на стадии 8 клеток, где E-кадгерины, а также альфа- и бета -катенины экспрессируются . В результате этого процесса образуется клубок эмбриональных клеток, способных взаимодействовать, а не группа диффузных и недифференцированных клеток. Адгезия E-кадгерина определяет апико-базальную ось в развивающемся эмбрионе и превращает эмбрион из нечеткого клубка клеток в более поляризованный фенотип , который создает основу для дальнейшего развития в полностью сформированную бластоцисту. [16]
Полярность мембраны Xenopus устанавливается при первом расщеплении клетки. EP-кадгерин амфибий и XB/U кадгерин выполняют ту же роль, что и E-кадгерин у млекопитающих, устанавливая полярность бластомеров и укрепляя межклеточные взаимодействия, которые имеют решающее значение для дальнейшего развития. [16]
Клинические последствия
[ редактировать ]Технологии внесения удобрений
[ редактировать ]Эксперименты с имплантацией на мышах показывают, что гормональная индукция , суперовуляция и искусственное оплодотворение успешно создают преимплантационные эмбрионы мышей. У мышей девяносто процентов самок были вынуждены с помощью механической стимуляции забеременеть и имплантировать хотя бы один эмбрион. [17] Эти результаты оказались обнадеживающими, поскольку они создают основу для потенциальной имплантации другим видам млекопитающих, например человеку.
Стволовые клетки
[ редактировать ]Клетки на стадии бластулы у многих видов могут вести себя как плюрипотентные стволовые клетки. Плюрипотентные стволовые клетки являются отправной точкой для производства органоспецифичных клеток, которые потенциально могут помочь в восстановлении и предотвращении повреждений и дегенерации. Сочетание экспрессии факторов транскрипции и локационного позиционирования клеток бластулы может привести к развитию индуцированных функциональных органов и тканей. Плюрипотентные клетки Xenopus при использовании в стратегии in vivo смогли сформировать функциональную сетчатку . Трансплантировав их в поле глаза на нервной пластинке и индуцировав несколько неправильных экспрессий факторов транскрипции, клетки были переданы в линию сетчатки и могли управлять поведением, основанным на зрении, у Xenopus . [18]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Взрыв» . веб-книги.com .
- ^ «Бластула» . Британская энциклопедия . 2013.
- ^ Jump up to: Перейти обратно: а б с Гилберт, Скотт (2010). Биология развития, 9-е изд. + Лаборатория Devbio Vade Mecum3 . Sinauer Associates Inc., стр. 243–247, 161. ISBN. 978-0-87893-558-1 . [ постоянная мертвая ссылка ]
- ^ Ломбарди, Джулиан (1998). «Эмбриогенез» . Сравнительное размножение позвоночных . Спрингер. п. 226. ИСБН 978-0-7923-8336-9 .
- ^ Форгач и Ньюман, 2005: с. 27
- ^ Jump up to: Перейти обратно: а б с д Кокберн, Кэти; Россант, Джанет (1 апреля 2010 г.). «Изготовление бластоцисты: уроки мыши» . Журнал клинических исследований . 120 (4): 995–1003. дои : 10.1172/JCI41229 . ПМК 2846056 . ПМИД 20364097 .
- ^ Jump up to: Перейти обратно: а б с Хисман, Дж. (ноябрь 1997 г.). «Формирование бластулы Xenopus » . Разработка . 124 (21): 4179–91. дои : 10.1242/dev.124.21.4179 . ПМИД 9334267 .
- ^ Тадрос, Ваэль; Липшиц, Ховард Д. (1 марта 2004 г.). «Подготовка почвы для развития: трансляция и стабильность мРНК во время созревания ооцитов и активации яйцеклеток у дрозофилы » . Динамика развития . 232 (3): 593–608. дои : 10.1002/dvdy.20297 . ПМИД 15704150 .
- ^ Гурдон, Джон Б.; Стэндли, Генриетта Дж. (декабрь 2002 г.). «Незафиксированные клетки бластулы Xenopus могут быть направлены на однородную экспрессию мышечных генов путем интерпретации градиента и эффекта сообщества» . Международный журнал биологии развития (Кембридж, Великобритания) . 46 (8): 993–8. ПМИД 12533022 .
- ^ Тот, Аттила. «Лечение: устранение причин бесплодия у мужчин и женщин» . Лаборатория Маклеода . Проверено 22 марта 2013 г.
- ^ Кальт, MR (август 1971 г.). «Взаимосвязь между дроблением и образованием бластоцеля у Xenopus laevis . I. Световые микроскопические наблюдения». Журнал эмбриологии и экспериментальной морфологии . 26 (1): 37–49. ПМИД 5565077 .
- ^ Тадрос, В; Липшиц, HD (март 2005 г.). «Подготовка почвы для развития: трансляция и стабильность мРНК во время созревания ооцитов и активации яйцеклеток у дрозофилы » . Динамика развития . 232 (3): 593–608. дои : 10.1002/dvdy.20297 . ПМИД 15704150 .
- ^ Jump up to: Перейти обратно: а б Эткин Л.Д. (1988) [1985]. «Регуляция перехода средней бластулы у амфибий». В Браудер Л.В. (ред.). Молекулярная биология клеточной детерминации и дифференцировки клеток . Биология развития. Том. 5. Нью-Йорк. стр. 209–25. дои : 10.1007/978-1-4615-6817-9_7 . ISBN 978-1-4615-6819-3 . ПМИД 3077975 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Тадрос, В; Вествуд, Джей Ти; Липшиц, HD (июнь 2007 г.). «Переход от матери к ребенку» . Развивающая клетка . 12 (6): 847–9. дои : 10.1016/j.devcel.2007.05.009 . ПМИД 17543857 .
- ^ Вейгель, Д; Изаурральде, Э. (24 марта 2006 г.). «Крошечный помощник облегчает материнскую ношу» . Клетка . 124 (6): 1117–8. дои : 10.1016/j.cell.2006.03.005 . ПМИД 16564001 .
- ^ Jump up to: Перейти обратно: а б с д Флеминг, Том П.; Папенброк, Том; Фесенко Ирина; Хаузен, Питер; Шет, Бхавванти (1 августа 2000 г.). «Сборка плотных соединений на раннем этапе развития позвоночных». Семинары по клеточной биологии и биологии развития . 11 (4): 291–299. дои : 10.1006/scdb.2000.0179 . ПМИД 10966863 .
- ^ Уотсон, Дж. Г. (октябрь 1977 г.). «Сбор и перенос предимплантационных эмбрионов мыши» . Биология размножения . 17 (3): 453–8. дои : 10.1095/biolreprod17.3.453 . ПМИД 901897 .
- ^ Вичиан, Андреа С.; Солессио, Эдуардо К.; Лю, Юнг; Зубер, Майкл Э. (август 2009 г.). «Поколение функциональных глаз из плюрипотентных клеток» . ПЛОС Биология . 7 (8): e1000174. дои : 10.1371/journal.pbio.1000174 . ПМЦ 2716519 . ПМИД 19688031 .
Библиография
[ редактировать ]- Форгач Г. и Ньюман Стюарт А. (2005). «Дробление и образование бластулы» . Биологическая физика развивающегося эмбриона . Издательство Кембриджского университета. ISBN 978-0-521-78337-8 .
- Каллен, К.Э. (2009). «эмбриология и раннее развитие животных» . Энциклопедия наук о жизни, Том 2 . Информационная база. ISBN 978-0-8160-7008-4 .
- Макгиди, Томас А., изд. (2006). «Гаструляция» . Ветеринарная эмбриология . Уайли-Блэквелл. ISBN 978-1-4051-1147-8 .