Эмбриональное развитие животных
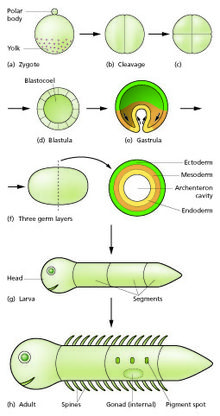
В биологии развития эмбриональное развитие животных , также известное как эмбриогенез животных , представляет собой стадию развития животного эмбриона . Эмбриональное развитие начинается с оплодотворения яйцеклетки ) (яйцеклетки) сперматозоидом ( сперматозоидом . [1] После оплодотворения яйцеклетка становится одной диплоидной клеткой, известной как зигота . Зигота подвергается митотическим делениям без значительного роста (процесс, известный как дробление ) и клеточной дифференциации , что приводит к развитию многоклеточного эмбриона. [2] после прохождения организационного контрольно-пропускного пункта в середине эмбриогенеза. [3] У млекопитающих этот термин относится главным образом к ранним стадиям пренатального развития , тогда как термины «плод» и «развитие плода» описывают более поздние стадии. [2] [4]
Основные этапы эмбрионального развития животных следующие:
- Зигота . претерпевает серию клеточных делений (так называемое расщепление), образуя структуру, называемую морулой
- Морула развивается в структуру, называемую бластулой, посредством процесса, называемого бластуляцией .
- Бластула развивается в структуру, называемую гаструлой, посредством процесса, называемого гаструляцией .
- Затем гаструла подвергается дальнейшему развитию, включая образование органов ( органогенез ).
Затем эмбрион переходит на следующую стадию развития, характер которой варьируется у разных видов животных (примерами возможных следующих стадий являются плод и личинка ).
Оплодотворение и зигота [ править ]
Яйцеклетка в целом асимметрична, имеет анимальный полюс (будущую эктодерму ).Он покрыт защитными конвертами с разными слоями. Первая оболочка, соприкасающаяся с оболочкой яйца , состоит из гликопротеинов и известна как желточная оболочка ( zona pellucida у млекопитающих ). Различные таксоны имеют разные клеточные и бесклеточные оболочки, окружающие желточную мембрану. [2] [5]
Оплодотворение – это слияние гамет с образованием нового организма. У животных этот процесс включает слияние сперматозоида с яйцеклеткой , что в конечном итоге приводит к развитию эмбриона . В зависимости от вида животного процесс может происходить внутри организма самки при внутреннем оплодотворении или снаружи при наружном оплодотворении. Оплодотворенная яйцеклетка известна как зигота . [2] [5]
Чтобы предотвратить оплодотворение яйцеклетки более чем одним сперматозоидом ( полиспермия ), используются быстрый блок и медленный блок полиспермии. Быстрый блок, когда мембранный потенциал быстро деполяризуется, а затем возвращается к норме, происходит сразу после оплодотворения яйцеклетки одним сперматозоидом. Медленный блок начинается в первые несколько секунд после оплодотворения и происходит, когда высвобождение кальция вызывает кортикальную реакцию , при которой из кортикальных гранул в плазматической мембране яйцеклетки высвобождаются различные ферменты, вызывая расширение и затвердевание внешней мембраны, предотвращая появление большего количества сперматозоидов. от входа. [6] [5]
Расщепление и морула [ править ]

Деление клеток без значительного роста, в результате которого образуется кластер клеток того же размера, что и исходная зигота, называется дроблением . Происходит как минимум четыре начальных клеточных деления, в результате чего образуется плотный клубок, состоящий как минимум из шестнадцати клеток, называемый морулой . У ранних эмбрионов мышей сестринские клетки каждого деления остаются соединенными во время интерфазы мостиками из микротрубочек . [7] Различные клетки, полученные в результате дробления вплоть до стадии бластулы , называются бластомерами . В зависимости от количества желтка в яйце расщепление может быть голобластическим (тотальным) или меробластическим (частичным). [8] [9]
Голобластическое дробление происходит у животных, у которых в яйцах мало желтка. [10] например, люди и другие млекопитающие, которые получают питание в виде эмбрионов от матери через плаценту или молоко , которое может выделяться из сумчатых . Меробластическое дробление происходит у животных, яйца которых содержат больше желтка (например, у птиц и рептилий). Поскольку расщепление затруднено на вегетативном полюсе , наблюдается неравномерное распределение и размер клеток: на анимальном полюсе зиготы они более многочисленны и мельче. [8] [9]
В голобластических яйцах первое дробление всегда происходит вдоль растительно-животной оси яйца, а второе дробление перпендикулярно первому. Отсюда пространственное расположение бластомеров может иметь различные закономерности, обусловленные разными плоскостями расщепления, у разных организмов:
| Голобластический | Меробластический |
|---|---|
|
Конец дробления известен как переход средней бластулы и совпадает с началом зиготической транскрипции .
У амниот клетки морулы сначала плотно агрегируются, но вскоре образуют наружный или периферический слой — трофобласт , не способствующий образованию собственно зародыша, и внутреннюю клеточную массу , из которой эмбрион развит. Жидкость скапливается между трофобластом и большей частью внутренней клеточной массы, и таким образом морула превращается в везикулу , называемую бластодермическим пузырьком. Однако внутренняя клеточная масса остается в контакте с трофобластом на одном полюсе яйцеклетки; это называется эмбриональным полюсом, поскольку указывает место, где будет развиваться будущий эмбрион. [18] [9]
Образование бластулы [ править ]
После того, как в результате седьмого дробления образовалось 128 клеток , морула становится бластулой . [8] Бластула обычно представляет собой сферический слой клеток ( бластодерма ), окружающий полость, заполненную жидкостью или желтком, бластоцель . [ нужна ссылка ]
Млекопитающие на этом этапе образуют структуру, называемую бластоцистой . [1] характеризуется внутренней клеточной массой , отличной от окружающей бластулы. [19] [20] [21] Бластоциста по строению похожа на бластулу, но их клетки имеют разную судьбу. У мышей первичные зародышевые клетки возникают из внутренней клеточной массы ( эпибласта ) в результате обширного репрограммирования всего генома . [22] Перепрограммирование включает глобальное деметилирование ДНК, чему способствует путь репарации вырезания оснований ДНК, а также реорганизация хроматина и приводит к клеточной тотипотентности . [23] [20]
Перед гаструляцией клетки трофобласта дифференцируются на два слоя: внешний слой образует синцитий (т. е. слой протоплазмы, усеянный ядрами, но не имеющий признаков разделения на клетки), называемый синцитиотрофобластом , а внутренний слой - цитотрофобласт , состоит из четко определенных клеток. Как уже говорилось, клетки трофобласта не способствуют образованию собственно зародыша; они образуют эктодерму хориона и играют важную роль в развитии плаценты . На глубокой поверхности внутренней клеточной массы слой уплощенных клеток, называемый энтодермой , дифференцируется и быстро принимает форму небольшого мешочка, называемого желточным мешком . Между остальными клетками массы появляются промежутки, и в результате расширения и слияния этих пространств полость, называемая амниотической полостью постепенно развивается . Дно этой полости образовано эмбриональным диском , который состоит из слоя призматических клеток — эмбриональной эктодермы, происходящей из внутренней клеточной массы и лежащей в аппозиции с энтодермой. [18] [20]
Формирование зародышевых листков [ править ]

Эмбриональный диск становится овальным, а затем грушевидным, причем более широкий конец направлен вперед. К узкому, заднему концу образуется непрозрачная примитивная полоска , простирающаяся вдоль середины диска примерно на половину его длины; на переднем конце полоски имеется бугорковидное утолщение, называемое примитивным узлом или узлом (у птиц известное как узел Генсена ). На поверхности полоски появляется неглубокая бороздка — примитивная бороздка , передний конец которой сообщается посредством отверстия — бластопора — с желточным мешком . Первичная полоска образуется в результате утолщения осевой части эктодермы, клетки которой размножаются, растут вниз и сливаются с клетками нижележащей энтодермы. С боков первичной полоски третий слой клеток — мезодерма латерально между эктодермой и энтодермой простирается ; каудальный мембрану конец первичной полоски образует клоакальную . Бластодерма теперь состоит из трех слоев: внешней эктодермы, средней мезодермы и внутренней энтодермы; каждый имеет отличительные характеристики и дает начало определенным тканям организма. У многих млекопитающих это происходит на этапе формирования зародышевых листков. происходит имплантация эмбриона в матку матери. [18] [20]
Формирование гаструлы [ править ]
Во время гаструляции клетки мигрируют внутрь бластулы, образуя впоследствии два (у диплобластных животных) или три ( триблобластные ) зародышевых листка . Зародыш во время этого процесса называется гаструлой . Зародышевые листки называются эктодермой, мезодермой и энтодермой. У диплобластических животных присутствуют только эктодерма и энтодерма. [8] * У разных животных происходят разные комбинации следующих процессов для размещения клеток внутри эмбриона:
- Эпиболия – расширение одного клеточного листа над другими клетками. [2] [9]
- Ингрессия – миграция отдельных клеток в эмбрион (клетки перемещаются ложноножками ). [3] [9]
- Инвагинация – сворачивание клеточного листа в зародыш, образуя рот , анус и архентерон . [8] [9]
- Деламинация – разделение или миграция одного листа на два листа. [9]
- Инволюция – переворачивание клеточного листа по базальной поверхности наружного слоя. [9]
- Полярная пролиферация. Клетки на полярных концах бластулы/гаструлы пролиферируют, в основном на анимальном полюсе. [9]
- Другие важные изменения во время гаструляции:
- Транскрипция тяжелой РНК с использованием эмбриональных генов; до этого момента используемые РНК были материнскими (хранились в неоплодотворенной яйцеклетке).
- Клетки запускают основные процессы дифференцировки , теряя свой тотипотенциал .
У большинства животных бластопор образуется в месте миграции клеток внутрь. можно выделить две основные группы животных В зависимости от судьбы бластопора . У вторичноротых анус формируется из бластопора, а у протостомов — в рот. [9]
ранней нервной системы - нервная борозда, трубка Формирование и хорда
Перед первичной полоской появляются два продольных гребня, образовавшиеся в результате складывания эктодермы, по одному с каждой стороны от средней линии, образованной полоской. Их называют нервными складками ; они начинаются на некотором небольшом расстоянии позади переднего конца зародышевого диска , где переходят друг в друга, и оттуда постепенно распространяются назад, по одному с каждой стороны от переднего конца первичной полоски. Между этими складками находится неглубокая срединная борозда — нервная борозда . Борозда постепенно углубляется по мере поднятия нервных складок и в конечном итоге складки встречаются и сливаются по средней линии и превращают борозду в замкнутую трубку, нервную трубку или канал, эктодермальная стенка которого образует зачаток нервной системы. После слияния нервных складок на переднем конце первичной полоски бластопор открывается уже не на поверхности, а в закрытый канал нервной трубки и, таким образом, образует временную коммуникацию, нервно-кишечный канал , устанавливается между нервной трубкой и первичной пищеварительной трубкой . Слияние нервных складок происходит сначала в области заднего мозга , а оттуда распространяется вперед и назад; к концу третьей недели переднее отверстие ( передняя нейропора ) трубки окончательно закрывается на переднем конце будущего мозга и образует углубление, соприкасающееся на время с вышележащей эктодермой; задняя часть нервной борозды какое-то время имеет ромбовидную форму термин sinus rhomboidalis , и к этой расширенной части был применен . Прежде чем нервная борозда закрывается, вдоль выступающего края каждой нервной складки появляется гребень эктодермальных клеток; это называется нервным гребнем или ганглиозным гребнем, и из него ганглии спинномозговых и черепных нервов, а также ганглии симпатической нервной системы . развиваются [ нужна ссылка ] В результате роста мезодермы вверх нервная трубка в конечном итоге отделяется от вышележащей эктодермы. [24] [9]

Головной . конец нервной борозды имеет несколько расширений, которые при закрытии трубки принимают форму трех первичных мозговых пузырьков и соответствуют соответственно будущему переднему мозгу (prosencephalon), среднему мозгу (mesencephalon) и заднему мозгу (rhombencephalon) ) (рис. 18). Стенки везикул развиваются в нервную ткань и нейроглию головного мозга, а их полости видоизменяются, образуя его желудочки. Остальная часть трубки образует спинной мозг (medulla spinalis); из его эктодермальной стенки развиваются нервные и нейроглиальные элементы спинного мозга, а полость сохраняется в виде центрального канала . [24] [9]
Формирование ранней перегородки [ править ]
Распространение мезодермы происходит по всей эмбриональной и внеэмбриональной части яйцеклетки, за исключением отдельных участков. Один из них виден непосредственно перед нервной трубкой. Здесь мезодерма выдвигается вперед в виде двух серповидных масс, которые сходятся по средней линии так, что замыкают за собой участок, лишенный мезодермы. На этом участке эктодерма и энтодерма непосредственно соприкасаются друг с другом и образуют тонкую мембрану — щечно-глоточную мембрану , образующую перегородку между примитивным ртом и глоткой . [18] [9]
Раннее формирование сердца и других примитивных структур [ править ]
Перед щечноглоточной областью, где латеральные полулуния мезодермы сливаются по средней линии, впоследствии развивается перикард , и поэтому эта область называется областью перикарда. Вторая область, где мезодерма отсутствует, по крайней мере какое-то время, находится непосредственно перед областью перикарда. Это называется проамниотической областью, и это область, где развивается проамнион; однако у людей оказывается, что проамнион никогда не формируется. Третья область находится на заднем конце эмбриона, где эктодерма и энтодерма соединяются и образуют клоакальную мембрану. [18] [9]
Somitogenesis[editСомитогенез
Сомитогенез — это процесс сомитов образования (примитивных сегментов). Эти сегментированные тканевые блоки дифференцируются в скелетные мышцы, позвонки и дерму всех позвоночных. [25]
Сомитогенез начинается с образования сомитомеров (витков концентрической мезодермы), обозначающих будущие сомиты в пресомитной мезодерме (несегментированной параксиальной мезодерме). Пресомитная мезодерма дает начало последовательным парам сомитов, идентичных по внешнему виду, которые дифференцируются в одни и те же типы клеток, но структуры, образуемые клетками, различаются в зависимости от передне-задней части (например, грудные позвонки имеют ребра, а поясничные - нет). Сомиты имеют уникальные позиционные значения вдоль этой оси, и считается, что они определяются Hox гомеозисными генами . [25]
К концу второй недели после оплодотворения начинается поперечная сегментация параксиальной мезодермы , и она превращается в ряд четко выраженных, более или менее кубических масс, известных также как сомиты, занимающих всю длину туловища. по обе стороны от средней линии от затылочной области головы. Каждый сегмент содержит центральную полость (известную как миоцель), которая, однако, вскоре заполняется угловатыми и веретенообразными клетками. Сомиты лежат непосредственно под эктодермой на латеральной стороне нервной трубки и хорды и соединены с латеральной мезодермой промежуточной клеточной массой . Те части туловища можно разделить на следующие группы, а именно: шейные 8, грудные 12, поясничные 5, крестцовые 5 и копчиковые от 5 до 8. Те, что находятся в затылочной области головы, обычно описываются как четыре. . У млекопитающих сомиты головы можно распознать только в затылочной области, но изучение низших позвоночных приводит к убеждению, что они присутствуют и в передней части головы и что всего в ней представлены девять сегментов. головной отдел. [26] [25]
Органогенез [ править ]
В какой-то момент после определения различных зародышевых листков органогенез начинается . Первая стадия у позвоночных называется нейруляцией , когда нервная пластинка складывается, образуя нервную трубку (см. выше). [8] Другие общие органы или структуры, возникающие в это время, включают сердце и сомиты (также выше), но с этого момента эмбриогенез не следует общей схеме среди различных таксонов животных . [2]
У большинства животных органогенез наряду с морфогенезом приводит к образованию личинки . Вылупление личинки, которая затем должна подвергнуться метаморфозу , знаменует собой окончание эмбрионального развития. [2]
См. также [ править ]
Ссылки [ править ]
- ^ Гилберт, Скотт (2000). Биология развития. 6-е издание. Глава 7. Оплодотворение: зарождение нового организма . Проверено 3 октября 2020 г. .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Гилберт, Скотт (2000). Биология развития. 6-е издание. Круг жизни: этапы развития животных . Проверено 3 октября 2020 г. .
- ^ Дрост, Хайк-Георг; Яница, Филипп; Гросс, Иво; Квинт, Марсель (2017). «Сравнение песочных часов развития между королевствами» . Текущее мнение в области генетики и развития . 45 : 69–75. дои : 10.1016/j.где.2017.03.003 . ПМИД 28347942 .
- ^ Гилберт, Скотт (2000). Биология развития. 6-е издание. Раннее развитие млекопитающих . Проверено 3 октября 2020 г. .
- ↑ Перейти обратно: Перейти обратно: а б с Хинтон-Шели, Фиби. «Этапы раннего эмбрионального развития» . Проверено 6 октября 2020 г.
- ^ Альбертс, Брюс; Джонсон, Александр; Льюис, Джулиан; Рафф, Мартин; Робертс, Кейт; Уолтер, Питер (2002). «Оплодотворение» . Архивировано из оригинала 14 мая 2017 г.
- ^ Зенкер, Дж.; Уайт, доктор медицины; Темплин, РМ; Партон, Р.Г.; Торн-Сешолд, О.; Биссьер, С.; Плахта, Н. (сентябрь 2017 г.). «Центр организации микротрубочек, управляющий внутриклеточным транспортом у раннего эмбриона мыши» . Наука . 357 (6354): 925–928. Бибкод : 2017Sci...357..925Z . дои : 10.1126/science.aam9335 . ISSN 0036-8075 . ПМИД 28860385 . S2CID 206658036 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Что такое клетка? Архивировано 18 января 2006 г. в Wayback Machine 2004. Учебник по науке: базовое введение в науку, лежащую в основе ресурсов NCBI. НЦБИ; и Кэмпбелл, Нил А.; Рис, Джейн Б.; Биология Бенджамин Каммингс, Pearson Education, 2002.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в Гилберт, Скотт (2000). Биология развития. 6-е издание. Введение в процесс раннего развития . Проверено 3 октября 2020 г. .
- ^ Гилберт, Скотт (2000). Раннее развитие нематоды Caenorhabditis elegans (Биология развития, 6-е изд.) . Проверено 3 октября 2020 г. .
- ^ Гилберт, Скотт (2000). Биология развития. 6-е издание. Раннее развитие морских ежей . Проверено 3 октября 2020 г. .
- ^ Гилберт, Скотт (2000). Биология развития. 6-е издание. Раннее развитие оболочников . Проверено 4 октября 2020 г.
- ^ Гилберт, Скотт (2000). Биология развития. 6-е издание. Раннее развитие амфибий . Проверено 3 октября 2020 г. .
- ^ Гилберт, Скотт (2000). Биология развития. 6-е издание. Раннее развитие улиток . Проверено 4 октября 2020 г.
- ↑ Перейти обратно: Перейти обратно: а б Гилберт, Скотт (2000). Биология развития. 6-е издание. Глава 11. Раннее развитие позвоночных животных: Рыбы, птицы и млекопитающие . Проверено 3 октября 2020 г. .
- ^ Гилберт, Скотт (2000). Биология развития. 6-е издание. Раннее развитие нематоды Caenorhabditis elegans . Проверено 4 октября 2020 г.
- ^ Гилберт, Скотт (2000). Биология развития. 6-е издание. Раннее развитие дрозофилы . Проверено 4 октября 2020 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и "Йаху" . Яху . Архивировано из оригинала 22 декабря 2009 г.
- ^ Ниссен С.Б., Перера М., Гонсалес Дж.М., Моргани С.М., Дженсен М.Х., Снеппен К. и др. (июль 2017 г.). «Четыре простых правила, которых достаточно для образования бластоцисты млекопитающих» . ПЛОС Биология . 15 (7): e2000737. дои : 10.1371/journal.pbio.2000737 . ПМК 5507476 . ПМИД 28700688 .
- ↑ Перейти обратно: Перейти обратно: а б с д Балано, Алекс (25 февраля 2019 г.). «Что такое бластоциста» . Тенденции науки . Проверено 5 октября 2020 г.
- ^ «Бластула» . Британская энциклопедия . Проверено 5 октября 2020 г.
- ^ Хакетт Дж.А., Сенгупта Р., Зилич Дж.Дж., Мураками К., Ли С., Даун Т.А., Сурани М.А. (январь 2013 г.). «Динамика деметилирования зародышевой ДНК и стирание отпечатка с помощью 5-гидроксиметилцитозина» . Наука . 339 (6118): 448–52. Бибкод : 2013Sci...339..448H . дои : 10.1126/science.1229277 . ПМЦ 3847602 . ПМИД 23223451 .
- ^ Хайкова П., Джеффрис С.Дж., Ли С., Миллер Н., Джексон С.П., Сурани М.А. (июль 2010 г.). «Полногеномное перепрограммирование в зародышевой линии мыши влечет за собой путь эксцизионной репарации основания» . Наука . 329 (5987): 78–82. Бибкод : 2010Sci...329...78H . дои : 10.1126/science.1187945 . ПМЦ 3863715 . ПМИД 20595612 .
- ↑ Перейти обратно: Перейти обратно: а б «Нейральная канавка и трубка» . Яху . Архивировано из оригинала 22 августа 2007 г.
- ↑ Перейти обратно: Перейти обратно: а б с Пурке, Оливер (ноябрь 2001 г.). «Сомитогенез позвоночных» . Ежегодный обзор клеточной биологии и биологии развития . 17 : 311–350. дои : 10.1146/annurev.cellbio.17.1.311 . ПМИД 11687492 . Проверено 5 октября 2020 г.
- ^ «Примитивные сегменты» . Яху . Архивировано из оригинала 11 сентября 2007 г.
Внешние ссылки [ править ]
- Клеточный дарвинизм
- Эмбриогенез и ММП , PMAP Карта протеолиза - анимация
- Развитие эмбриона (получено 20 ноября 2007 г.)
- Видео эмбриогенеза лягушки Xenopus laevis вскоре после оплодотворения до вылупления головастика; получено с помощью МРТ ( DOI бумаги )