Гаструляция
| Гаструляция | |
|---|---|
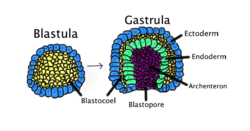 Гаструляция происходит, когда бластула , состоящая из одного слоя, складывается внутрь и увеличивается, образуя гаструлу. Эта диаграмма имеет цветовую маркировку: эктодерма — синий; эндодерма , зеленая; бластоцель (желточный мешок) желтый; и архентерон (первичная кишка) — фиолетовый. | |
| Идентификаторы | |
| МеШ | D054262 |
| Анатомическая терминология | |
Гаструляция — стадия раннего эмбрионального развития большинства животных , во время которой бластула (однослойная полая сфера клеток ) или у млекопитающих бластоциста реорганизуется в двухслойный или трехслойный зародыш, известный как гаструла. . [1] До гаструляции зародыш представляет собой сплошной эпителиальный листок клеток; к концу гаструляции эмбрион начинает дифференцировку, образуя отдельные клеточные линии , устанавливает основные оси тела (например, дорсально-вентральную , передне-заднюю ) и интернализует один или несколько типов клеток, включая проспективную кишку . [2]
Слои гаструлы
[ редактировать ]У триплобластных организмов гаструла трехламинарная (трехслойная). Этими тремя зародышевыми листками являются эктодерма (наружный слой), мезодерма (средний слой) и энтодерма (внутренний слой). [3] [4] У диплобластических организмов, таких как Cnidaria и Ctenophora , гаструла имеет только эктодерму и энтодерму. Эти два слоя также иногда называют гипобластом и эпибластом . [5] Губки не проходят стадию гаструлы.
Гаструляция происходит после дробления и образования бластулы или бластоцисты. За гаструляцией следует органогенез , когда отдельные органы развиваются внутри вновь образовавшихся зародышевых листков. [6] Каждый слой дает начало определенным тканям и органам развивающегося эмбриона.
- Эктодерма дает начало эпидермису , нервной системе и нервному гребню у позвоночных. [2]
- Энтодерма дает начало эпителию пищеварительной и системы дыхательной системы , а также органов, связанных с пищеварительной системой, таких как печень и поджелудочная железа . [2]
- Мезодерма мышечная дает начало многим типам клеток, таких как , костная и соединительная ткань . У позвоночных производные мезодермы включают хорду , сердце , кровь и кровеносные сосуды , хрящи ребер , и позвонков а также дерму . [2] [7]
После гаструляции клетки организма либо организуются в листы связанных клеток (как в эпителии ), либо в виде сетки изолированных клеток, например, мезенхимы . [4] [8]
Основные движения клеток
[ редактировать ]Хотя закономерности гаструляции демонстрируют огромные различия во всем животном мире, их объединяют пять основных типов движений клеток, которые происходят во время гаструляции: [2] [9]
Этимология
[ редактировать ]Термины «гаструла» и «гаструляция» были придуманы Эрнстом Геккелем в его работе 1872 года «Биология известковых губок» . [10] Гаструла (буквально «живот») — неолатинское уменьшительное слово, основанное на древнегреческом γαστήρ gastḗr («живот»).
Важность
[ редактировать ]Льюису Уолперту , новаторскому биологу развития в этой области, приписывают замечание: «Не рождение, брак или смерть, а гаструляция действительно является самым важным периодом в вашей жизни». [2] [11]
Модельные системы
[ редактировать ]Гаструляция сильно различается в животном мире, но имеет основные сходства. Гаструляцию изучали на многих животных, но некоторые модели использовались дольше, чем другие. Кроме того, легче изучать развитие животных, развивающихся вне матери. К модельным организмам, гаструляция которых изучена наиболее детально, относятся моллюски , морские ежи , лягушки и курицы . Модельной системой человека является гаструлоид .
Протостомы против вторичноротых
[ редактировать ]
Различие между протостомами и вторичноротыми основано на направлении развития рта (стомы) по отношению к бластопору . Протостома происходит от греческого слова protostoma, означающего «первый рот» (πρῶτος + στόμα), тогда как этимология Второстома — «второй рот» от слов второй и рот (δεύτερος + στόμα). [ нужна ссылка ]
Основные различия между вторичноротыми и протостомами обнаруживаются в эмбриональном развитии :
- Рот/анус
- В развитии протостома животного первым отверстием в развитии, бластопером, становится рот .
- При развитии дейтеростома животного бластопор становится анусом .
- Расщепление
- У протостом есть так называемое спиральное расщепление , которое является детерминантным , что означает, что судьба клеток определяется по мере их формирования.
- У вторичноротых есть так называемое радиальное расщепление , которое является неопределенным .
Морские ежи
[ редактировать ]Морские ежи были важными модельными организмами в биологии развития с 19 века. [12] Их гаструляцию часто считают архетипом вторичноротых беспозвоночных. [13] Для получения знаний о гаструляции морского ежа использовались эксперименты и компьютерное моделирование. Недавнее моделирование показало, что полярности плоских клеток достаточно, чтобы стимулировать гаструляцию морских ежей. [14]
Определение зародышевого слоя
[ редактировать ]Морские ежи демонстрируют весьма стереотипные модели расщепления и судьбы клеток. Депонированные у матери мРНК создают организующий центр эмбриона морского ежа. Каноническая передача сигналов Wnt и Delta-Notch постепенно разделяет прогрессирующую энтодерму и мезодерму. [15]
Интернализация клеток
[ редактировать ]У морских ежей первыми интернализующимися клетками являются первичные мезенхимные клетки (PMC), имеющие скелетогенную судьбу и проникающие на стадии бластулы. Гаструляция – интернализация проспективной энтодермы и нескелетогенной мезодермы – начинается вскоре после этого с инвагинации и других клеточных перестроек вегетативного полюса , которые составляют примерно 30% окончательной длины архентерона . зависит Окончательная длина кишечника от перестройки клеток внутри архентерона. [16]
Земноводные
[ редактировать ]Род лягушек модельного Xenopus использовался в качестве организма для изучения гаструляции. [17]
Нарушение симметрии
[ редактировать ]Сперматозоид дает одну из двух митотических астр, необходимых для завершения первого дробления. Сперматозоид может проникнуть в любую часть животной половины яйцеклетки, но его точная точка входа нарушит радиальную симметрию яйцеклетки, организовав цитоскелет . Перед первым дроблением кора яйцеклетки вращается относительно внутренней цитоплазмы за счет скоординированного действия микротрубочек в процессе, известном как вращение коры. Это смещение приводит в контакт материнские детерминанты клеточной судьбы из экваториальной цитоплазмы и вегетативной коры, и вместе эти детерминанты создают организатор . Таким образом, организатором станет область на вегетативной стороне, противоположная месту входа сперматозоидов. [18] Хильда Мангольд , работавшая в лаборатории Ганса Спеманна , продемонстрировала, что этот особый «организатор» зародыша необходим и достаточен для индукции гаструляции. [19] [20] [21]
Определение зародышевого слоя
[ редактировать ]Спецификация эндодермы зависит от перестройки депонированных материнских детерминант, что приводит к нуклеаризации Beta-catenin . Мезодерма индуцируется передачей сигналов от презумптивной энтодермы к клеткам, которые в противном случае стали бы эктодермой. [18]
Интернализация клеток
[ редактировать ]Дорсальная губа бластопора является механическим двигателем гаструляции. Первым признаком инвагинации у лягушки является спинная губа. [ нужна ссылка ]
Передача сигналов ячейки
[ редактировать ]У лягушки Xenopus одним из сигналов является ретиноевая кислота (РА). [22] Передача сигналов RA в этом организме может влиять на формирование энтодермы и, в зависимости от времени передачи сигналов, может определять судьбу панкреатического, кишечного или дыхательного пути. Другие сигналы, такие как Wnt и BMP, также играют роль в респираторной судьбе Xenopus путем активации индикаторов клеточных клонов. [22]
Амниоты
[ редактировать ]Обзор
[ редактировать ]У амниот (рептилий, птиц и млекопитающих) гаструляция включает создание бластопора — отверстия в архентероне . Обратите внимание, что бластопор не является отверстием в бластоцеле , пространстве внутри бластулы , а представляет собой новый карман, который сближает существующие поверхности бластулы. У амниот гаструляция происходит в следующей последовательности: (1) зародыш становится асимметричным ; (2) примитивные полоски формируются ; (3) клетки эпибласта первичной полоски подвергаются эпителиально -мезенхимальному переходу и проникают в примитивную полоску с образованием зародышевых листков . [7]
Нарушение симметрии
[ редактировать ]При подготовке к гаструляции эмбрион должен стать асимметричным как по проксимально-дистальной оси, так и по переднезадней оси . Проксимально-дистальная ось формируется, когда клетки эмбриона образуют «цилиндр яйца», который состоит из внеэмбриональных тканей, дающих начало таким структурам, как плацента , на проксимальном конце и эпибласт на дистальном конце. Многие сигнальные пути способствуют этой реорганизации, включая BMP , FGF , nodal и Wnt . Висцеральная энтодерма окружает эпибласт . Дистальная висцеральная энтодерма ( DVE ) мигрирует в переднюю часть эмбриона, образуя переднюю висцеральную энтодерму (AVE). Это нарушает передне-заднюю симметрию и регулируется узловой передачей сигналов. [7]
Определение зародышевого слоя
[ редактировать ]Примитивная полоска формируется в начале гаструляции и находится в месте соединения внеэмбриональной ткани с эпибластом на задней стороне зародыша и в месте ингрессии . [23] Формирование примитивной полоски зависит от узловой передачи сигналов. [7] в серпе Коллера внутри клеток, участвующих в первичной полоске и передаче сигналов BMP4 из внеэмбриональной ткани. [23] [24] Более того, Cer1 и Lefty1 ограничивают примитивную полоску соответствующим местоположением, противодействуя передаче узловых сигналов. [25] Область, определяемая как примитивная полоска, продолжает расти к дистальному кончику. [7]
На ранних стадиях развития примитивная полоска представляет собой структуру, которая устанавливает двустороннюю симметрию , определяет место гаструляции и инициирует образование зародышевого листка. [26] Для формирования полоски рептилии, птицы и млекопитающие располагают мезенхимальные клетки вдоль предполагаемой срединной линии, устанавливая первую эмбриональную ось, а также место, куда клетки будут проникать и мигрировать в процессе гаструляции и формирования зародышевого листка. [27] Примитивная полоска проходит через эту срединную линию и образует передне-заднюю ось тела. [28] становится первым событием, нарушающим симметрию эмбриона , и знаменует начало гаструляции. [29] Этот процесс включает в себя проникновение предшественников мезодермы и энтодермы и их миграцию в конечное положение. [28] [30] где они дифференцируются в три зародышевых листка. [27] Локализация клеточной адгезии и сигнальной молекулы бета-катенина имеет решающее значение для правильного формирования области-организатора, которая отвечает за инициацию гаструляции.
Интернализация клеток
[ редактировать ]Чтобы клетки могли переместиться из эпителия эпибласта ( через примитивную полоску с образованием нового слоя, клетки должны претерпеть эпителиально-мезенхимальный переход EMT), чтобы потерять свои эпителиальные характеристики, такие как межклеточная адгезия . Передача сигналов FGF необходима для правильного ЕМТ. FGFR1 необходим для повышения регуляции SNAI1 , который снижает регуляцию E-кадгерина , вызывая потерю клеточной адгезии. После ЕМТ клетки проникают через примитивную полоску и распространяются, образуя новый слой клеток или присоединяясь к существующим слоям. FGF8 участвует в процессе расселения из первичной полоски . [25]
Передача сигналов ячейки
[ редактировать ]Существуют определенные сигналы, которые играют роль в детерминации и формировании трех зародышевых листков, таких как FGF, RA и Wnt. [22] У млекопитающих, таких как мыши, передача сигналов RA может играть роль в формировании легких. Если РА недостаточно, произойдет ошибка в работе легких. RA также регулирует дыхательную компетентность в этой мышиной модели. [ нужна ссылка ]
Передача сигналов в клетках, управляющая гаструляцией
[ редактировать ]Во время гаструляции клетки дифференцируются в эктодерму или мезендодерму , которая затем разделяется на мезодерму и энтодерму. [22] Энтодерма и мезодерма формируются за счет узловой передачи сигналов . Для передачи сигналов через узлы используются лиганды, которые являются частью семейства TGFβ . Эти лиганды будут сигнализировать трансмембранные рецепторы серин/треонинкиназы, которые затем фосфорилируют Smad2 и Smad3 . Затем этот белок прикрепится к Smad4 и переместится в ядро, где начнут транскрибироваться гены мезентодермы. Путь Wnt вместе с β-catenin играет ключевую роль в передаче сигналов nodal и формировании энтодермы. [31] Факторы роста фибробластов (FGF), канонический путь Wnt, костный морфогенетический белок (BMP) и ретиноевая кислота (RA) играют важную роль в формировании и развитии энтодермы. [22] FGF играют важную роль в производстве гена гомеобокса , который регулирует раннее анатомическое развитие. Передача сигналов BMP играет роль в печени и способствует судьбе печени. Передача сигналов RA также индуцирует гены гомеобокса, такие как Hoxb1 и Hoxa5. У мышей, если отсутствует передача сигналов RA, легкие не развиваются. [22] Передача сигналов RA также имеет множество применений в формировании органов глоточных дуг, передней и задней кишки. [22]
Гаструляция in vitro
[ редактировать ]Был предпринят ряд попыток понять процессы гаструляции с использованием методов in vitro, параллельных и дополняющих исследования на эмбрионах, обычно с использованием 2D-изображений. [32] [33] [34] 3D-клеток ( эмбриональных органоидов ) и методы культивирования [35] [36] [37] [38] с использованием эмбриональных стволовых клеток (ЭСК) или индуцированных плюрипотентных стволовых клеток (ИПСК). Это связано с рядом явных преимуществ использования протоколов, основанных на культурах тканей, некоторые из которых включают снижение стоимости связанных с ними работ in vivo (тем самым сокращая, заменяя и совершенствуя использование животных в экспериментах; 3R ), возможность точно применять агонисты/антагонисты в определенном пространстве и времени [36] [37] что может быть технически сложно выполнить во время гаструляции. Однако для контекста важно связать наблюдения в культуре с процессами, происходящими в эмбрионе.
Чтобы проиллюстрировать это, управляемая дифференцировка ЭСК мыши привела к образованию примитивных полосоподобных клеток, которые демонстрируют многие характеристики клеток эпибласта, которые проходят через примитивную полоску. [32] (например, временная регуляция брахюрии и клеточные изменения, связанные с эпителиально-мезенхимальным переходом [32] ), а ЭСК человека, культивированные на микроструктурах, обработанные BMP4 , могут генерировать паттерн пространственной дифференциации, аналогичный расположению зародышевых листков в человеческом эмбрионе. [33] [34] Наконец, используя трехмерные методы, основанные на эмбриоидных телах и органоидах , небольшие агрегаты мышиных ЭСК ( эмбриональных органоидов или гаструлоидов ) способны продемонстрировать ряд процессов раннего развития эмбрионов млекопитающих, таких как нарушение симметрии, поляризация экспрессии генов, гаструляция. -подобные движения, осевое удлинение и формирование всех трех эмбриональных осей (передне-задней, дорсовентральной и лево-правой осей). [35] [36] [37] [39]
Экстракорпоральное оплодотворение происходит в лаборатории. Процесс экстракорпорального оплодотворения заключается в том, что зрелые яйцеклетки извлекаются из яичников и помещаются в питательную среду, где они оплодотворяются спермой. В культуре сформируется эмбрион. [40] Через 14 дней после оплодотворения формируется примитивная полоска. Формирование примитивной черты известно в некоторых странах как «человеческая индивидуальность». [41] Это означает, что эмбрион теперь сам является существом, он представляет собой собственную сущность. Страны, которые так считают, создали правило 14 дней, согласно которому запрещено изучать или экспериментировать на человеческом эмбрионе после 14-дневного периода in vitro . Исследования проводились в первые 14 дней эмбриона, но никаких известных исследований после 14 дней не проводилось. [42] При наличии этого правила используются эмбрионы мышей, развитие которых происходит через 14 дней; однако существуют различия в развитии между мышами и людьми.
См. также
[ редактировать ]- Бластоциста
- вторичноротый
- Картирование судьбы
- Примитивный узел
- Инвагинация
- нейруляция
- Протостом
- Вегетативный севооборот
Ссылки
[ редактировать ]Примечания
[ редактировать ]- ^ Урри, Лиза (2016). Кэмпбелл Биология (11-е изд.). Пирсон. п. 1047. ИСБН 978-0134093413 .
- ^ Jump up to: а б с д и ж Гилберт, Скотт Ф.; Майкл Дж. Ф. Баррези (2016). Биология развития (Одиннадцатое изд.). Сандерленд, Массачусетс: Синауэр. ISBN 978-1-60535-470-5 . OCLC 945169933 .
- ^ Безмолвный 2009: с. 422
- ^ Jump up to: а б Макгиди, 2004: с. 34
- ^ Джонатон М.В., Slack (2013). Основная биология развития . Западный Суссекс, Великобритания: Уайли-Блэквелл. п. 122. ИСБН 978-0-470-92351-1 .
- ^ Холл, 1998: стр. 132-134.
- ^ Jump up to: а б с д и Арнольд и Робинсон, 2009 г.
- ^ Холл, 1998: с. 177
- ^ Гилберт, Скотт Ф. (2000). «Рисунок 8.6, [Типы движений клеток во время...]» . www.ncbi.nlm.nih.gov . Проверено 11 мая 2022 г.
- ^ Эресковский 2010: с. 236
- ^ Wolpert L (2008) Триумф эмбриона . Курьерская корпорация, стр. 12. ISBN 978-0-486-46929-4
- ^ Лаубихлер, доктор медицинских наук и Дэвидсон, Э.Х. (2008). «Длительный эксперимент Бовери: мерогоны морских ежей и установление роли ядерных хромосом в развитии». Биология развития . 314(1):1–11. дои : 10.1016/j.ydbio.2007.11.024 .
- ^ Макклей, Дэвид Р.; Гросс, Дж. М.; Рэндж, Райан; Петерсон, Р.Э.; Брэдэм, Синтия (2004). «Глава 9: Гаструляция морского ежа». В Штерне, Клаудио Д. (ред.). Гаструляция: от клеток к эмбрионам . Лабораторный пресс Колд-Спринг-Харбор. стр. 123–137. ISBN 978-0-87969-707-5 .
- ^ Нильсен, Бьярке Фрост; Ниссен, Сайлас Бойе; Снеппен, Ким; Матисен, Иоахим; Трусина Аля (21 февраля 2020 г.). «Модель, связывающая форму и полярность клеток с органогенезом» . iScience . 23 (2): 100830. Бибкод : 2020iSci...23j0830N . дои : 10.1016/j.isci.2020.100830 . ПМК 6994644 . ПМИД 31986479 . S2CID 210934521 .
- ^ МакКлей, Д.Р. 2009. Дробление и гаструляция морского ежа. эЛС. два : 10.1002/9780470015902.a0001073.pub2
- ^ Хардин Дж.Д. (1990). «Контекстно-зависимое поведение клеток во время гаструляции» (PDF) . Семин. Дев. Биол . 1 : 335–345.
- ^ Блюм, Мартин; Бейер, Тина; Вебер, Томас; Вик, Филипп; Андре, Филипп; Битцер, Ева; Швейкерт, Аксель (июнь 2009 г.). «Xenopus, идеальная модельная система для изучения лево-правой асимметрии позвоночных» . Динамика развития . 238 (6): 1215–1225. дои : 10.1002/dvdy.21855 . ПМИД 19208433 . S2CID 39348233 .
- ^ Jump up to: а б Гилберт, Скотт Ф. (2000). «Формирование осей у амфибий: феномен организатора, прогрессивное определение осей земноводных» . Биология развития . Синауэр Ассошиэйтс.
- ^ Гилберт, Скотт Ф. (2000). «Рисунок 10.20, [Организация вторичной оси...]» . www.ncbi.nlm.nih.gov . Проверено 1 июня 2020 г.
- ^ Спеманн Х., Мангольд Х. (1924). «Об индукции эмбриональных зачатков путем имплантации чужеродных организаторов». Ру Арч. Ф. Дев . 100 (3–4): 599–638. дои : 10.1007/bf02108133 . S2CID 12605303 .
- ^ Де Робертис Эдвард (2006). «Организатор Спемана и саморегуляция у эмбрионов амфибий» . Nature Reviews Молекулярно-клеточная биология . 7 (4): 296–302. дои : 10.1038/nrm1855 . ПМЦ 2464568 . ПМИД 16482093 .
- ^ Jump up to: а б с д и ж г Зорн А., Уэллс Дж. (2009). «Развитие эндодермы позвоночных и формирование органов» . Annu Rev Cell Dev Biol . 25 : 221–251. doi : 10.1146/annurev.cellbio.042308.113344 . ПМЦ 2861293 . ПМИД 19575677 .
- ^ Jump up to: а б Тэм и Берингер, 1997 г.
- ^ Катала, 2005: с. 1535 г.
- ^ Jump up to: а б Тэм, ПП; Лебель, Д.А. (2007). «Функция гена в эмбриогенезе мышей: подготовка к гаструляции». Нат преподобный Жене . 8 (5): 368–81. дои : 10.1038/nrg2084 . ПМИД 17387317 . S2CID 138874 .
- ^ Шэн, Гоцзюнь; Ариас, Альфонсо Мартинес; Сазерленд, Энн (3 декабря 2021 г.). «Примитивная полоска и клеточные принципы построения амниотного тела посредством гаструляции» . Наука . 374 (6572): abg1727. дои : 10.1126/science.abg1727 . ПМИД 34855481 . S2CID 244841366 .
- ^ Jump up to: а б Микава Т., По А.М., Келли К.А., Исии Ю., Риз Д.Е. (2004). «Индукция и формирование паттерна первичной полоски, организующего центра гаструляции амниоты» . Дев Дин . 229 (3): 422–32. дои : 10.1002/dvdy.10458 . ПМИД 14991697 . S2CID 758473 .
- ^ Jump up to: а б Даунс К.М. (2009). «Загадочная примитивная полоса: преобладающие представления и проблемы, касающиеся оси тела млекопитающих» . Биоэссе . 31 (8): 892–902. doi : 10.1002/bies.200900038 . ПМЦ 2949267 . ПМИД 19609969 .
- ^ Чуай М., Цзэн В., Ян Х., Бойченко В., Стекольщик Дж.А., Вейер С.Дж. (2006). «Движение клеток во время формирования примитивной полоски цыпленка» . Дев. Биол . 296 (1): 137–49. дои : 10.1016/j.ydbio.2006.04.451 . ПМК 2556955 . ПМИД 16725136 .
- ^ Чуай М., Вейер С.Дж. (2008). «Механизмы, лежащие в основе формирования примитивных полосок у куриного эмбриона». Актуальные темы биологии развития . Том. 81. стр. 135–56. дои : 10.1016/S0070-2153(07)81004-0 . ISBN 978-0-12-374253-7 . ПМИД 18023726 .
- ^ Грейпин-Боттон, А.; Констам, Д. (2007). «Эволюция механизмов и молекулярный контроль формирования эндодермы». Механизмы развития . 124 (4): 253–78. дои : 10.1016/j.mod.2007.01.001 . ПМИД 17307341 . S2CID 16552755 .
- ^ Jump up to: а б с Тернер, Дэвид А.; Рю, По; Маккензи, Джонатан П.; Дэвис, Элеонора; Мартинес Ариас, Альфонсо (1 января 2014 г.). «Brachyury взаимодействует с передачей сигналов Wnt/β-catenin, вызывая поведение, подобное примитивным полоскам, при дифференцировке эмбриональных стволовых клеток мыши» . БМК Биология . 12:63 . дои : 10.1186/s12915-014-0063-7 . ISSN 1741-7007 . ПМЦ 4171571 . ПМИД 25115237 .
- ^ Jump up to: а б Вармфлэш, Арье; Сорре, Бенуа; Эток, Фред; Сиггиа, Эрик Д; Бриванлу, Али Х (2014). «Метод повторения раннего эмбрионального пространственного паттерна в эмбриональных стволовых клетках человека» . Природные методы . 11 (8): 847–854. дои : 10.1038/nmeth.3016 . ПМК 4341966 . ПМИД 24973948 .
- ^ Jump up to: а б Эток, Фред; Мецгер, Якоб; Рузо, Альберт; Кирст, Кристоф; Йони, Анна; Озаир, М. Зишан; Бриванлу, Али Х.; Сиггиа, Эрик Д. (2016). «Баланс между секретируемыми ингибиторами и контролем краевой чувствительности гаструлоидной самоорганизации» . Развивающая клетка . 39 (3): 302–315. дои : 10.1016/j.devcel.2016.09.016 . ПМК 5113147 . ПМИД 27746044 .
- ^ Jump up to: а б Бринк, Сюзанна К. ван ден; Бэйли-Джонсон, Питер; Балайо, Тина; Хаджантонакис, Анна-Катерина; Новочин, Соня; Тернер, Дэвид А.; Ариас, Альфонсо Мартинес (15 ноября 2014 г.). «Нарушение симметрии, спецификация зародышевого листка и аксиальная организация в агрегатах эмбриональных стволовых клеток мыши» . Разработка . 141 (22): 4231–4242. дои : 10.1242/dev.113001 . ISSN 0950-1991 . ПМК 4302915 . ПМИД 25371360 .
- ^ Jump up to: а б с Тернер, Дэвид Эндрю; Глодовски, Шериз Р.; Луз, Алонсо-Кризостомо; Бэйли-Джонсон, Питер; Хейворд, Пенни С.; Коллиньон, Жером; Густавсен, Карстен; Серуп, Палле; Шретер, Кристиан (13 мая 2016 г.). «Взаимодействие между передачей сигналов Nodal и Wnt обеспечивает надежное нарушение симметрии и аксиальную организацию в гаструлоидах (эмбриональных органоидах)». bioRxiv 10.1101/051722 .
- ^ Jump up to: а б с Тернер, Дэвид; Алонсо-Кризостомо, Луз; Гиргин, Мехмет; Бэйли-Джонсон, Питер; Глодовски, Шериз Р.; Хейворд, Пенелопа К.; Коллиньон, Жером; Густавсен, Карстен; Серуп, Палле (31 января 2017 г.). «У гаструлоидов развиваются три оси тела при отсутствии внеэмбриональных тканей и пространственно локализованной передачи сигналов». bioRxiv 10.1101/104539 .
- ^ Беккари, Леонардо; Морис, Наоми; Гиргин, Мехмет; Тернер, Дэвид А.; Бэйли-Джонсон, Питер; Косси, Анна-Катрин; Лютольф, Матиас П.; Дюбул, Денис; Ариас, Альфонсо Мартинес (октябрь 2018 г.). «Свойства многоосной самоорганизации эмбриональных стволовых клеток мыши в гаструлоиды» . Природа . 562 (7726): 272–276. Бибкод : 2018Natur.562..272B . дои : 10.1038/s41586-018-0578-0 . ISSN 0028-0836 . ПМИД 30283134 . S2CID 52915553 .
- ^ Тернер, Дэвид А.; Гиргин, Мехмет; Алонсо-Кризостомо, Луз; Триведи, Викас; Бэйли-Джонсон, Питер; Глодовски, Шериз Р.; Хейворд, Пенелопа К.; Коллиньон, Жером; Густавсен, Карстен (01 ноября 2017 г.). «Переднезадняя полярность и удлинение при отсутствии внеэмбриональных тканей и пространственно-локализованной передачи сигналов в гаструлоидах: эмбриональные органоиды млекопитающих» . Разработка . 144 (21): 3894–3906. дои : 10.1242/dev.150391 . ISSN 0950-1991 . ПМК 5702072 . ПМИД 28951435 .
- ^ «Экстракорпоральное оплодотворение (ЭКО) – Клиника Майо» . www.mayoclinic.org . Проверено 11 апреля 2022 г.
- ^ Асплунд, Кьелл (2020). «Использование экстракорпорального оплодотворения — этические проблемы» . Упсальский журнал медицинских наук . 125 (2): 192–199. дои : 10.1080/03009734.2019.1684405 . ISSN 2000-1967 . ПМК 7721055 . PMID 31686575 . S2CID 207896932 .
- ^ Дэвис, Кейтлин (01 марта 2019 г.). «Границы исследований эмбрионов: расширение правила четырнадцати дней». Журнал биоэтических исследований . 16 (1): 133–140. дои : 10.1007/s11673-018-09895-w . ISSN 1872-4353 . ПМИД 30635823 . S2CID 58643344 .
Библиография
[ редактировать ]- Арнольд, Себастьян Дж.; Робертсон, Элизабет Дж. (2009). «Принятие на себя обязательств: распределение клеточных линий и формирование осевого рисунка у раннего эмбриона мыши». Нат. Преподобный мол. Клеточная Биол. 10 (2): 91–103. дои : 10.1038/nrm2618 . ПМИД 19129791 . S2CID 94174 .

- Катала, Мартин (2005). «Эмбриология позвоночника и спинного мозга» . В Тортори-Донати, Паоло; и др. (ред.). Детская нейрорадиология: мозг . Спрингер. ISBN 978-3-540-41077-5 .
- Эресковский, Александр В. (2010). Сравнительная эмбриология губок . Спрингер. ISBN 978-90-481-8574-0 .
- Гилберт, Скотт Ф. (2010). Биология развития (Девятое изд.). Синауэр Ассошиэйтс. ISBN 978-0-87893-558-1 .
- Холл, Брайан Кейт (1998). «8.3.3 Гаструла и гаструляция» . Эволюционная биология развития (2-е изд.). Нидерланды: Kluwer Academic Publishers. ISBN 978-0-412-78580-1 .
- Харрисон, Лайонел Г. (2011). Формирование жизни: создание биологической модели . Издательство Кембриджского университета. ISBN 978-0-521-55350-6 .
- Макгиди, Томас А., изд. (2006). «Гаструляция». Ветеринарная эмбриология . Уайли-Блэквелл. ISBN 978-1-4051-1147-8 .
- Мундлос, Стефан (2009). «Действие генов: генетика развития». В Спейчере, Майкл; и др. (ред.). Генетика человека Фогеля и Мотульского: проблемы и подходы (4-е изд.). Спрингер. дои : 10.1007/978-3-540-37654-5 . ISBN 978-3-540-37653-8 .
- Тэм, Патрик П.Л.; Берингер, Ричард Р. (1997). «Гаструляция мыши: формирование плана тела млекопитающих» . Мех. Дев. 68 (1–2): 3–25. дои : 10.1016/S0925-4773(97)00123-8 . ПМИД 9431800 . S2CID 14052942 .

Дальнейшее чтение
[ редактировать ]- Барон, Маргарет Х. (2001). «Эмбриональная индукция кроветворения и васкулогенеза млекопитающих» . В Зоне, Леонард И. (ред.). Гематопоэз: подход к развитию . Издательство Оксфордского университета. ISBN 978-0-19-512450-7 .
- Каллен, К.Э. (2009). «эмбриология и раннее развитие животных» . Энциклопедия наук о жизни, Том 2 . Информационная база. ISBN 978-0-8160-7008-4 .
- Форгач, Г.; Ньюман, Стюарт А. (2005). «Дробление и образование бластулы» . Биологическая физика развивающегося эмбриона . Издательство Кембриджского университета. Бибкод : 2005bpde.book.....F . ISBN 978-0-521-78337-8 .
- Форгач, Г.; Ньюман, Стюарт А. (2005). «Эпителиальный морфогенез: гаструляция и нейруляция» . Биологическая физика развивающегося эмбриона . Издательство Кембриджского университета. Бибкод : 2005bpde.book.....F . ISBN 978-0-521-78337-8 .
- Харт, Натан Х.; Флак, Ричард А. (1995). «Эпиболия и гаструляция» . В Capco, Дэвид (ред.). Цитоскелетные механизмы в развитии животных . Академическая пресса. ISBN 978-0-12-153131-7 .
- Кнуст, Элизабет (1999). «Гаструляционные движения» . В Бирчмайере, Уолтер; Бирхмайер, Кармен (ред.). Эпителиальный морфогенез в развитии и заболеваниях . ЦРК Пресс. стр. 152–153. ISBN 978-90-5702-419-1 .
- Кунц, Иветт В. (2004). «Гаструляция» . Биология развития костистых рыб . Спрингер. ISBN 978-1-4020-2996-7 .
- Нация, Джеймс Л., изд. (2009). «Гаструляция» . Физиология и биохимия насекомых . ЦРК Пресс. ISBN 978-0-8493-1181-9 .
- Росс, Лоуренс М.; Ламперти, Эдвард Д., ред. (2006). «Онтогенез человека: гаструляция, нейруляция и формирование сомитов» . Атлас анатомии: общая анатомия и опорно-двигательный аппарат . Тиме. ISBN 978-3-13-142081-7 .
- Санес, Дэн Х.; и др. (2006). «Ранняя эмбриология многоклеточных животных» . Развитие нервной системы (2-е изд.). Академическая пресса. стр. 1–2. ISBN 978-0-12-618621-5 .
- Стангер, Бен З.; Мелтон, Дуглас А. (2004). «Развитие энтодермальных производных в легких, печени, поджелудочной железе и кишечнике» . В Эпштейне, Чарльз Дж.; и др. (ред.). Врожденные ошибки развития: молекулярные основы клинических нарушений морфогенеза . Издательство Оксфордского университета. ISBN 978-0-19-514502-1 .
Внешние ссылки
[ редактировать ]- Анимация гаструляции. Архивировано 20 октября 2015 г. на Wayback Machine.
- Иллюстрации и фильмы о гаструляции из книги «Гаструляция: от клеток к эмбриону» под редакцией Клаудио Стерна.
- Видео гаструляции лягушки
| Базы данных органов управления : Национальные |
|---|