Ретиноевая кислота
 | |
 | |
| Имена | |
|---|---|
| Название ИЮПАК Ретиноевая кислота | |
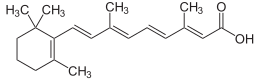
| Систематическое название ИЮПАК (2 E ,4 E ,6 E ,8 E )-3,7-Диметил-9-(2,6,6-триметилциклогекс-1-ен-1-ил)нона-2,4,6,8-тетраеновая кислота кислота | |
| Другие имена витамин А кислота; РА | |
| Идентификаторы | |
3D model ( JSmol ) | |
| ЧЭБИ | |
| ЧЕМБЛ | |
| ХимическийПаук | |
ПабХим CID | |
| НЕКОТОРЫЙ | |
| Характеристики | |
| С 20 Н 28 О 2 | |
| Молярная масса | 300.43512 g/mol |
| Появление | Кристаллический порошок от желтого до светло-оранжевого цвета с характерным цветочным запахом. [1] |
| Температура плавления | От 180 до 182 ° C (от 356 до 360 ° F; от 453 до 455 К) кристаллы этанола [1] |
| почти нерастворимый | |
| Растворимость в жире | растворимый |
| Родственные соединения | |
Родственные соединения | ретинол ; сетчатка ; бета-каротин |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
Ретиноевая кислота (упрощенно используемая здесь для обозначения полностью транс -ретиноевой кислоты) представляет собой метаболит витамина А 1 (полностью транс - ретинола ), который опосредует функции витамина А 1, необходимые для роста и развития. Полностью транс -ретиноевая кислота необходима хордовым животным, включая всех высших животных от рыб до человека. Во время раннего эмбрионального развития полностью транс -ретиноевая кислота, вырабатываемая в определенной области эмбриона, помогает определять положение вдоль передней/задней оси эмбриона, служа межклеточной сигнальной молекулой, которая направляет развитие задней части эмбриона. [2] Он действует через Hox-гены , которые в конечном итоге контролируют формирование переднего/заднего паттерна на ранних стадиях развития. [3]
Полностью транс -ретиноевая кислота (ATRA) является основной ретиноевой кислотой, в то время как изомеры, такие как 13- цис- и 9- цис -ретиноевая кислота, также присутствуют в гораздо меньших количествах. [4]
Ключевая роль транс - ретиноевой кислоты в эмбриональном развитии обуславливает высокую тератогенность ретиноидных фармацевтических препаратов, таких как изотретиноин (13- цис -ретиноевая кислота), используемый для лечения рака и прыщей . Пероральные мегадозы предварительно полученного витамина А ( ретинилпальмитата ) и самой полностью транс -ретиноевой кислоты также обладают тератогенным потенциалом по тому же механизму.
Механизм биологического действия [ править ]
Полностью транс -ретиноевая кислота действует путем связывания с рецептором ретиноевой кислоты (RAR), который связан с ДНК в виде гетеродимера с рецептором ретиноида X (RXR) в областях, называемых элементами ответа на ретиноевую кислоту (RARE). Связывание лиганда all -trans -ретиноевой кислоты с RAR изменяет конформацию RAR, что влияет на связывание других белков, которые либо индуцируют, либо подавляют транскрипцию близлежащего гена (включая Hox-гены и некоторые другие гены-мишени). RAR опосредуют транскрипцию различных наборов генов, контролирующих дифференцировку различных типов клеток, таким образом, регулируемые гены-мишени зависят от клеток-мишеней. [5] В некоторых клетках одним из генов-мишеней является ген самого рецептора ретиноевой кислоты ( RAR-бета у млекопитающих), который усиливает ответ. [6] Контроль уровня ретиноевой кислоты поддерживается набором белков, которые контролируют синтез и деградацию ретиноевой кислоты. [2] [3]
Молекулярная основа взаимодействия полностью транс -ретиноевой кислоты с Hox-генами была изучена с помощью делеционного анализа на трансгенных мышах, несущих конструкции репортерных генов GFP . Такие исследования выявили функциональные RARE во фланкирующих последовательностях некоторых из большинства 3'-генов Hox (включая HOXA1 , HOXB1 , HOXB4 , HOXD4 ), что указывает на прямое взаимодействие между генами и ретиноевой кислотой. Исследования такого типа убедительно подтверждают нормальную роль ретиноидов в формировании паттерна эмбриогенеза позвоночных посредством Hox-генов. [7]
Биосинтез [ править ]
Полностью транс- ретиноевая кислота может вырабатываться в организме в результате двух последовательных стадий окисления, которые превращают полностью транс -ретинол в ретинальдегид и в полностью транс -ретиноевую кислоту, но после образования она не может быть снова восстановлена до полностью транс -ретинола. Ферменты, которые генерируют ретиноевую кислоту для регуляции экспрессии генов, включают ретинолдегидрогеназу (Rdh10), которая метаболизирует ретинол в ретинальдегид, и три типа ретинальдегиддегидрогеназы , а именно ALDH1A1 (RALDH1), ALDH1A2 (RALDH2) и ALDH1A3 (RALDH3). [8] которые метаболизируют ретинальдегид до ретиноевой кислоты. [2] Ферменты, которые метаболизируют избыток транс -ретинола для предотвращения токсичности, включают алкогольдегидрогеназу и цитохром P450 ( CYP26 ). [9]
Функция при отсутствии предшественников [ править ]
Полностью -транс -ретиноевая кислота отвечает за большую часть активности витамина А 1 , за исключением эффектов зрительного пигмента, для которых требуется ретиналь (ретинальдегид), и эффектов клеточного метаболизма, для которых может потребоваться ретинол сам . Кроме того, некоторые биохимические функции, необходимые для фертильности у самцов и самок млекопитающих с дефицитом витамина А, изначально требовали полностью транс -ретинола для спасения, но это связано с необходимостью локального превращения полностью транс -ретинола в полностью транс -ретиноат. кислота, поскольку при введении полностью транс -ретиноевая кислота не достигает некоторых критически важных тканей, если ее не вводить в больших количествах. Таким образом, если животных кормят только полностью транс -ретиноевой кислотой, но не содержат витамина А 1 (полностью транс -ретинола или ретиналя), они не страдают от эффектов задержки роста или повреждения эпителия, вызванных недостатком витамина А 1 (включая отсутствие витамина А 1). ксерофтальмия — сухость роговицы). Они страдают дегенерацией сетчатки и слепотой из-за дефицита сетчатки.
Кроме того, у крыс-самцов, лишенных витамина А 1, но получавших полностью транс -ретиноевую кислоту, наблюдается гипогонадизм и бесплодие из-за отсутствия локального синтеза ретиноевой кислоты в семенниках; подобное лечение самок крыс вызывает бесплодие вследствие внутриутробной резорбции, вызванной отсутствием локального синтеза ретиноевой кислоты в эмбрионе. [10] [11] Синтез ретиноевой кислоты в семенниках катализируется преимущественно альдегиддегидрогеназой ALDH1A2 (RALDH2). Подавление этого фермента было предложено как возможный способ изготовления противозачаточных таблеток для мужчин, поскольку ретиноевая кислота необходима для сперматогенеза у людей, как и у крыс. [12]
в эмбриональном развитии Функция
Полностью транс -ретиноевая кислота (ATRA) представляет собой сигнальную молекулу морфогена , что означает, что она зависит от концентрации; пороки развития могут возникнуть при избытке или недостатке концентрации ATRA. Другими молекулами, которые взаимодействуют с ATRA, являются гены FGF8 , Cdx и Hox, которые участвуют в развитии различных структур внутри эмбриона. Например, ATRA играет важную роль в активации Hox-генов, необходимых для развития заднего мозга . Задний мозг, который позже дифференцируется в ствол мозга , служит основным сигнальным центром, определяющим границу головы и туловища. [13] Двусторонний градиент ретиноевой кислоты, высокий в туловище и низкий в месте соединения с головой и хвостом, репрессирует FGF8 в развивающемся туловище, обеспечивая нормальный сомитогенез , передних конечностей инициацию зачатков и формирование предсердий в сердце . [14] При воздействии избытка ATRA задний мозг увеличивается, препятствуя росту других частей мозга; другие аномалии развития, которые могут возникнуть при избытке ATRA, — это отсутствие или слияние сомитов , а также проблемы с аортой и крупными сосудами сердца. При накоплении этих пороков развития у человека может быть диагностирован синдром ДиДжорджа . [15] Однако, поскольку ATRA участвует в различных процессах развития, аномалии, связанные с потерей ATRA, не ограничиваются только участками, связанными с синдромом ДиДжорджа. Ретиноевая кислота необходима на протяжении всей жизни человека, но наиболее важна во время беременности. Без надлежащих концентраций ATRA могут возникнуть серьезные отклонения и даже фатальные для растущего плода. Исследования генетической потери функции у эмбрионов мышей и рыбок данио, которые устраняют синтез ATRA или рецепторы ATRA (RAR), выявили аномальное развитие сомитов, зачатков передних конечностей, сердца, заднего мозга, спинного мозга, глаз, переднего мозга базальных ганглиев , почек, энтодермы передней кишки. , и т. д. [14]
Сопутствующие фармацевтические препараты [ править ]
- Таларозол
- Третиноин / полностью транс -ретиноевая кислота (торговое название: Retin-A)
- Изотретиноин /13- цис -ретиноевая кислота (торговое название: Аккутан (США), Роаккутан)
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б Индекс Merck , 13-е издание, 8251 .
- ↑ Перейти обратно: Перейти обратно: а б с Дустер Дж. (сентябрь 2008 г.). «Синтез ретиноевой кислоты и передача сигналов на раннем органогенезе» . Клетка . 134 (6): 921–31. дои : 10.1016/j.cell.2008.09.002 . ПМК 2632951 . ПМИД 18805086 .
- ↑ Перейти обратно: Перейти обратно: а б Голландия LZ (май 2007 г.). «Биология развития: хордовые с отличием» . Природа . 447 (7141): 153–5. Бибкод : 2007Natur.447..153H . дои : 10.1038/447153а . ПМИД 17495912 . S2CID 5549210 .
- ^ Рюль Р., Крезель В., де Лера А.Р. (декабрь 2018 г.). «9-цис-13,14-дигидроретиноевая кислота, новый эндогенный лиганд ретиноидного рецептора X млекопитающих и активный лиганд потенциально новой категории витамина А: витамина А5» . Обзоры питания . 76 (12): 929–941. дои : 10.1093/nutrit/nuy057 . ПМИД 30358857 .
- ^ Венкатеш К., Шрикант Л., Венгамма Б., Чандрасекхар С., Сандживкумар А., Мулешвара Прасад Б.К., Сарма П.В. (2013). «Дифференциация in vitro культивируемых клеток CD34+ человека в астроциты» . Неврология Индия . 61 (4): 383–8. дои : 10.4103/0028-3886.117615 . ПМИД 24005729 .
- ^ Вингендер Э (1993). «Рецепторы стероидных/тиреоидных гормонов». Регуляция генов у эукариот . Нью-Йорк: ВЧ. п. 316. ИСБН 1-56081-706-2 .
- ^ Маршалл Х., Моррисон А., Студер М., Попперл Х., Крумлауф Р. (1996). «Ретиноиды и Hox-гены» . Журнал ФАСЭБ . 10 (9): 969–978. дои : 10.1096/fasebj.10.9.8801179 . ПМИД 8801179 . S2CID 16062049 .
- ^ «Семья АЛДХ 1» . Лаборатория доктора Василиса Василиу в Центре медицинских наук Университета Колорадо. Архивировано из оригинала 13 января 2013 года . Проверено 22 октября 2012 г.
- ^ Молотков А, Гизелинк Н.Б., Шамбон П., Дустер Г (октябрь 2004 г.). «Противоположные действия клеточного ретинол-связывающего белка и алкогольдегидрогеназы контролируют баланс между хранением и распадом ретинола» . Биохимический журнал . 383 (Часть 2): 295–302. дои : 10.1042/BJ20040621 . ПМК 1134070 . ПМИД 15193143 .
- ^ Мур Т., Холмс PD (октябрь 1971 г.). «Выработка экспериментального дефицита витамина А у крыс и мышей». Лабораторные животные . 5 (2): 239–50. дои : 10.1258/002367771781006492 . ПМИД 5126333 . S2CID 34221571 .
- ^ ван Пелт А.М., де Рой Д.Г. (февраль 1991 г.). «Ретиноевая кислота способна повторно инициировать сперматогенез у крыс с дефицитом витамина А, а высокие повторные дозы поддерживают полное развитие сперматогенных клеток». Эндокринология . 128 (2): 697–704. дои : 10.1210/эндо-128-2-697 . ПМИД 1989855 .
- ^ Кин С. (октябрь 2012 г.). «Исследование контрацепции. Новое изобретение таблетки: мужской контроль над рождаемостью». Наука . 338 (6105): 318–20. Бибкод : 2012Sci...338..318K . дои : 10.1126/science.338.6105.318 . ПМИД 23087225 .
- ^ Ли К., Скромне I (ноябрь 2014 г.). «Ретиноевая кислота регулирует размер, структуру и расположение тканей на границе головы и туловища» . Разработка . 141 (22): 4375–84. дои : 10.1242/dev.109603 . ПМИД 25371368 .
- ↑ Перейти обратно: Перейти обратно: а б Каннингем Т.Дж., Дустер Дж. (февраль 2015 г.). «Механизмы передачи сигналов ретиноевой кислоты и ее роль в развитии органов и конечностей» . Обзоры природы. Молекулярно-клеточная биология . 16 (2): 110–23. дои : 10.1038/nrm3932 . ПМК 4636111 . ПМИД 25560970 .
- ^ Ринн М., Долле П. (март 2012 г.). «Передача сигналов ретиноевой кислоты во время развития» . Разработка . 139 (5): 843–58. дои : 10.1242/dev.065938 . ПМИД 22318625 .