Каротиноид

Каротиноиды ( / k ə ˈ r ɒ t ɪ n ɔɪ d / ) — желтые, оранжевые и красные органические пигменты , которые производятся растениями и водорослями , а также некоторыми бактериями, архей и грибами . [1] Каротиноиды придают характерный цвет тыквам , моркови , пастернаку , кукурузе , помидорам , канареекам , фламинго , лососю , омарам , креветкам и нарциссам . Более 1100 идентифицированных каротиноидов можно разделить на два класса — ксантофиллы (содержащие кислород) и каротины (которые представляют собой чисто углеводороды и не содержат кислорода). [2]
Все они являются , что производными тетратерпенов означает, что они производятся из 8 единиц изопрена и содержат 40 атомов углерода. В целом каротиноиды поглощают волны длиной от 400 до 550 нанометров (от фиолетового до зеленого света). Это приводит к тому, что соединения приобретают насыщенный желтый, оранжевый или красный цвет. Каротиноиды являются доминирующим пигментом осенней окраски листьев примерно у 15-30% древесных пород. [3] но многие цвета растений, особенно красные и пурпурные, обусловлены полифенолами .

Каротиноиды выполняют две ключевые роли в растениях и водорослях: они поглощают энергию света для использования в фотосинтезе и обеспечивают фотозащиту посредством нефотохимического тушения . [4] Каротиноиды, которые содержат незамещенные бета-иононовые кольца (включая β-каротин , α-каротин , β-криптоксантин и γ-каротин ), обладают активностью витамина А (это означает, что они могут превращаться в ретинол ). В глазах лютеин , мезо -зеаксантин и зеаксантин присутствуют в виде макулярных пигментов , значение которых для зрительной функции по состоянию на 2016 год остается в стадии клинических исследований . [3] [5]
Структура и функции
[ редактировать ]


Каротиноиды производятся всеми фотосинтезирующими организмами и в основном используются в качестве пигментов хлорофилла вспомогательных в светособирающей части фотосинтеза.
Они сильно ненасыщены сопряженными двойными связями , что позволяет каротиноидам поглощать свет различных длин волн . В то же время концевые группы регулируют полярность и свойства липидных мембран .
Большинство каротиноидов являются тетратерпеноидами , регулярными изопреноиды . Существует несколько модификаций этих структур: включая циклизацию , различную степень насыщения или ненасыщенности и другие функциональные группы . [6] Каротины обычно содержат только углерод и водород, т. е. являются углеводородами . Выдающиеся члены включают α-каротин , β-каротин и ликопин , известные как каротины . Каротиноиды, содержащие кислород, включают лютеин и зеаксантин . Они известны как ксантофиллы . [3] Их цвет, варьирующийся от бледно-желтого через ярко-оранжевый до темно-красного, напрямую связан с их строением, особенно с длиной спряжения. [3] Ксантофиллы часто имеют желтый цвет, что дает название их класса.

Каротиноиды также участвуют в различных типах клеточной передачи сигналов. [7] Они способны сигнализировать о выработке абсцизовой кислоты , которая регулирует рост растений, покой семян , созревание и прорастание эмбрионов , деление и удлинение клеток, рост цветков и реакции на стресс. [8]
Фотофизика
[ редактировать ]Длина кратных сопряженных двойных связей определяет их цвет и фотофизику. [9] [10] После поглощения фотона каротиноид передает свой возбужденный электрон хлорофиллу для использования в фотосинтезе. [9] При поглощении света каротиноиды передают энергию возбуждения хлорофиллу и обратно . Синглет-синглетный перенос энергии представляет собой перенос состояний с более низкой энергией и используется во время фотосинтеза. [7] Триплет-триплетный перенос представляет собой более высокое энергетическое состояние и имеет важное значение для фотозащиты. [7] Свет производит вредные виды во время фотосинтеза, причем наиболее разрушительными являются активные формы кислорода (АФК). Поскольку эти высокоэнергетические АФК производятся в хлорофилле, энергия передается полиеновому хвосту каротиноида и подвергается серии реакций, в которых электроны перемещаются между каротиноидными связями, чтобы найти наиболее сбалансированное (наименьшее энергетическое) состояние для каротиноида. [9]
Каротиноиды защищают растения от синглетного кислорода как путем передачи энергии, так и посредством химических реакций. Они также защищают растения, подавляя триплет хлорофилла. [11] Защищая липиды от повреждения свободными радикалами, которые генерируют заряженные пероксиды липидов и другие окисленные производные, каротиноиды поддерживают кристаллическую архитектуру и гидрофобность липопротеинов и клеточных липидных структур, следовательно, растворимость кислорода и его диффузию в них. [12]
Отношения структура-свойство
[ редактировать ]Как и некоторые жирные кислоты , каротиноиды липофильны из-за наличия длинных ненасыщенных алифатических цепей. [3] Как следствие, каротиноиды обычно присутствуют в липопротеинах плазмы и клеточных липидных структурах. [13]
Морфология
[ редактировать ]Каротиноиды расположены преимущественно вне ядра клетки , в различных органеллах цитоплазмы, липидных каплях , цитосомах и гранулах. Они были визуализированы и количественно оценены с помощью рамановской спектроскопии в клетке водоросли . [14]
разработкой моноклональных антител к трансликопину С удалось локализовать этот каротиноид в различных клетках животных и человека. [15]

Продукты питания
[ редактировать ]Бета-каротин , содержащийся в тыкве , сладком картофеле , моркови и тыкве , отвечает за их оранжево-желтый цвет. [3] Сушеная морковь содержит самое большое количество каротина среди всех продуктов питания на 100-граммовую порцию, измеряемую в эквивалентах активности ретинола (эквивалентах провитамина А). [3] [16] Вьетнамские плоды гака содержат самую высокую известную концентрацию каротиноида ликопина . [17] Хотя зелень, капуста , шпинат , листовая капуста и зелень репы содержат значительное количество бета-каротина. [3] Рацион фламинго богат каротиноидами, придающими перьям этих птиц оранжевый цвет. [18]
Обзоры предварительных исследований 2015 года показали, что продукты с высоким содержанием каротиноидов могут снизить риск развития рака головы и шеи. [19] и рак простаты . [20] Нет никакой корреляции между потреблением продуктов с высоким содержанием каротиноидов и витамина А и риском болезни Паркинсона . [21]
Люди и другие животные по большей части не способны синтезировать каротиноиды и должны получать их с пищей. Каротиноиды являются обычным и часто декоративным свойством животных. Например, розовый цвет лосося , а также красная окраска приготовленных омаров и чешуя желтой морфы обыкновенных стенных ящериц обусловлены каротиноидами. [22] [ нужна ссылка ] Было предложено использовать каротиноиды в декоративных признаках (крайние примеры см. Тупики ), поскольку, учитывая их физиологические и химические свойства, они могут использоваться в качестве видимых индикаторов здоровья особи и, следовательно, используются животными при выборе потенциальных партнеров. [23]
Каротиноиды, поступающие с пищей, сохраняются в жировых тканях животных. [3] и исключительно плотоядные животные получают соединения из животного жира. В рационе человека всасывание каротиноидов улучшается при употреблении их с жиром во время еды. [24] Приготовление содержащих каротиноиды овощей в масле и измельчение овощей увеличивают биодоступность каротиноидов . [3] [24] [25]
Цвета растений
[ редактировать ]

Наиболее распространенные каротиноиды включают ликопин и предшественник витамина А β-каротин. В растениях ксантофилл лютеин является наиболее распространенным каротиноидом, и его роль в предотвращении возрастных заболеваний глаз в настоящее время изучается. [5] Лютеин и другие каротиноидные пигменты, обнаруженные в зрелых листьях, часто не заметны из-за маскирующего присутствия хлорофилла . Когда хлорофилла нет, как в осенней листве, преобладают желтые и оранжевые каротиноиды. По той же причине каротиноиды часто преобладают в спелых фруктах после того, как их демаскирует исчезновение хлорофилла.
Каротиноиды ответственны за ярко-желтый и оранжевый цвет лиственной листвы (например, умирающих осенних листьев ) некоторых видов лиственных пород, таких как гикори , ясень , клен , желтый тополь , осина , береза , черешня , платан , тополь , сассафрас и ольха . Каротиноиды являются доминирующим пигментом осенней окраски листьев примерно у 15-30% древесных пород. [26] Однако красные, пурпурные и их смешанные комбинации, украшающие осеннюю листву, обычно происходят из другой группы пигментов в клетках, называемых антоцианами . В отличие от каротиноидов, эти пигменты не присутствуют в листе на протяжении всего вегетационного периода, но активно вырабатываются к концу лета. [27]
Цвет птиц и половой отбор
[ редактировать ]Диетические каротиноиды и их метаболические производные ответственны за окраску птиц от ярко-желтой до красной. [28] По оценкам исследований, около 2956 современных видов птиц обладают каротиноидной окраской и что способность использовать эти пигменты для внешней окраски развивалась независимо много раз на протяжении всей истории эволюции птиц. [29] Окраска каротиноидов демонстрирует высокий уровень полового диморфизма : взрослые самцы птиц обычно имеют более яркую окраску, чем самки того же вида. [30]
Эти различия возникают из-за выбора желтой и красной окраски у самцов по предпочтениям самки . [31] [30] У многих видов птиц самки тратят больше времени и ресурсов на воспитание потомства, чем их партнеры-самцы. Поэтому крайне важно, чтобы самки птиц тщательно отбирали качественных партнеров. Современная литература поддерживает теорию о том, что яркая окраска каротиноидов коррелирует с мужскими качествами – либо через прямое воздействие на иммунную функцию и окислительный стресс, либо через прямое воздействие на иммунную функцию и окислительный стресс. [32] [33] [34] или через связь между путями метаболизма каротиноидов и путями клеточного дыхания. [35] [36]
Обычно считается, что признаки, выбранные половым путем, такие как окраска на основе каротиноидов, развиваются, потому что они являются честными сигналами фенотипического и генетического качества. Например, среди самцов птиц вида Parus major более ярко украшенные самцы производят сперму, которая лучше защищена от окислительного стресса из-за повышенного присутствия каротиноидных антиоксидантов . [37] Однако есть также свидетельства того, что привлекательная окраска самцов может быть ложным сигналом о мужских качествах. Среди колюшек самцы, которые более привлекательны для самок из-за каротиноидных красителей, по-видимому, недостаточно распределяют каротиноиды в своих зародышевых клетках. [38] Поскольку каротиноиды являются полезными антиоксидантами, их недостаточное распределение в зародышевых клетках может привести к усилению окислительного повреждения ДНК этих клеток. [38] Поэтому самки колюшки могут рисковать плодовитостью и жизнеспособностью своего потомства, выбирая более красных, но более испорченных партнеров с пониженным качеством спермы .
Ароматические химикаты
[ редактировать ]Продукты деградации каротиноидов, такие как иононы , дамасконы и дамасценоны, также являются важными ароматическими химическими веществами, которые широко используются в парфюмерной и парфюмерной промышленности. И β-дамасценон, и β-ионон, хотя их концентрация в дистиллятах розы низкая , являются основными соединениями, обуславливающими запах цветов. Фактически, сладкие цветочные запахи, присутствующие в черном чае , выдержанном табаке , винограде и многих фруктах , обусловлены ароматическими соединениями, образующимися в результате распада каротиноидов.
Болезнь
[ редактировать ]Некоторые каротиноиды производятся бактериями для защиты от окислительной иммунной атаки. Пигмент aureus (золотой), который дал название некоторым штаммам Staphylococcus aureus, представляет собой каротиноид, называемый стафилоксантин . Этот каротиноид является фактором вирулентности с антиоксидантным действием, который помогает микробу избежать смерти от активных форм кислорода, используемых иммунной системой хозяина. [39]
Биосинтез
[ редактировать ]
Основными строительными блоками каротиноидов являются изопентенилдифосфат (IPP) и диметилаллилдифосфат (DMAPP). [40] Эти два изомера изопрена используются для создания различных соединений в зависимости от биологического пути, используемого для синтеза изомеров. [41] Известно, что растения используют два разных пути производства IPP: цитозольный путь мевалоновой кислоты (MVA) и пластидный метилэритрит-4-фосфат (MEP). [40] У животных производство холестерина начинается с создания IPP и DMAPP с использованием MVA. [41] На предприятиях по производству каротиноидов MEP используется для получения IPP и DMAPP. [40] Путь MEP приводит к образованию смеси IPP:DMAPP 5:1. [41] IPP и DMAPP подвергаются нескольким реакциям, в результате которых образуется основной предшественник каротиноидов — геранилгеранилдифосфат (GGPP). GGPP может превращаться в каротины или ксантофиллы, проходя ряд различных стадий пути биосинтеза каротиноидов. [40]
Путь MEP
[ редактировать ]Глицеральдегид-3-фосфат и пируват , промежуточные продукты фотосинтеза , превращаются в дезокси-D-ксилулозо-5-фосфат (DXP), катализируемый DXP-синтазой (DXS). DXP-редуктоизомераза катализирует восстановление НАДФН и последующую перегруппировку. [40] [41] Полученный MEP превращается в 4-(цитидин-5'-дифосфо)-2-C-метил-D-эритрит (CDP-ME) в присутствии CTP с использованием фермента MEP цитидилилтрансферазы. Затем CDP-ME превращается в присутствии АТФ в 2-фосфо-4-(цитидин-5'-дифосфо)-2-C-метил-D-эритрит (CDP-ME2P). Превращение в CDP-ME2P катализируется киназой CDP-ME . Затем CDP-ME2P превращается в 2-C-метил-D-эритрит-2,4-циклодифосфат (MECDP). Эта реакция происходит, когда MECDP-синтаза катализирует реакцию и CMP удаляется из молекулы CDP-ME2P. Затем MECDP превращается в (е)-4-гидрокси-3-метилбут-2-ен-1-илдифосфат (HMBDP) посредством HMBDP-синтазы в присутствии флаводоксина и НАДФН. HMBDP восстанавливается до IPP в присутствии ферредоксина и НАДФН ферментом HMBDP-редуктазой . Последние два этапа с участием HMBPD-синтазы и редуктазы могут происходить только в полностью анаэробной среде. Затем IPP может изомеризоваться в DMAPP с помощью изомеразы IPP. [41]
Путь биосинтеза каротиноидов
[ редактировать ]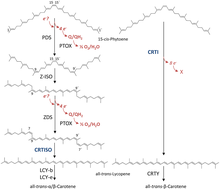
Две молекулы GGPP конденсируются посредством фитоенсинтазы PSY), образуя 15-цис- изомер фитоена ( . PSY принадлежит к семейству сквален/фитоенсинтаз и гомологичен скваленсинтазе , которая принимает участие в биосинтезе стероидов . Последующее превращение фитоена в полностью трансликопин зависит от организма. единственный фермент — бактериальную фитоендесатуразу Бактерии и грибы используют для катализа (CRTI). Однако растения и цианобактерии используют для этого процесса четыре фермента. [42] Первым из этих ферментов является фитоендесатураза растительного типа , которая вводит две дополнительные двойные связи в 15-цис-фитоен путем дегидрирования и изомеризует две из его существующих двойных связей из транс в цис, образуя 9,15,9'-три-цис-фитоен. ζ-каротин. Центральная двойная связь этого три-цис-ζ-каротина изомеризуется зета -каротин-изомеразой Z-ISO, и образующийся 9,9'-ди-цис-ζ-каротин снова дегидрируется с помощью ζ-каротиндесатуразы (ZDS). ) . Это снова приводит к образованию двух двойных связей, в результате чего образуется 7,9,7',9'-тетра-цис-ликопин. CRTISO , каротиноидизомераза, необходима для превращения цис -ликопина в полностью транс -ликопин в присутствии восстановленного FAD .
Этот полностью транс-ликопин циклизован; циклизация приводит к появлению разнообразия каротиноидов, которые можно отличить по концевым группам. Это может быть либо бета-кольцо , либо эпсилон-кольцо, каждое из которых генерируется разными ферментами ( ликопин-бета-циклаза [бета-LCY] или ликопин-эпсилон-циклаза [эпсилон-LCY]). α-каротин образуется, когда полностью транс-ликопин сначала подвергается реакции с эпсилон-LCY, а затем второй реакции с бета-LCY; тогда как β-каротин образуется в результате двух реакций с бета-LCY. α- и β-каротины являются наиболее распространенными каротиноидами в фотосистемах растений , но их все же можно в дальнейшем превращать в ксантофиллы с помощью бета-гидролазы и эпсилон-гидролазы, что приводит к образованию множества ксантофиллов. [40]
Регулирование
[ редактировать ]Считается, что и DXS, и DXR являются ферментами, определяющими скорость, что позволяет им регулировать уровень каротиноидов. [40] Это было обнаружено в эксперименте, в котором DXS и DXR были генетически сверхэкспрессированы, что привело к увеличению экспрессии каротиноидов в полученных проростках. [40] Кроме того, считается, что шапероны J-белка (J20) и белка теплового шока 70 (Hsp70) участвуют в посттранскрипционной регуляции активности DXS, так что мутанты с дефектной активностью J20 демонстрируют пониженную активность фермента DXS при накоплении неактивного белка DXS. [43] Регуляция также может быть вызвана внешними токсинами , которые влияют на ферменты и белки, необходимые для синтеза. Кетокломазон получают из гербицидов , вносимых в почву, и связывается с DXP-синтазой. [41] Это ингибирует DXP-синтазу, предотвращая синтез DXP и останавливая путь MEP. [41] Использование этого токсина приводит к снижению уровня каротиноидов в растениях, выращиваемых в загрязненной почве. [41] Фосмидомицин , антибиотик , является конкурентным ингибитором редуктоизомеразы DXP из-за его структуры, сходной с ферментом. [41] Применение указанного антибиотика предотвращает снижение уровня DXP, снова останавливая путь MEP. [41]
Каротиноиды природного происхождения
[ редактировать ]- Углеводороды
- Ликоперсен 7,8,11,12,15,7',8',11',12',15'-Декагидро-γ,γ-каротин
- Фитофлуен
- Ликопин
- Гексагидроликопин 15- цис -7,8,11,12,7',8'-Гексагидро-γ,γ-каротин
- Торулен 3',4'-дидегидро-β,γ-каротин
- α-зеакаротин 7',8'-дигидро-ε,γ-каротин
- α-каротины
- β-каротины
- γ-каротины
- δ-каротин
- е-каротин
- ζ-каротины
- Спирты
- Аллоксантин
- Бактериоруберин 2,2'-Бис(3-гидрокси-3-метилбутил)-3,4,3',4'-тетрадегидро-1,2,1',2'-тетрагидро-γ,γ-каротин-1,1 '-диол
- Синтиаксантин
- Пектеноксантин
- Криптомонаксантин (3R,3'R)-7,8,7',8'-тетрадегидро-β,β-каротин-3,3'-диол
- Крустаксантин β,-каротин-3,4,3',4'-тетрол
- Газаниаксантин (3R)-5'-цис-β,γ-каротин-3-ол
- OH-Хлоробактен 1',2'-Дигидро-f,γ-каротин-1'-ол
- Лороксантин β,ε-каротин-3,19,3'-триол
- Лютеин (3R,3'R,6'R)-β,ε-каротин-3,3'-диол
- Ликоксантин γ,γ-каротин-16-ол
- Родопин 1,2-дигидро-γ,γ-каротин-1-ол
- Родопинол, он же Вармингол 13- цис -1,2-дигидро-γ,γ-каротин-1,20-диол
- Сапроксантин 3',4'-дидегидро-1',2'-дигидро-β,γ-каротин-3,1'-диол
- Зеаксантин
- Гликозиды
- Осциллаксантин 2,2'-Бис(β-L-рамнопираносилокси)-3,4,3',4'-тетрадегидро-1,2,1',2'-тетрагидро-γ,γ-каротин-1,1'- диол
- Флейксантофилл 1'-(β-D-глюкопиранозилокси)-3',4'-дидегидро-1',2'-дигидро-β,γ-каротин-2'-ол
- Эфиры
- Родовибрин 1'-Метокси-3',4'-дидегидро-1,2,1',2'-тетрагидро-γ,γ-каротин-1-ол
- Сфероиден- 1-метокси-3,4-дидегидро-1,2,7',8'-тетрагидро-γ,γ-каротин
- Эпоксиды
- Диадиноксантин 5,6-эпокси-7',8'-дидегидро-5,6-дигидрокаротин-3,3-диол
- Лютеоксантин 5,6: 5',8'-Диэпокси-5,6,5',8'-тетрагидро-β,β-каротин-3,3'-диол
- Мутатоксантин
- Цитроксантин
- Зеаксантин фураноксид 5,8-эпокси-5,8-дигидро-β,β-каротин-3,3’-диол
- Неохром 5',8'-Эпокси-6,7-дидегидро-5,6,5',8'-тетрагидро-β,β-каротин-3,5,3'-триол
- Фолиахром
- Троллихром
- Ваухериаксантин 5',6'-Эпокси-6,7-дидегидро-5,6,5',6'-тетрагидро-β,β-каротин-3,5,19,3'-тетрол
- Альдегиды
- Родопинал
- Вармингон 13-цис-1-гидрокси-1,2-дигидро-γ,γ-каротин-20-ал
- Торулародинальдегид 3',4'-дидегидро-β,γ-каротин-16'-аль
- Кислоты и эфиры кислот
- Торуларходин 3',4'-дидегидро-β,γ-каротин-16'-овая кислота
- Метиловый эфир торуларходина Метил 3',4'-дидегидро-β,γ-каротин-16'-оат
- Кетоны
- Астасен
- Астаксантин
- Кантаксантин [44] он же афаницин, хлореллаксантин β,β-каротин-4,4'-дион
- Капсантин (3R,3'S,5'R)-3,3'-Дигидрокси-β,κ-каротин-6'-он
- Капсорубин (3S,5R,3'S,5'R)-3,3'-Дигидрокси-κ,κ-каротин-6,6'-дион
- Криптокапсин (3'R,5'R)-3'-гидрокси-β,κ-каротин-6'-он
- 2,2'-Дикетоспириллоксантин 1,1'-Диметокси-3,4,3',4'-тетрадегидро-1,2,1',2'-тетрагидро-γ,γ-каротин-2,2'-дион
- Эхиненон β,β-каротин-4-он
- 3'-Гидроксихиненон
- Флексиксантин 3,1'-дигидрокси-3',4'-дидегидро-1',2'-дигидро-β,γ-каротин-4-он
- 3-ОН-кантаксантин, он же адонирубин, он же феникоксантин 3-гидрокси-β,β-каротин-4,4’-дион
- Гидроксисфериоденон 1'-Гидрокси-1-метокси-3,4-дидегидро-1,2,1',2',7',8'-гексагидро-γ,γ-каротин-2-он
- Окенон 1'-Метокси-1',2'-дигидро-с,γ-каротин-4'-он
- Пектенолон 3,3'-Дигидрокси-7',8'-дидегидро-β,β-каротин-4-он
- Фениконон, он же дегидроадонирубин 3-гидрокси-2,3-дидегидро-β,β-каротин-4,4’-дион
- Феникоптерон β,ε-каротин-4-он
- Рубиксантон 3-гидрокси-β,γ-каротин-4’-он
- Сифонаксантин 3,19,3'-тригидрокси-7,8-дигидро-β,ε-каротин-8-он
- Эфиры спиртов
- Астацеин 3,3'-биспальмитоилокси-2,3,2',3'-тетрадегидро-β,β-каротин-4,4'-дион или 3,3'-дигидрокси-2,3,2',3'- тетрадегидро-β,β-каротин-4,4'-дион дипальмитат
- Фукоксантин 3'-Ацетокси-5,6-эпокси-3,5'-дигидрокси-6',7'-дидегидро-5,6,7,8,5',6'-гексагидро-β,β-каротин-8 -один
- Изофукоксантин 3'-Ацетокси-3,5,5'-тригидрокси-6',7'-дидегидро-5,8,5',6'-тетрагидро-β,β-каротин-8-он
- Физалиен
- Сифонеин 3,3'-дигидрокси-19-лауроилокси-7,8-дигидро-β,ε-каротин-8-он или 3,19,3’-тригидрокси-7,8-дигидро-β,ε-каротин-8 -один 19-лауреат
- Апокаротиноиды
- β-Апо-2'-каротин 3',4'-дидегидро-2'-апо-b-каротин-2'-ал
- Апо-2-ликопенал
- Апо-6'-ликопенал 6'-Апо-y-каротин-6'-ал
- Азафринальдегид 5,6-дигидрокси-5,6-дигидро-10'-апо-β-каротин-10'-ал
- Биксин 6'-Метилводород 9'-цис-6,6'-диапокаротин-6,6'-диоат
- Цитранаксантин 5',6'-Дигидро-5'-апо-β-каротин-6'-он или 5',6'-дигидро-5'-апо-18'-нор-β-каротин-6'-он или 6'-метил-6'-апо-β-каротин-6'-он
- Кроцетин 8,8'-Диапо-8,8'-каротиндиовая кислота
- Кроцетинсемиальдегид 8'-оксо-8,8'-диапо-8-каротиновая кислота
- Кроцин Дигентиобиозил 8,8'-диапо-8,8'-каротендиоат
- Хопкинсиаксантин 3-гидрокси-7,8-дидегидро-7',8'-дигидро-7'-апо-b-каротин-4,8'-дион или 3-гидрокси-8'-метил-7,8-дидегидро- 8'-апо-b-каротин-4,8'-дион
- Метилапо-6'-ликопеноат Метил-6'-апо-у-каротин-6'-оат
- Парацентрон 3,5-дигидрокси-6,7-дидегидро-5,6,7',8'-тетрагидро-7'-апо-b-каротин-8'-он или 3,5-дигидрокси-8'-метил- 6,7-дидегидро-5,6-дигидро-8'-апо-b-каротин-8'-он
- Синтаксантин 7',8'-Дигидро-7'-апо-b-каротин-8'-он или 8'-метил-8'-апо-b-каротин-8'-он
- Нор- и секокаротиноиды
- Актиниоэритрин 3,3'-бисацилокси-2,2'-динор-b,b-каротин-4,4'-дион
- β-каротенон 5,6:5',6'-Дизеко-b,b-каротин-5,6,5',6'-тетрон
- Перидинин 3'-Ацетокси-5,6-эпокси-3,5'-дигидрокси-6',7'-дидегидро-5,6,5',6'-тетрагидро-12',13',20'-тринор- б,б-каротин-19,11-олид
- Пирроксантининол 5,6-эпокси-3,3'-дигидрокси-7',8'-дидегидро-5,6-дигидро-12',13',20'-тринор-b,b-каротин-19,11-олид
- Полу-α-каротинон 5,6-Секо-b,е-каротин-5,6-дион
- Полу-β-каротинон 5,6-секо-b,b-каротин-5,6-дион или 5',6'-секо-b,b-каротин-5',6'-дион
- Трифазиаксантин 3-Гидроксисеми-b-каротенон 3'-Гидрокси-5,6-секо-b,b-каротин-5,6-дион или 3-гидрокси-5',6'-секо-b,b-каротин-5 ',6'-дион
- Ретро-каротиноиды и ретро-апо-каротиноиды
- Эшшольцксантин 4',5'-дидегидро-4,5'-ретро-b,b-каротин-3,3'-диол
- Эшшольцксантон 3'-Гидрокси-4',5'-дидегидро-4,5'-ретро-b,b-каротин-3-он
- Родоксантин 4',5'-дидегидро-4,5'-ретро-b,b-каротин-3,3'-дион
- Тангераксантин 3-гидрокси-5'-метил-4,5'-ретро-5'-апо-b-каротин-5'-он или 3-гидрокси-4,5'-ретро-5'-апо-b-каротин -5'-один
- Высшие каротиноиды
- Нонапреноксантин 2-(4-гидрокси-3-метил-2-бутенил)-7',8',11',12'-тетрагидро-е,y-каротин
- Декапреноксантин 2,2'-бис(4-гидрокси-3-метил-2-бутенил)-е,е-каротин
- Cp 450 2-[4-Гидрокси-3-(гидроксиметил)-2-бутенил]-2'-(3-метил-2-бутенил)-b,b-каротин
- Cp 473 2'-(4-Гидрокси-3-метил-2-бутенил)-2-(3-метил-2-бутенил)-3',4'-дидегидро-1',2'-дигидро-β,γ -каротин-1'-ол
- Бактериоруберин 2,2'-Бис(3-гидрокси-3-метилбутил)-3,4,3',4'-тетрадегидро-1,2,1',2'-тетрагидро-γ,γ-каротин-1,1 '-диол
См. также
[ редактировать ]- Список фитохимических веществ в продуктах питания
- CRT (генетика) — кластер генов, отвечающий за биосинтез каротиноидов.
- Номер E#E100–E199 (цвета)
- Фитохимия
Ссылки
[ редактировать ]- ^ Нельсон, Дэвид Л.; Кокс, Майкл М. (2005). Принципы биохимии (4-е изд.). Нью-Йорк: WH Freeman. ISBN 0-7167-4339-6 .
- ^ Ябузаки, Джунко (01 января 2017 г.). «База данных каротиноидов: структуры, химические признаки и распространение среди организмов» . База данных . 2017 (1). дои : 10.1093/база данных/bax004 . ПМК 5574413 . ПМИД 28365725 .
- ^ Перейти обратно: а б с д и ж г час я дж «Каротиноиды» . Информационный центр по микроэлементам, Институт Лайнуса Полинга, Университет штата Орегон. 1 августа 2016 года . Проверено 17 апреля 2019 г.
- ^ Армстронг Г.А., Херст Дж.Э. (1996). «Каротиноиды 2: Генетика и молекулярная биология биосинтеза каротиноидных пигментов» . ФАСЕБ Дж . 10 (2): 228–37. дои : 10.1096/fasebj.10.2.8641556 . ПМИД 8641556 . S2CID 22385652 .
- ^ Перейти обратно: а б Бернштейн, П.С.; Ли, Б; Вачали, ПП; Горусупуди, А; Шьям, Р; Хенриксен, бакалавр наук; Нолан, Дж. М. (2015). «Лютеин, зеаксантин и мезо-зеаксантин: фундаментальная и клиническая наука, лежащая в основе диетических вмешательств на основе каротиноидов против глазных заболеваний» . Прогресс в исследованиях сетчатки и глаз . 50 : 34–66. doi : 10.1016/j.preteyeres.2015.10.003 . ПМЦ 4698241 . ПМИД 26541886 .
- ^ Мареска, Джулия А.; Ромбергер, Стивен П.; Брайант, Дональд А. (28 мая 2008 г.). «Биосинтез изорениератена в зеленых серных бактериях требует совместного действия двух каротиноидциклаз» . Журнал бактериологии . 190 (19): 6384–6391. дои : 10.1128/JB.00758-08 . ISSN 0021-9193 . ПМК 2565998 . ПМИД 18676669 .
- ^ Перейти обратно: а б с Когделл, Р.Дж. (30 ноября 1978 г.). «Каротиноиды в фотосинтезе». Фил. Пер. Р. Сок. Лонд. Б. 284 (1002): 569–579. Бибкод : 1978RSPTB.284..569C . дои : 10.1098/rstb.1978.0090 . ISSN 0080-4622 .
- ^ Финкельштейн, Рут (1 ноября 2013 г.). «Синтез и реакция абсцизовой кислоты» . Книга «Арабидопсис» . 11 : e0166. дои : 10.1199/tab.0166 . ISSN 1543-8120 . ПМК 3833200 . ПМИД 24273463 .
- ^ Перейти обратно: а б с Вершинин, Александр (1 января 1999 г.). «Биологические функции каротиноидов - разнообразие и эволюция». Биофакторы . 10 (2–3): 99–104. дои : 10.1002/biof.5520100203 . ISSN 1872-8081 . ПМИД 10609869 . S2CID 24408277 .
- ^ Поливка, Томаш; Сундстрем, Вилли (2004). «Сверхбыстрая динамика каротиноидных возбужденных состояний - от решения к природным и искусственным системам». Химические обзоры . 104 (4): 2021–2072. дои : 10.1021/cr020674n . ПМИД 15080720 .
- ^ Рамель, Фанни; Биртик, Симона; Куине, Стефан; Триантафилидес, Кристиан; Раванат, Жан-Люк; Хаво, Мишель (2012). «Химическое тушение синглетного кислорода каротиноидами в растениях» . Физиология растений . 158 (3): 1267–1278. дои : 10.1104/стр.111.182394 . ПМК 3291260 . ПМИД 22234998 .
- ^ Джон Томас Ландрам (2010). Каротиноиды: физические, химические и биологические функции и свойства . Бока-Ратон: CRC Press. ISBN 978-1-4200-5230-5 . OCLC 148650411 .
- ^ Грушецкий, Веслав И. (2004), Франк, Гарри А.; Янг, Эндрю Дж.; Бриттон, Джордж; Когделл, Ричард Дж. (ред.), «Каротиноиды в мембранах» , «Фотохимия каротиноидов» , «Достижения в области фотосинтеза и дыхания», том. 8, Дордрехт: Kluwer Academic Publishers, стр. 363–379, doi : 10.1007/0-306-48209-6_20 , ISBN. 978-0-7923-5942-5 , получено 28 марта 2021 г.
- ^ Тимлин, Джерилин А.; Коллинз, Аарон М.; Бичем, Томас А.; Шумская, Мария; Вурцель, Элеонора Т. (14 июня 2017 г.), «Локализация и количественная оценка каротиноидов в интактных клетках и тканях», Carotenoids , InTech, doi : 10.5772/68101 , ISBN 978-953-51-3211-0 , S2CID 54807067
- ^ Петяев Иван М.; Зигангирова Наиля А.; Пристенский Дмитрий; и др. (2018). «Неинвазивная иммунофлуоресцентная оценка статуса добавок ликопина в мазках кожи» . Моноклональные антитела в иммунодиагностике и иммунотерапии . 37 (3): 139–146. дои : 10.1089/mab.2018.0012 . ISSN 2167-9436 . ПМИД 29901405 . S2CID 49190846 .
- ^ «Продукты с самым высоким эквивалентом активности ретинола» . Nutritiondata.self.com . Проверено 4 декабря 2015 г.
- ^ Тран, ХТ; Паркс, ЮВ; Роуч, полиция; Голдинг, Дж. Б.; Нгуен, Миннесота (2015). «Влияние зрелости на физико-химические свойства плодов Gac (Momordica cochinchinensis Spreng.)» . Пищевая наука и питание . 4 (2): 305–314. дои : 10.1002/fsn3.291 . ПМЦ 4779482 . ПМИД 27004120 .
- ^ Йим, К.Дж.; Квон, Дж; Ча, ИТ; Ох, КС; Песня, HS; Ли, Х.В.; Ри, Дж. К.; Сонг, Э.Дж.; Ро, младший; Со, МЛ; Чой, Дж.С.; Чой, HJ; Ли, С.Дж.; Нам, Ю.Д.; Ро, SW (2015). «Появление жизнеспособных краснопигментированных галоархей в оперении содержащихся в неволе фламинго» . Научные отчеты . 5 : 16425. Бибкод : 2015NatSR...516425Y . дои : 10.1038/srep16425 . ПМЦ 4639753 . ПМИД 26553382 .
- ^ Леончини; Источники, природные; Голова; Рак, Шея; и др. (июль 2015 г.). «Систематический обзор и метаанализ эпидемиологических исследований». Биомаркеры эпидемиологии рака Пред . 24 (7): 1003–11. doi : 10.1158/1055-9965.EPI-15-0053 . ПМИД 25873578 . S2CID 21131127 .
- ^ Соарес Нда, К; и др. (октябрь 2015 г.). «Противораковые свойства каротиноидов при раке простаты. Обзор» (PDF) . Гистол Гистопатол . 30 (10): 1143–54. дои : 10.14670/HH-11-635 . ПМИД 26058846 .
- ^ Такеда, А; и др. (2014). «Витамин А и каротиноиды и риск болезни Паркинсона: систематический обзор и метаанализ» . Нейроэпидемиология . 42 (1): 25–38. дои : 10.1159/000355849 . ПМИД 24356061 . S2CID 12396064 .
- ^ Сакки, Роберто (4 июня 2013 г.). «Цветовые вариации полиморфной настенной ящерицы (Podarcis muralis): анализ с использованием цветовой системы RGB». Зоологический обзор . 252 (4): 431–439. дои : 10.1016/j.jcz.2013.03.001 .
- ^ Уайтхед Р.Д., Озакинчи Дж., Перретт Д.И. (2012). «Привлекательный цвет кожи: использование полового отбора для улучшения питания и здоровья» . Эвол Психол . 10 (5): 842–54. дои : 10.1177/147470491201000507 . ПМЦ 10429994 . ПМИД 23253790 . S2CID 8655801 .
- ^ Перейти обратно: а б Машурабад, Пурна Чандра; Палика, Равиндранад; Жирва, Иветт Вильда; Бхаскарачари, К.; Пуллахандам, Рагху (3 января 2017 г.). «Состав пищевых жиров, пищевая матрица и относительная полярность модулируют мицелляризацию и всасывание в кишечнике каротиноидов из овощей и фруктов» . Журнал пищевой науки и технологий . 54 (2): 333–341. дои : 10.1007/s13197-016-2466-7 . ISSN 0022-1155 . ПМК 5306026 . ПМИД 28242932 .
- ^ Родриго, Мария Хесус; Силла, Антонио; Барбера, Рейес; Закариас, Лоренцо (2015). «Биодоступность каротиноидов в мякоти и свежем соке богатых каротиноидами сладких апельсинов и мандаринов». Еда и функции . 6 (6): 1950–1959. дои : 10.1039/c5fo00258c . ПМИД 25996796 .
- ^ Арчетти, Марко; Деринг, Томас Ф.; Хаген, Снорре Б.; Хьюз, Николь М.; Кожа, Саймон Р.; Ли, Дэвид В.; Лев-Ядун, Симха; Манетас, Яннис; Огам, Хелен Дж. (2011). «Разгадка эволюции осенних цветов: междисциплинарный подход». Тенденции в экологии и эволюции . 24 (3): 166–73. дои : 10.1016/j.tree.2008.10.006 . ПМИД 19178979 .
- ^ Дэвис, Кевин М., изд. (2004). Растительные пигменты и манипуляции с ними . Ежегодные обзоры растений. Том. 14. Оксфорд: Издательство Блэквелл. п. 6. ISBN 978-1-4051-1737-1 .
- ^ Дели, Каспар; Питерс, Энн (16 ноября 2016 г.). «Влияние механизмов цветообразования на половой дихроматизм оперения у воробьиных и попугаев» . Функциональная экология . 31 (4): 903–914. дои : 10.1111/1365-2435.12796 . ISSN 0269-8463 .
- ^ Томас, Дэниел Б.; МакГроу, Кевин Дж.; Батлер, Майкл В.; Каррано, Мэтью Т.; Мэдден, Одиль; Джеймс, Хелен Ф. (7 августа 2014 г.). «Древнее происхождение и многократное появление перьев, пигментированных каротиноидами, у птиц» . Труды Королевского общества B: Биологические науки . 281 (1788): 20140806. doi : 10.1098/rspb.2014.0806 . ISSN 0962-8452 . ПМК 4083795 . ПМИД 24966316 .
- ^ Перейти обратно: а б Куни, Кристофер Р.; Варлей, Зои К.; Нури, Лара О.; Муди, Кристофер Дж.А.; Джардин, Майкл Д.; Томас, Гэвин Х. (16 апреля 2019 г.). «Половой отбор предсказывает скорость и направление цветового расхождения в большом птичьем излучении» . Природные коммуникации . 10 (1): 1773. Бибкод : 2019NatCo..10.1773C . дои : 10.1038/s41467-019-09859-7 . ISSN 2041-1723 . ПМК 6467902 . ПМИД 30992444 .
- ^ Хилл, Джеффри Э. (сентябрь 1990 г.). «Самки домашних зябликов предпочитают ярких самцов: половой отбор по признаку, зависящему от состояния» . Поведение животных . 40 (3): 563–572. дои : 10.1016/s0003-3472(05)80537-8 . ISSN 0003-3472 . S2CID 53176725 .
- ^ Уивер, Райан Дж.; Сантос, Эдуардо С.А.; Такер, Анна М.; Уилсон, Алан Э.; Хилл, Джеффри Э. (08 января 2018 г.). «Метаболизм каротиноидов усиливает связь между окраской перьев и индивидуальными качествами» . Природные коммуникации . 9 (1): 73. Бибкод : 2018NatCo...9...73W . дои : 10.1038/s41467-017-02649-z . ISSN 2041-1723 . ПМЦ 5758789 . ПМИД 29311592 .
- ^ Саймонс, Мирре Дж. П.; Коэн, Алан А.; Ферхюльст, Саймон (14 августа 2012 г.). «О чем говорит каротиноид-зависимая окраска? Уровень каротиноидов в плазме сигнализирует об иммунокомпетентности и окислительном стрессе у птиц - метаанализ» . ПЛОС ОДИН . 7 (8): e43088. Бибкод : 2012PLoSO...743088S . дои : 10.1371/journal.pone.0043088 . ISSN 1932-6203 . ПМК 3419220 . ПМИД 22905205 .
- ^ Кох, Ребекка Э.; Хилл, Джеффри Э. (14 мая 2018 г.). «Требуют ли украшения на основе каротиноидов ресурсный компромисс? Оценка теории и данных» . Функциональная экология . 32 (8): 1908–1920. дои : 10.1111/1365-2435.13122 . ISSN 0269-8463 .
- ^ Хилл, Джеффри Э.; Джонсон, Джеймс Д. (ноябрь 2012 г.). «Гипотеза о окислительно-восстановительном потенциале витамина А: биохимическая основа для честной передачи сигналов посредством каротиноидной пигментации» . Американский натуралист . 180 (5): Е127–Е150. дои : 10.1086/667861 . ISSN 0003-0147 . ПМИД 23070328 . S2CID 2013258 .
- ^ Пауэрс, Мэтью Дж; Хилл, Джеффри Э (3 мая 2021 г.). «Обзор и оценка гипотезы общего пути для поддержания честности сигнала при окраске на основе красных кетокаротиноидов» . Интегративная и сравнительная биология . 61 (5): 1811–1826. дои : 10.1093/icb/icab056 . ISSN 1540-7063 . ПМИД 33940618 .
- ^ Хельфенштейн, Фабрис; Лосдат, Сильвен; Мёллер, Андерс Папе; Блаунт, Джонатан Д.; Рихнер, Хайнц (февраль 2010 г.). «Сперма разноцветных самцов лучше защищена от окислительного стресса» . Экологические письма . 13 (2): 213–222. Бибкод : 2010EcolL..13..213H . дои : 10.1111/j.1461-0248.2009.01419.x . ISSN 1461-0248 . ПМИД 20059524 .
- ^ Перейти обратно: а б Ким, Син-Ён; Веландо, Альберто (январь 2020 г.). «Привлекательные самцы колюшек несут больше окислительных повреждений ДНК в соме и зародышевой линии» . Журнал эволюционной биологии . 33 (1): 121–126. дои : 10.1111/jeb.13552 . ISSN 1420-9101 . ПМИД 31610052 . S2CID 204702365 .
- ^ Лю Джи, Эссекс А., Бьюкенен Дж.Т. и др. (2005). «Золотой пигмент Staphylococcus aureus ухудшает уничтожение нейтрофилов и повышает вирулентность благодаря своей антиоксидантной активности» . Дж. Эксп. Мед . 202 (2): 209–15. дои : 10.1084/jem.20050846 . ПМК 2213009 . ПМИД 16009720 .
- ^ Перейти обратно: а б с д и ж г час Нисар, Назия; Ли, Ли; Лу, Шан; Кхин, Най Чи; Погсон, Барри Дж. (05 января 2015 г.). «Метаболизм каротиноидов у растений» . Молекулярный завод . Метаболизм растений и синтетическая биология. 8 (1): 68–82. дои : 10.1016/j.molp.2014.12.007 . ПМИД 25578273 . S2CID 26818009 .
- ^ Перейти обратно: а б с д и ж г час я дж КУЗУЯМА, Томохиса; СЭТО, Харуо (9 марта 2012 г.). «Два различных пути основных метаболических предшественников биосинтеза изопреноидов» . Труды Японской академии. Серия Б, Физические и биологические науки . 88 (3): 41–52. Бибкод : 2012PJAB...88...41K . дои : 10.2183/pjab.88.41 . ISSN 0386-2208 . ПМЦ 3365244 . ПМИД 22450534 .
- ^ Мойзе, Александр Р.; Аль-Бабили, Салим; Вурцель, Элеонора Т. (31 октября 2013 г.). «Механистические аспекты биосинтеза каротиноидов» . Химические обзоры . 114 (1): 164–93. дои : 10.1021/cr400106y . ПМЦ 3898671 . ПМИД 24175570 .
- ^ Нисар, Назия; Ли, Ли; Лу, Шан; ЧиХин, Нет; Погсон, Барри Дж. (5 января 2015 г.). «Метаболизм каротиноидов у растений» . Молекулярный завод . 8 (1): 68–82. дои : 10.1016/j.molp.2014.12.007 . ПМИД 25578273 . S2CID 26818009 .
- ^ Чхве, Сеён; Ку, Санго (2005). «Эффективный синтез кетокаротиноидов кантаксантина, астаксантина и астацена». Дж. Орг. Хим . 70 (8): 3328–3331. дои : 10.1021/jo050101l . ПМИД 15823009 .
Внешние ссылки
[ редактировать ]- Каротиноиды Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)