Ферредоксин
Ферредоксины (от латинского Ferrum : железо + окислительно-восстановительный потенциал , часто сокращенно «fd») представляют собой железо-серные белки , которые опосредуют перенос электронов в ряде метаболических реакций. Термин «ферредоксин» был придуман Д.С. Уортоном из компании DuPont Co. и применен к «железному белку», впервые выделенному в 1962 году Мортенсоном, Валентайном и Карнаханом из анаэробной бактерии Clostridium Pasteurianum . [1] [2]
Другой окислительно-восстановительный белок, выделенный из хлоропластов шпината , получил название «хлоропластный ферредоксин». [3] Ферредоксин хлоропластов участвует как в циклических, так и в нециклических фотофосфорилирования реакциях фотосинтеза . При нециклическом фотофосфорилировании ферредоксин является последним акцептором электронов, тем самым восстанавливая фермент НАДФ. + редуктаза. Он принимает электроны, образующиеся под действием солнечного света – возбуждает хлорофилл и передает их ферменту ферредоксину: НАДФ. + оксидоредуктаза ЕС 1.18.1.2 .
Ферредоксины представляют собой небольшие белки, содержащие атомы железа и серы, организованные в виде железо-серных кластеров . Эти биологические « конденсаторы » могут принимать или разряжать электроны, что приводит к изменению степени окисления атомов железа между +2 и +3. Таким образом, ферредоксин действует как агент переноса электронов в биологических окислительно-восстановительных реакциях.
Другие бионеорганические системы транспорта электронов включают рубредоксины , цитохромы , белки синей меди и структурно родственные белки Риске .
Ферредоксины можно классифицировать по природе железо-серных кластеров и сходству последовательностей.
Биоэнергетика ферредоксинов
[ редактировать ]Ферредоксины обычно осуществляют перенос одного электрона.
- Фд 0
бык + е − Фд −
красный

Однако некоторые бактериальные ферредоксины (типа 2[4Fe4S]) имеют два железо-серных кластера и могут осуществлять две реакции переноса электрона. В зависимости от последовательности белка два переноса могут иметь почти одинаковые потенциалы восстановления или могут существенно различаться. [4] [5]
- Фд 0
бык + е − Фд −
красный
- Фд −
красный + е − Фд 2−
красный
Ферредоксины являются одними из наиболее восстанавливающих биологических переносчиков электронов. Обычно они имеют потенциал средней точки -420 мВ. [6] Восстановительный потенциал вещества в клетке будет отличаться от его среднего потенциала в зависимости от концентрации его восстановленной и окисленной форм. Для одноэлектронной реакции потенциал изменяется примерно на 60 мВ при каждом изменении соотношения концентрации в десятикратной степени. Например, если пул ферредоксина уменьшен примерно на 95%, потенциал восстановления составит около -500 мВ. [7] Для сравнения, другие биологические реакции , основной биосинтетический восстановитель клетки, в основном имеют меньший восстановительный потенциал: например, НАДФН имеет клеточный окислительно-восстановительный потенциал -370 мВ ( E
0 = -320 мВ).
В зависимости от последовательности поддерживающих белков ферредоксины имеют потенциал восстановления примерно от -500 мВ. [6] [8] до -340 мВ. [9] В одной клетке может быть несколько типов ферредоксинов, каждый из которых настроен на оптимальное выполнение различных реакций. [10]
Снижение ферредоксина
[ редактировать ]Ферредоксины с высокой степенью восстановления восстанавливаются либо с помощью другого сильного восстановителя, либо с использованием какого-либо источника энергии для «перегона» электронов от менее восстанавливающих источников к ферредоксину. [11]
Прямое сокращение
[ редактировать ]Реакции, которые восстанавливают Fd, включают окисление альдегидов до кислот, таких как реакция глицеральдегида в глицерат (-580 мВ), реакция дегидрогеназы монооксида углерода (-520 мВ) и реакции 2-оксокислоты: Fd-оксидоредуктазы (-500 мВ). [12] [8] подобно реакции, осуществляемой пируватсинтазой . [7]
Снижение мембранного потенциала
[ редактировать ]Уровень ферредоксина также можно снизить с помощью НАДН (-320 мВ) или H.
2 (-414 мВ), но эти процессы связаны с потреблением мембранного потенциала для «перевода» электронов в более высокое энергетическое состояние. [6] Комплекс Rnf представляет собой широко распространенный мембранный белок бактерий, который обратимо переносит электроны между НАДН и ферредоксином при перекачке Na. +
или Ч +
ионы через мембрану. Хемиосмотический потенциал мембраны расходуется на неблагоприятное восстановление Fd.
бык от НАДН. Эта реакция является важным источником Fd. −
красный у многих автотрофных организмов. Если клетка растет на субстратах, обеспечивающих избыток Fd −
красный , комплекс Rnf может переносить эти электроны на НАД +
и сохранить полученную энергию в мембранном потенциале. [13] Энергетически преобразующие гидрогеназы (Ech) представляют собой семейство ферментов, которые обратимо связывают перенос электронов между Fd и H.
2 во время накачки H +
ионы через мембрану, чтобы уравновесить разницу в энергии. [14]
- Фд 0
окс + NaDH + Na +
снаружи Фд 2−
красный + НАД +
+ Уже +
внутри
- Фд 0
бык + Н
2 + Ч +
снаружи Фд 2−
красный + Н +
+ Ч +
внутри
Электронная бифуркация
[ редактировать ]Неблагоприятное восстановление Fd от менее восстанавливающего донора электронов может сочетаться одновременно с благоприятным восстановлением окислителя посредством реакции бифуркации электронов . [6] Примером реакции бифуркации электронов является генерация Fd −
красный для фиксации азота у некоторых аэробных диазотрофов . Обычно при окислительном фосфорилировании перенос электронов от НАДН к убихинону (Q) сопровождается зарядкой движущей силы протона. У Azotobacter энергия, выделяемая при переносе одного электрона от НАДН к Q, используется для одновременного ускорения переноса одного электрона от НАДН к Fd. [15] [16]
Прямое восстановление высокопотенциальных ферредоксинов
[ редактировать ]Некоторые ферредоксины обладают достаточно высоким окислительно-восстановительным потенциалом, поэтому их можно напрямую восстанавливать НАДФН. Одним из таких ферредоксинов является адреноксин (-274 мВ), который принимает участие в биосинтезе многих стероидов млекопитающих. [17] Ферредоксин Fd3 в корнях растений, восстанавливающий нитраты и сульфиты, имеет потенциал средней точки -337 мВ и также восстанавливается НАДФН. [10]
Fe 2 S 2 ферредоксины
[ редактировать ]| Домен связывания кластера железа и серы 2Fe-2S | |||
|---|---|---|---|
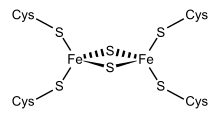 Структурное представление ферредоксина Fe 2 S 2 . | |||
| Идентификаторы | |||
| Символ | Фер2 | ||
| Пфам | PF00111 | ||
| Пфам Клан | CL0486 | ||
| ИнтерПро | IPR001041 | ||
| PROSITE | PDOC00642 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 3fxc / SCOPe / СУПФАМ | ||
| белок OPM | 1kf6 | ||
| |||
Члены суперсемейства ферредоксинов 2Fe–2S ( InterPro : IPR036010 ) имеют общую структуру ядра, состоящую из бета (2)-альфа-бета (2), которая включает путидаредоксин, терпредоксин и адренодоксин. [18] [19] [20] [21] Они представляют собой белки, состоящие примерно из ста аминокислот с четырьмя консервативными остатками цистеина, с которыми связан кластер 2Fe–2S. Эта консервативная область также встречается в виде домена в различных метаболических ферментах и в многодоменных белках, таких как альдегидоксидоредуктаза ( N -конец), ксантиноксидаза ( N -конец), фталатдиоксигеназаредуктаза ( С -конец), сукцинатдегидрогеназа железо-сера. белок ( N -концевой) и редуктаза метанмонооксигеназы ( N -концевой).
Ферредоксины растительного типа
[ редактировать ]Одну группу ферредоксинов, первоначально обнаруженную в мембранах хлоропластов , назвали «хлоропластным» или «растительным типом» ( InterPro : IPR010241 ). Его активным центром является кластер [Fe 2 S 2 ], в котором атомы железа тетраэдрически координированы как атомами неорганической серы, так и серами четырех консервативных остатков цистеина (Cys).
В хлоропластах ферредоксины Fe 2 S 2 функционируют как переносчики электронов в фотосинтетической электрон-транспортной цепи и как доноры электронов для различных клеточных белков, таких как глутаматсинтаза, нитритредуктаза, сульфитредуктаза и циклаза биосинтеза хлорофилла . [22] Поскольку циклаза является ферредоксин-зависимым ферментом, она может обеспечивать механизм координации между фотосинтезом и потребностью хлоропластов в хлорофилле путем связывания биосинтеза хлорофилла с фотосинтетической цепью переноса электронов. В гидроксилирующих бактериальных диоксигеназных системах они служат промежуточными переносчиками электронов между редуктазными флавопротеинами и оксигеназой.
Тиоредоксинподобные ферредоксины
[ редактировать ]Ферредоксин Fe 2 S 2 из Clostridium Pasteurianum ( Cp 2FeFd; P07324 ) был признан отдельным семейством белков на основании его аминокислотной последовательности, спектроскопических свойств его железо-серного кластера и уникальной способности двух цистеиновых лигандов заменять лиганды. кластер [Fe 2 S 2 ]. Хотя физиологическая роль этого ферредоксина остается неясной, сильное и специфическое взаимодействие Cp 2FeFd с молибден-железным белком нитрогеназой обнаружено гомологичные ферредоксины Azotobacter vinelandii ( Av 2FeFdI; P82802 ) и Aquifex aeolicus ( Aa Fd; O66511 . Охарактеризованы ). Кристаллическая структура Aa Fd решена. Aa Fd существует в виде димера. Строение мономера Aa Fd отличается от других ферредоксинов Fe 2 S 2 . Складка принадлежит к классу α+β, при этом первые четыре β-цепи и две α-спирали имеют вариант тиоредоксиновой складки . [23] UniProt классифицирует их как семейство «ферредоксинов типа 2Fe2S Шетны». [24]
Ферредоксины адренодоксинового типа
[ редактировать ]| ферредоксин 1 | |||
|---|---|---|---|
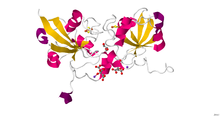 Кристаллическая структура человеческого ферредоксина-1 (FDX1). [25] | |||
| Идентификаторы | |||
| Символ | FDX1 | ||
| Альт. символы | ФДКС | ||
| ген NCBI | 2230 | ||
| HGNC | 3638 | ||
| МОЙ БОГ | 103260 | ||
| RefSeq | НМ_004109 | ||
| ЮниПрот | P10109 | ||
| Другие данные | |||
| Локус | Хр. 11 q22.3 | ||
| |||
Адренодоксин (надпочечниковый ферредоксин; InterPro : IPR001055 ), путидаредоксин и терпредоксин составляют семейство растворимых белков Fe 2 S 2 , которые действуют как переносчики одиночных электронов, в основном обнаруживаются в митохондриях эукариот и Pseudomonadota . Человеческий вариант адренодоксин называется ферредоксин-1 и ферредоксин-2 . В митохондриальных монооксигеназных системах адренодоксин переносит электрон от НАДФН:адренодоксинредуктазы к мембраносвязанному цитохрому P450 . У бактерий путидаредоксин и терпредоксин переносят электроны между соответствующими НАДН-зависимыми ферредоксинредуктазами и растворимыми P450. [26] [27] Точные функции других членов этого семейства неизвестны, хотя показано, что Escherichia coli Fdx участвует в биогенезе кластеров Fe–S. [28] Несмотря на низкое сходство последовательностей между ферредоксинами адренодоксинового и растительного типа, эти два класса имеют сходную топологию сворачивания.
Ферредоксин-1 у человека участвует в синтезе гормонов щитовидной железы. Он также переносит электроны от адренодоксинредуктазы к CYP11A1 , ферменту CYP450, ответственному за расщепление боковой цепи холестерина. FDX-1 обладает способностью связываться с металлами и белками. [29] Ферредоксин-2 участвует в синтезе гема А и железо-серного белка. [30]
Fe 4 S 4 и Fe 3 S 4 ферредоксины
[ редактировать ]Ферредоксины [Fe 4 S 4 ] можно далее подразделить на ферредоксины с низким потенциалом (бактериального типа) и ферредоксины с высоким потенциалом (HiPIP) .
Низко- и высокопотенциальные ферредоксины связаны следующей окислительно-восстановительной схемой:

Формальные степени окисления ионов железа могут быть [2Fe 3+ , 2Fe 2+ ] или [1Fe 3+ , 3Fe 2+ ] в низкопотенциальных ферредоксинах. Степени окисления ионов железа в высокопотенциальных ферредоксинах могут составлять [3Fe 3+ , 1Fe 2+ ] или [2Fe 3+ , 2Fe 2+ ].
Ферредоксины бактериального типа
[ редактировать ]| Связывающий домен 3Fe-4S | |||
|---|---|---|---|
 Структурное представление ферредоксина Fe 3 S 4 . | |||
| Идентификаторы | |||
| Символ | Фер4 | ||
| Пфам | PF00037 | ||
| ИнтерПро | ИПР001450 | ||
| PROSITE | PDOC00176 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 5фд1 / СКОПе / СУПФАМ | ||
| белок OPM | 1ккф | ||
| |||
Группа ферредоксинов Fe 4 S 4 , первоначально обнаруженная у бактерий, получила название «бактериального типа». Ферредоксины бактериального типа, в свою очередь, можно подразделить на дополнительные группы в зависимости от свойств их последовательности. Большинство из них содержат по крайней мере один консервативный домен, включая четыре остатка цистеина, которые связываются с кластером [Fe 4 S 4 ]. В ферредоксине Pyrococcus Furiosus Fe 4 S 4 один из консервативных остатков Cys заменен на аспарагиновую кислоту.
В ходе эволюции ферредоксинов бактериального типа происходили внутрипоследовательные дупликации генов, события транспозиции и слияния, что приводило к появлению белков с множественными железо-серными центрами. В некоторых бактериальных ферредоксинах один из дуплицированных доменов потерял один или несколько из четырех консервативных остатков Cys. Эти домены либо утратили способность связывать железо с серой, либо связываются с кластером [Fe 3 S 4 ] вместо кластера [Fe 4 S 4 ]. [31] и дикластерного типа. [32]
Трехмерные структуры известны для ряда монокластерных и дикластерных ферредоксинов бактериального типа. Складка принадлежит к классу α+β: 2–7 α-спиралей и четыре β-тяжи образуют бочкообразную структуру, а выдавленная петля содержит три «проксимальных» Cys-лиганда железо-серного кластера.
Высокопотенциальные железо-серные белки
[ редактировать ]Высокопотенциальные железо-серные белки (HiPIPs) образуют уникальное семейство Fe 4 S 4 ферредоксинов, которые функционируют в анаэробных электрон-транспортных цепях. Некоторые HiPIP имеют окислительно-восстановительный потенциал выше, чем любой другой известный железо-серный белок (например, HiPIP из Rhodopila globiformis имеет окислительно-восстановительный потенциал примерно -450 мВ). К настоящему времени структурно охарактеризованы несколько HiPIP, их складки относятся к классу α+β. Как и в других бактериальных ферредоксинах, единица [Fe 4 S 4 ] образует кластер кубанового типа и связана с белком через четыре остатка Cys.
Белки человека из семейства ферредоксинов
[ редактировать ]Ссылки
[ редактировать ]- ^ Мортенсон Л.Е., Валентайн Р.К., Карнахан Дж.Е. (июнь 1962 г.). «Фактор транспорта электронов Clostridium Pasteurianum». Связь с биохимическими и биофизическими исследованиями . 7 (6): 448–52. дои : 10.1016/0006-291X(62)90333-9 . ПМИД 14476372 .
- ^ Валентайн Р.К. (декабрь 1964 г.). «Бактериальный Ферредоксин» . Бактериологические обзоры . 28 (4): 497–517. дои : 10.1128/MMBR.28.4.497-517.1964 . ПМК 441251 . ПМИД 14244728 .
- ^ Тагава К., Арнон Д.И. (август 1962 г.). «Фередоксины как переносчики электронов при фотосинтезе, а также при биологическом производстве и потреблении газообразного водорода». Природа . 195 (4841): 537–43. Бибкод : 1962Natur.195..537T . дои : 10.1038/195537a0 . ПМИД 14039612 . S2CID 4213017 .
- ^ Майокко С.Дж., Арцинас А.Дж., Букер С.Дж., Эллиотт С.Дж. (январь 2019 г.). «Анализ окислительно-восстановительных потенциалов пяти ферредоксинов, обнаруженных в Thermotoga maritima» . Белковая наука . 28 (1): 257–266. дои : 10.1002/pro.3547 . ПМК 6295886 . ПМИД 30418685 .
- ^ Гао-Шеридан Х.С., Першад Х.Р., Армстронг Ф.А., Берджесс Б.К. (март 1998 г.). «Открытие нового ферредоксина из Azotobacter vinelandii, содержащего два кластера [4Fe-4S] с сильно различающимися и очень отрицательными восстановительными потенциалами» . Журнал биологической химии . 273 (10): 5514–9. дои : 10.1074/jbc.273.10.5514 . ПМИД 9488675 .
- ^ Перейти обратно: а б с д Бакель В., Тауэр Р.К. (2018). «Бифуркация электронов на основе флавина, ферредоксин, флаводоксин и анаэробное дыхание с протонами (Ech) или НАД». + (Rnf) как акцепторы электронов: исторический обзор» . Frontiers in Microbiology . 9 : 401. doi : 10.3389/ . PMC 5861303. . PMID 29593673 fmicb.2018.00401
- ^ Перейти обратно: а б Хувилер С.Г., Леффлер С., Ансельманн С.Е., Старк Х.Дж., фон Берген М., Флехслер Дж. и др. (февраль 2019 г.). «Металлоферментный комплекс массой один мегадальтон у Geobacter metallireducens, участвующий в восстановлении бензольного кольца за пределами биологического окислительно-восстановительного окна» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (6): 2259–2264. Бибкод : 2019PNAS..116.2259H . дои : 10.1073/pnas.1819636116 . ПМК 6369795 . ПМИД 30674680 .
- ^ Перейти обратно: а б Ли Б, Эллиот С.Дж. (2016). «Каталитическая ошибка 2-оксокислоты:ферредоксиноксидоредуктазы в CO2: эволюция и восстановление с помощью электрокаталитического анализа, опосредованного ферредоксином» . Электрохимика Акта . 199 : 349–356. дои : 10.1016/j.electacta.2016.02.119 .
- ^ Тамер В., Цирпус И., Ханс М., Пьерик А.Дж., Селмер Т., Билл Э. и др. (март 2003 г.). «Ферредоксин, содержащий два [4Fe-4S]-кластера, как альтернативный донор электронов для 2-гидроксиглутарил-КоА-дегидратазы из Acidaminococcus Fermentans». Архив микробиологии . 179 (3): 197–204. Бибкод : 2003ArMic.179..197T . дои : 10.1007/s00203-003-0517-8 . ПМИД 12610725 . S2CID 23621034 .
- ^ Перейти обратно: а б Ханке Г.Т., Кимата-Арига Ю., Танигучи И., Хасэ Т. (январь 2004 г.). «Постгеномная характеристика ферредоксинов Arabidopsis» . Физиология растений . 134 (1): 255–64. дои : 10.1104/стр.103.032755 . ПМК 316305 . ПМИД 14684843 .
- ^ Бойд Э.С., Аменабар М.Дж., Пудел С., Темплтон А.С. (февраль 2020 г.). «Биоэнергетические ограничения происхождения автотрофного метаболизма» . Философские труды. Серия А. Математические, физические и технические науки . 378 (2165): 20190151. Бибкод : 2020RSPTA.37890151B . дои : 10.1098/rsta.2019.0151 . ПМК 7015307 . ПМИД 31902344 .
- ^ Гибсон М.И., Чен П.И., Дреннан К.Л. (декабрь 2016 г.). «Структурная филогения для понимания функции оксидоредуктазы 2-оксокислот» . Современное мнение в области структурной биологии . 41 : 54–61. дои : 10.1016/j.sbi.2016.05.011 . ПМК 5381805 . ПМИД 27315560 .
- ^ Вестфаль Л., Вихманн А., Бейкер Дж., Минтон Н.П., Мюллер В. (ноябрь 2018 г.). «Комплекс Rnf представляет собой энергосвязанную трансгидрогеназу, необходимую для обратимого связывания клеточных пулов НАДН и ферредоксина в ацетогене Acetobacterium woodii» . Журнал бактериологии . 200 (21). дои : 10.1128/JB.00357-18 . ПМК 6182241 . ПМИД 30126940 .
- ^ Шельмерих MC, Мюллер В (апрель 2020 г.). «Энергопреобразующие гидрогеназы: связь между метаболизмом H 2 и сохранением энергии». Клеточные и молекулярные науки о жизни . 77 (8): 1461–1481. дои : 10.1007/s00018-019-03329-5 . ПМИД 31630229 . S2CID 204786346 .
- ^ Ледбеттер Р.Н., Гарсия Костас А.М., Лубнер К.Э., Малдер Д.В., Токмина-Лукашевска М., Арц Дж.Х. и др. (август 2017 г.). «Белковый комплекс FixABCX с раздвоением электронов из Azotobacter vinelandii: создание низкопотенциально восстанавливающих эквивалентов для нитрогеназного катализа» . Биохимия . 56 (32): 4177–4190. doi : 10.1021/acs.biochem.7b00389 . ПМЦ 7610252 . ПМИД 28704608 .
- ^ Пудель С., Колман Д.Р., Фиксен К.Р., Ледбеттер Р.Н., Чжэн Ю., Пенс Н. и др. (май 2018 г.). «Перенос электрона к нитрогеназе в различных геномных и метаболических условиях» . Журнал бактериологии . 200 (10). дои : 10.1128/JB.00757-17 . ПМЦ 5915786 . ПМИД 29483165 .
- ^ Юэн К.М., Рингл М., Бернхардт Р. (июнь 2012 г.). «Адренодоксин — универсальный ферредоксин» . ИУБМБ Жизнь . 64 (6): 506–12. дои : 10.1002/iub.1029 . ПМИД 22556163 .
- ^ Арменгауд Дж., Сайнс Г., Жуанно Ю., Зикер Л.С. (февраль 2001 г.). «Кристаллизация и предварительный рентгеноструктурный анализ [2Fe-2S] ферредоксина (FdVI) из Rhodobacter capsulatus». Акта Кристаллографика. Раздел D. Биологическая кристаллография . 57 (Часть 2): 301–3. дои : 10.1107/S0907444900017832 . ПМИД 11173487 .
- ^ Севрюкова И.Ф. (апрель 2005 г.). «Редокса-зависимая структурная реорганизация в путидаредоксине, ферредоксине [2Fe-2S] позвоночного типа из Pseudomonas putida». Журнал молекулярной биологии . 347 (3): 607–21. дои : 10.1016/j.jmb.2005.01.047 . ПМИД 15755454 .
- ^ Мо Х, Почапский С.С., Почапский Т.С. (апрель 1999 г.). «Модель структуры раствора окисленного терпредоксина, ферредоксина Fe2S2 из Pseudomonas». Биохимия . 38 (17): 5666–75. CiteSeerX 10.1.1.34.4745 . дои : 10.1021/bi983063r . ПМИД 10220356 .
- ^ Бейлке Д., Вайс Р., Лёр Ф., Пристовсек П., Ханнеманн Ф., Бернхардт Р. и др. (июнь 2002 г.). «Новый механизм транспорта электронов в митохондриальных стероидгидроксилазных системах, основанный на структурных изменениях при восстановлении адренодоксин». Биохимия . 41 (25): 7969–78. дои : 10.1021/bi0160361 . ПМИД 12069587 .
- ^ Стюарт Д., Сандстрем М., Юсеф Х.М., Захрабекова С., Йенсен П.Е., Болливар Д.В. и др. (сентябрь 2020 г.). «Аэробный ячменный Mg-протопорфирин IX монометилэфирциклаза питается электронами ферредоксина» . Растения . 9 (9): 1157. doi : 10.3390/plants9091157 . ПМК 7570240 . ПМИД 32911631 .
- ^ Йех А.П., Амброджио XI, Андраде С.Л., Эйнсле О., Шатле С., Мейер Дж. и др. (сентябрь 2002 г.). «Кристаллические структуры дикого типа с высоким разрешением и варианты Cys-55→Ser и Cys-59→Ser тиоредоксинподобного [2Fe-2S] ферредоксина из Aquifex aeolicus» . Журнал биологической химии . 277 (37): 34499–507. дои : 10.1074/jbc.M205096200 . ПМИД 12089152 .
- ^ семья: "Семейство ферредоксина типа 2fe2s shethna"
- ^ ПДБ : 3П1М ; Чайкуад А., Йоханссон С., Кройер Т., Юэ и др. (2010). «Кристаллическая структура человеческого ферредоксина-1 (FDX1) в комплексе с железо-серным кластером». Всемирный банк данных по белкам . дои : 10.2210/pdb3p1m/pdb .
- ^ Петерсон Дж. А., Лоренс MC, Амарне Б. (апрель 1990 г.). «Путидаредоксинредуктаза и путидаредоксин. Клонирование, определение последовательности и гетерологичная экспрессия белков» . Журнал биологической химии . 265 (11): 6066–73. дои : 10.1016/S0021-9258(19)39292-0 . ПМИД 2180940 .
- ^ Петерсон Дж.А., Лу Дж.Ю., Гейссельсодер Дж., Грэм-Лоренс С., Кармона С., Уитни Ф. и др. (июль 1992 г.). «Цитохром Р-450терп. Выделение и очистка белка, клонирование и секвенирование его оперона» . Журнал биологической химии . 267 (20): 14193–203. дои : 10.1016/S0021-9258(19)49697-X . ПМИД 1629218 .
- ^ Токумото У, Такахаси Ю (июль 2001 г.). «Генетический анализ оперона isc Escherichia coli, участвующего в биогенезе клеточных железо-серных белков». Журнал биохимии . 130 (1): 63–71. doi : 10.1093/oxfordjournals.jbchem.a002963 . ПМИД 11432781 .
- ^ «Ген Энтреза: FDX1 ферредоксин 1» .
- ^ «Ферредоксин 2 FDX2 [Homo sapiens (человек)] - Ген - NCBI» . www.ncbi.nlm.nih.gov . Проверено 8 апреля 2019 г.
- ^ Фукуяма К., Мацубара Х., Цукихара Т., Кацубе Ю. (ноябрь 1989 г.). «Структура [4Fe-4S] ферредоксина из Bacillus thermoproteolyticus, уточненная с разрешением 2,3 А. Структурное сравнение бактериальных ферредоксинов». Журнал молекулярной биологии . 210 (2): 383–98. дои : 10.1016/0022-2836(89)90338-0 . ПМИД 2600971 .
- ^ Дуэ ЭД, Фанчон Э, Викат Дж, Сикер Л.С., Мейер Дж, Мулис Дж.М. (ноябрь 1994 г.). «Уточненная кристаллическая структура ферредоксина 2[4Fe-4S] из Clostridium acidurici при разрешении 1,84 А». Журнал молекулярной биологии . 243 (4): 683–95. дои : 10.1016/0022-2836(94) 90041-8 ПМИД 7966291 .
Дальнейшее чтение
[ редактировать ]- Бруски М, Герлекен Ф (1988). «Структура, функции и эволюция бактериальных ферредоксинов» . Обзоры микробиологии FEMS . 4 (2): 155–75. дои : 10.1111/j.1574-6968.1988.tb02741.x . ПМИД 3078742 .
- Чурли С., Мусиани Ф (2005). «Высокопотенциальные железо-серные белки и их роль в качестве растворимых переносчиков электронов в бактериальном фотосинтезе: рассказ об открытии». Исследования фотосинтеза . 85 (1): 115–31. Бибкод : 2005PhoRe..85..115C . дои : 10.1007/s11120-004-6556-4 . ПМИД 15977063 . S2CID 27768048 .
- Фукуяма К. (2004). «Структура и функции ферредоксинов растительного типа». Исследования фотосинтеза . 81 (3): 289–301. Бибкод : 2004PhoRe..81..289F . дои : 10.1023/B:PRES.0000036882.19322.0a . ПМИД 16034533 . S2CID 24574958 .
- Гринберг А.В., Ханнеманн Ф., Шиффлер Б., Мюллер Дж., Хайнеманн У., Бернхардт Р. (сентябрь 2000 г.). «Адренодоксин: структура, стабильность и свойства переноса электронов». Белки . 40 (4): 590–612. doi : 10.1002/1097-0134(20000901)40:4<590::AID-PROT50>3.0.CO;2-P . ПМИД 10899784 . S2CID 113757 .
- Холден Х.М., Джейкобсон Б.Л., Херли Дж.К., Толлин Г., О Б.Х., Скъельдал Л. и др. (февраль 1994 г.). «Структурно-функциональные исследования ферредоксинов [2Fe-2S]». Журнал биоэнергетики и биомембран . 26 (1): 67–88. дои : 10.1007/BF00763220 . ПМИД 8027024 . S2CID 12560221 .
- Мейер Дж. (ноябрь 2001 г.). «Ферредоксины третьего рода» . Письма ФЭБС . 509 (1): 1–5. дои : 10.1016/S0014-5793(01)03049-6 . ПМИД 11734195 . S2CID 8101608 .
Внешние ссылки
[ редактировать ]- InterPro : IPR006057 - субдомен ферредоксина 2Fe – 2S.
- ИнтерПро : IPR001055 - Адренодоксин
- InterPro : IPR001450 - ферредоксин 4Fe–4S, железо-серное связывание.
- InterPro : IPR000170 - Железо-серный белок с высоким потенциалом.
- PDB : 1F37 - Рентгеновская структура тиоредоксинподобного ферредоксина из Aquifex aeolicus ( Aa Fd)
| Базы данных органов управления : Национальные |
|---|