Эктодерма
| Эктодерма | |
|---|---|
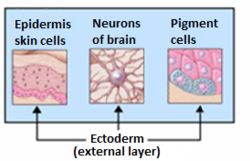 Органы, происходящие из эктодермы. | |
 Срез эмбрионального диска Vespertilio murinus . | |
| Подробности | |
| Дни | 16 |
| Идентификаторы | |
| МеШ | D004475 |
| ФМА | 69070 |
| Анатомическая терминология | |
Эктодерма образующихся — один из трёх первичных зародышевых листков, на ранних стадиях эмбрионального развития . Это самый внешний слой, расположенный поверхностно по отношению к мезодерме (средний слой) и энтодерме (самый внутренний слой). [1] Он возникает и берет свое начало из наружного слоя зародышевых клеток. Слово эктодерма происходит от греческого ektos, что означает «снаружи», и derma, что означает «кожа». [2]
Вообще говоря, эктодерма дифференцируется с образованием эпителиальных и нервных тканей ( спинного мозга , нервов и головного мозга ). Сюда входят кожа , слизистая оболочка рта, анус, ноздри, потовые железы, волосы и ногти. [3] и зубная эмаль . Другие типы эпителия происходят из энтодермы. [3]
У эмбрионов позвоночных эктодерму можно разделить на две части: эктодерму дорсальной поверхности, также известную как внешняя эктодерма, и нервную пластинку , которая инвагинирует, образуя нервную трубку и нервный гребень . [4] Поверхностная эктодерма дает начало большинству эпителиальных тканей, а нервная пластинка дает начало большинству нервных тканей . По этой причине нервная пластинка и нервный гребень также называются нейроэктодермой .
История
[ редактировать ]Хайнцу Кристиану Пандеру , балтийскому немецко -российскому биологу , приписывают открытие трёх зародышевых листков , которые формируются во время эмбриогенеза . Пандер получил докторскую степень по зоологии в Вюрцбургском университете в 1817 году. Он начал свои исследования в области эмбриологии с использованием куриных яиц, что позволило ему открыть эктодерму, мезодерму и энтодерму . Благодаря своим открытиям Пандера иногда называют «основателем эмбриологии».
Работу Пандера по раннему эмбриону продолжил прусско - эстонский биолог Карл Эрнст фон Бэр . Баер взял концепцию Пандера о зародышевых листках и посредством обширных исследований множества различных видов видов смог распространить этот принцип на всех позвоночных. Баер также получил признание за открытие бластулы . Бэр опубликовал свои открытия, в том числе теорию зародышевого слоя, в учебнике «О развитии животных» , который он выпустил в 1828 году. [5]
Дифференциация
[ редактировать ]Первоначальный внешний вид
[ редактировать ]Эктодерму впервые можно наблюдать у амфибий и рыб на поздних стадиях гаструляции . В начале этого процесса развивающийся эмбрион разделяется на множество клеток, образуя полый шар, называемый бластулой . Бластула полярна , две ее половины называются животным полушарием и вегетативным полушарием . Именно животное полушарие со временем станет эктодермой. [2]
Раннее развитие
[ редактировать ]Как и два других зародышевых листка – то есть мезодерма и энтодерма – эктодерма формируется вскоре после оплодотворения , после чего начинается быстрое деление клеток . Положение эктодермы относительно других зародышевых листков эмбриона определяется «селективным сродством», что означает, что внутренняя поверхность эктодермы имеет сильное (положительное) сродство к мезодерме и слабое (отрицательное) сродство к слой эндодермы. [6] Это избирательное сродство меняется на разных стадиях развития. Сила притяжения между двумя поверхностями двух зародышевых листков определяется количеством и типом молекул кадгерина, присутствующих на поверхности клеток. Например, экспрессия N-кадгерина имеет решающее значение для поддержания отделения нервных клеток-предшественников от эпителиальных клеток-предшественников. [2] Аналогично, пока поверхностная эктодерма становится эпидермисом , [6] нейроэктодерма , индуцируется вдоль нервного пути хордой которая обычно расположена над ней. [2] [4]
Гаструляция
[ редактировать ]В процессе гаструляции бутылочные клетки инвагинируют на дорсальной поверхности бластулы, образуя бластопор . Клетки продолжают расширяться внутрь и мигрировать вдоль внутренней стенки бластулы, образуя заполненную жидкостью полость, называемую бластоцель . Некогда поверхностным клеткам анимального полюса суждено стать клетками среднего зародышевого листка, называемого мезодермой. В процессе радиального растяжения клетки анимального полюса , которые когда-то имели толщину в несколько слоев, делятся, образуя тонкий слой. В то же время, когда этот тонкий слой делящихся клеток достигает дорсальной губы бластопора , происходит другой процесс, называемый конвергентным расширением . Во время конвергентного растяжения клетки, приближающиеся к губе, вставляются медиально-латерально, таким образом, что клетки вытягиваются за губу и внутрь эмбриона. Эти два процесса позволяют проспективным клеткам мезодермы располагаться между эктодермой и энтодермой. Как только начинаются конвергентное расширение и радиальная интеркаляция, остальная часть вегетативный полюс , который станет клетками энтодермы, полностью поглощается проспективной эктодермой, поскольку эти верхние клетки подвергаются эпиболии , при которой клетки эктодермы делятся, образуя один слой. Это создает однородный эмбрион, состоящий из трех зародышевых листков, расположенных в соответствующих положениях. [2]
Дальнейшее развитие
[ редактировать ]Как только будут созданы три зародышевых листка, клеточная дифференциация может произойти . Первым основным процессом здесь является нейруляция , при которой эктодерма дифференцируется с образованием нервной трубки , клеток нервного гребня и эпидермиса . Каждый из этих трех компонентов даст начало определенному набору клеток. Клетки нервной трубки дают начало центральной нервной системе , клетки нервного гребня дают начало периферической и кишечной нервной системе, меланоцитам и лицевым хрящам , а эпидермальная область дает начало эпидермису , волосам , ногтям , сальным железам , обонятельным органам. и эпителий полости рта, и глаза . [2]
нейруляция
[ редактировать ]Нейруляция состоит из двух частей: первичной и вторичной нейруляции. Оба процесса располагают клетки нервного гребня между поверхностным эпидермальным слоем и глубокой нервной трубкой. Во время первичной нейруляции клетки хорды мезодермы сигнализируют соседним, поверхностным клеткам эктодермы, чтобы они переместились в столбчатый узор с образованием клеток эктодермальной нервной пластинки . [7] Поскольку клетки продолжают удлиняться, группа клеток непосредственно над хордой меняет свою форму, образуя клин в эктодермальной области. Эти специальные клетки называются медиальными шарнирными клетками (MHP). Поскольку эктодерма продолжает удлиняться, эктодермальные клетки нервной пластинки сгибаются внутрь. Складывание эктодермы внутрь за счет преимущественно деления клеток продолжается до тех пор, пока внутри нервной пластинки не образуется другая группа клеток. Эти клетки называются дорсолатеральными шарнирными клетками (DLHP), и после их формирования сворачивание эктодермы внутрь прекращается. Клетки DLHP функционируют аналогично клеткам MHP в отношении их клиновидной формы, однако клетки DLHP приводят к сближению эктодермы. Эту конвергенцию возглавляют эктодермальные клетки над клетками DLHP, известные как нервный гребень. Клетки нервного гребня в конечном итоге стягивают соседние эктодермальные клетки вместе, в результате чего клетки нервного гребня остаются между предполагаемым эпидермисом и полой нервной трубкой. [2]
Органогенез
[ редактировать ]
Все органы, которые возникают из эктодермы, такие как нервная система, зубы, волосы и многие экзокринные железы, происходят из двух соседних слоев ткани: эпителия и мезенхимы. [8] Несколько сигналов опосредуют органогенез эктодермы, такие как: FGF , TGFβ , Wnt и регуляторы из семейства hedgehog . Конкретные сроки и способ формирования эктодермальных органов зависят от инвагинации эпителиальных клеток. [9] FGF-9 является важным фактором инициации развития зубных зачатков. Скорость эпителиальной инвагинации значительно увеличивается под действием FGF-9, который экспрессируется только в эпителии, а не в мезенхиме. FGF-10 помогает стимулировать пролиферацию эпителиальных клеток, что приводит к увеличению зубных зародышей. Зубы млекопитающих развиваются из эктодермы, происходящей из мезенхимы: оральной эктодермы и нервного гребня. Эпителиальные компоненты стволовых клеток непрерывно растущих зубов образуются из слоев ткани, называемых звездчатой сетью, и супрабазального слоя поверхностной эктодермы. [9]
Клиническое значение
[ редактировать ]Эктодермальная дисплазия
[ редактировать ]Эктодермальная дисплазия — редкое, но тяжелое заболевание, при котором группы тканей (особенно зубы, кожа, волосы, ногти и потовые железы), происходящие из эктодермы, подвергаются аномальному развитию. Это расплывчатый термин, поскольку существует более 170 подтипов эктодермальной дисплазии. Принято считать, что заболевание вызвано мутацией или комбинацией мутаций в определенных генах. Исследования заболевания продолжаются, поскольку выявлена лишь часть мутаций, связанных с подтипом эктодермальной дисплазии. [10]

Гипогидротическая эктодермальная дисплазия (ГЭД) является наиболее распространенным подтипом заболевания. Клинические случаи пациентов с этим заболеванием демонстрируют ряд симптомов. Наиболее значимым нарушением ГЭД является гипогидроз , неспособность вырабатывать достаточное количество пота, что объясняется отсутствием или дисфункцией потовых желез. Этот аспект представляет собой серьезный недостаток, особенно летом, ограничивает способность пациента заниматься спортом, а также его работоспособность и может быть особенно опасным в теплом климате, где пострадавшие люди подвергаются риску опасной для жизни гипертермии . лица Пороки развития также связаны с ГЭД, например, заостренные или отсутствующие зубы, морщинистая кожа вокруг глаз, нос неправильной формы, а также редкие и тонкие волосы. кожные проблемы, такие как экзема . В ряде случаев также наблюдаются [11] Большинство пациентов являются носителями вариантов Х-хромосомного гена EDA . [12] Это заболевание обычно более тяжело поражает мужчин, поскольку у них есть только одна Х-хромосома , тогда как у женщин второй, обычно непораженной Х-хромосомы может быть достаточно для предотвращения большинства симптомов.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Медицинская эмбриология Лангмана, 11-е издание. 2010.
- ^ Jump up to: а б с д и ж г Гилберт, Скотт Ф. Биология развития. 9-е изд. Сандерленд, Массачусетс: Sinauer Associates, 2010: 333–370. Распечатать.
- ^ Jump up to: а б «Происхождение тканей | Обучение SEER» . Training.seer.cancer.gov .
- ^ Jump up to: а б Мариб, Элейн Н.; Хен, Катя (2019). Анатомия и физиология человека . Соединенные Штаты Америки: Пирсон. стр. 146, 482–483, 1102–1106. ISBN 978-0-13-458099-9 .
- ^ Баер К.Э. фон (1986) В: Оппенгеймер Дж. (ред.) и Шнайдер Х. (перевод), Автобиография доктора. Карл Эрнст фон Бэр. Кантон, Массачусетс: Публикации по истории науки.
- ^ Jump up to: а б Хоссейни, Хади С.; Гарсия, Кара Э.; Табер, Ларри А. (2017). «Новая гипотеза формирования передней кишки и сердечной трубки, основанная на дифференциальном росте и сокращении актомиозина» . Разработка . 144 (13): 2381–2391. дои : 10.1242/dev.145193 . ПМК 5536863 . ПМИД 28526751 .
- ^ О'Рахили, Р.; Мюллер, Ф (1994). «Нейруляция в нормальном человеческом эмбрионе». Симпозиум 181 Фонда Ciba — Дефекты нервной трубки . Симпозиумы Фонда Новартис. Том. 181. С. 70–82. дои : 10.1002/9780470514559.ch5 . ISBN 9780470514559 . ПМИД 8005032 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Писпа, Дж; Теслефф, I (15 октября 2003 г.). «Механизмы эктодермального органогенеза» . Биология развития . 262 (2): 195–205. дои : 10.1016/S0012-1606(03)00325-7 . ПМИД 14550785 .
- ^ Jump up to: а б Тай, ГГ; Чен, РС; Лин, Ю.; Линг, Тайвань; Чен, Миннесота (2012). «FGF-9 ускоряет инвагинацию эпителия для эктодермального органогенеза при биоинженерных манипуляциях с органами в реальном времени» . Сотовая связь и сигнализация . 10 (1): 34. дои : 10.1186/1478-811X-10-34 . ПМЦ 3515343 . ПМИД 23176204 .
- ^ Приоло, М.; Лагана, К. (сентябрь 2001 г.). «Эктодермальные дисплазии: новая клинико-генетическая классификация» . Журнал медицинской генетики . 38 (9): 579–585. дои : 10.1136/jmg.38.9.579 . ПМЦ 1734928 . ПМИД 11546825 .
- ^ Кларк, А.; Филлипс, ДИ; Браун, Р.; Харпер, PS (1987). «Клинические аспекты Х-сцепленной гипогидротической эктодермальной дисплазии» . Архив болезней в детстве . 62 (10): 989–96. дои : 10.1136/adc.62.10.989 . ПМЦ 1778691 . ПМИД 2445301 .
- ^ Байес, М.; Хартунг, Эй-Джей; Эзер, С.; Писпа, Дж.; Теслефф, И.; Шривастава, АК; Кере, Дж. (1998). «Ген ангидротической эктодермальной дисплазии (EDA) подвергается альтернативному сплайсингу и кодирует эктодисплазин-А с делеционными мутациями в коллагеновых повторах» . Молекулярная генетика человека . 7 (11): 1661–1669. дои : 10.1093/hmg/7.11.1661 . ПМИД 9736768 .
| Базы данных органов управления : Национальные |
|---|