Бластоциста
| Бластоциста | |
|---|---|
 Бластоциста непосредственно перед имплантацией | |
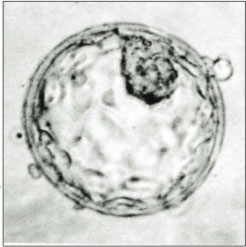 Человеческая бластоциста с внутренней клеточной массой вверху справа. | |
| Подробности | |
| Этап Карнеги | 3 |
| Дни | 5–9 |
| Дает начало | Гаструла |
| Идентификаторы | |
| латинский | бластоцистис |
| МеШ | D001755 |
| ТО | Е2.0.1.2.0.0.12 |
| ФМА | 83041 |
| Анатомическая терминология | |
Бластоциста стадиях — структура, образующаяся на ранних развития млекопитающих эмбрионального . Он имеет внутреннюю клеточную массу (ICM), также известную как эмбриобласт , которая впоследствии образует эмбрион , и внешний слой клеток трофобласта, называемый трофэктодермой . [1] [2] Этот слой окружает внутреннюю клеточную массу и заполненную жидкостью полость или просвет , известную как бластоцель . [3] В поздней бластоцисте трофэктодерма известна как трофобласт . [2] Трофобласт дает начало хориону и амниону — двум оболочкам плода , окружающим эмбрион. Плацента ворсинки происходит из эмбрионального хориона (части хориона, в которой развиваются ) и подлежащей ткани матки матери. [4] [5]
Название «бластоциста» происходит от греческого βλαστός blastós («росток») и κύστις kýstis («мочевой пузырь, капсула»).
У немлекопитающих животных это структура, состоящая из недифференцированного клубка клеток, и называется бластулой .
У человека образование бластоцисты начинается примерно через пять дней после оплодотворения , открывается заполненная жидкостью полость , когда в моруле , ранней эмбриональной стадии клубка из 16 клеток .Бластоциста имеет диаметр около 0,1–0,2 мм и состоит из 100–200 клеток после 7–8 раундов дробления (деление клеток без роста клеток). Примерно через семь дней после оплодотворения [6] бластоциста подвергается имплантации , внедряясь в эндометрий стенки матки , где она подвергается дальнейшим процессам развития, включая гаструляцию . Для внедрения бластоцисты в эндометрий необходимо, чтобы она вылупилась из зоны пеллюцида , яйцевой оболочки, которая предотвращает прикрепление к фаллопиевой трубе , когда преэмбрион попадает в матку.
Использование бластоцист при экстракорпоральном оплодотворении (ЭКО) предполагает культивирование оплодотворенной яйцеклетки в течение пяти дней перед ее переносом в матку. Это может быть более эффективным методом лечения бесплодия , чем традиционное ЭКО. Внутренняя клеточная масса бластоцисты является источником эмбриональных стволовых клеток , которые широко применимы в терапии стволовыми клетками, включая восстановление, замену и регенерацию клеток. Вспомогательный хэтчинг зоны также можно использовать при ЭКО и других методах лечения бесплодия.
Цикл разработки
[ редактировать ]Стадия бластоцисты наступает через 5–9 дней после зачатия. Во время эмбрионального развития , после оплодотворения (примерно через 5–6 дней у человека), клетки морулы начинают подвергаться клеточной дифференцировке , и морула превращается в бластоцисты путем перекачивания жидкости для роста просвета . В матке пеллюцидная зона , окружающая бластоцисту, разрушается, позволяя ей имплантироваться в стенку матки. Имплантация знаменует собой конец зародышевой стадии эмбриогенеза и начало беременности . [ нужна медицинская ссылка ]
Образование бластоцисты
[ редактировать ]
Зигота . несколько раундов митоза претерпевает После 3-го деления дробления у зародыша начинается процесс уплотнения, который у человека завершается только тогда, когда зародыш состоит из 8-16 клеток. [7] [8] затем стал известен как морула . актомиозина Уплотнение происходит в результате повышенной сократимости коры , которая стягивает клетки в более плотную конфигурацию. [9] [10] Повышенная сократимость при уплотнении наблюдается как у эмбрионов мыши, так и у человека. [11] [12] но сильнее у людей, что может способствовать его фрагментации . [13] До этой стадии развития клетки ( бластомеры ) не были отнесены к какой-либо конкретной клеточной линии, но при достижении 16-клеточной стадии клетки на поверхности эмбриона начинают дифференцироваться в трофэктодерму, в то время как клетки с внутренним положением инициируют свою дифференцировку во внутренние клетки. массовая судьба. [14] Затем морула в результате кавитации развивается и становится бластоцистой, а у многих других животных — бластулой. Затем дифференцировка клеток разделяет клетки морулы на два типа: клетки трофэктодермы , окружающие просвет , и внутреннюю массу клеток (эмбриобласты). Внутренняя клеточная масса лежит в основе эмбриональных стволовых клеток . [15] Концептус . тогда известен как бластоциста [16]
Прежде чем произойдет дифференцировка клеток, есть два фактора транскрипции , Oct-4 и nanog , которые равномерно экспрессируются во всех клетках, но оба этих фактора транскрипции отключаются в трофобласте после его формирования. [17] Внешние клетки трофэктодермы перекачивают ионы натрия в бластоцисты, что приводит к поступлению воды посредством осмоса . Накопление воды между межклеточными контактами разрушает их посредством гидроразрыва . [18] Затем жидкость собирается в единый просвет в процессе, похожем на созревание Оствальда , с образованием бластоцеля , который определяет первую ось симметрии эмбриона млекопитающих. [19] Сторона бластоцисты, где формируется внутренняя клеточная масса, называется эмбриональным полюсом, а противоположная сторона — абэмбриональным полюсом. Бластоцель трофэктодерма , внутренняя и клеточная масса являются отличительными чертами бластоцисты. [20]
Имплантация
[ редактировать ]Имплантация имеет решающее значение для выживания и развития раннего человеческого эмбриона. Он устанавливает связь между матерью и ранним эмбрионом, которая будет продолжаться до конца беременности. Имплантация становится возможной благодаря структурным изменениям как в бластоцисте, так и в стенке эндометрия. [21] Зона пеллюцида , окружающая бластоцисты, имеет разрывы, называемые штриховкой . Это устраняет ограничение на физический размер эмбриональной массы и открывает доступ внешним клеткам бластоцисты к внутренней части матки. Более того, гормональные изменения у матери, в частности пик лютеинизирующего гормона (ЛГ), подготавливают эндометрий к приему и обертыванию бластоцисты. Иммунная система также модулируется, чтобы обеспечить вторжение чужеродных эмбриональных клеток. После связывания с внеклеточным матриксом эндометрия клетки трофобласта секретируют ферменты и другие факторы, необходимые для внедрения бластоцисты в стенку матки. Высвобождаемые ферменты разрушают слизистую оболочку эндометрия, в то время как аутокринные факторы роста, такие как хорионический гонадотропин человека (ХГЧ) и инсулиноподобный фактор роста (ИФР), позволяют бластоцисте дальше проникать в эндометрий. [22]
Имплантация в стенку матки позволяет осуществить следующий этап эмбриогенеза — гаструляцию , которая включает формирование плаценты из трофобластических клеток и дифференцировку внутренней клеточной массы в амниотический мешок и эпибласт .
Структура
[ редактировать ]Существует два типа бластомеров: [23]
- Внутренняя клеточная масса, также известная как эмбриобласт , дает начало примитивной энтодерме и собственно эмбриону (эпибласту).
- Примитивная энтодерма развивается в амниотический мешок , который образует заполненную жидкостью полость, в которой находится эмбрион во время беременности. [24]
- Эпибласт дает начало трем зародышевым листкам развивающегося зародыша во время гаструляции ( энтодерма , мезодерма и эктодерма ).
- Трофобласт — это слой клеток, образующий внешнее кольцо бластоцисты, который соединяется с материнским эндометрием, образуя плаценту. Клетки трофобласта также секретируют факторы, образующие бластоцель. [25]
- После имплантации цитотрофобласт представляет собой внутренний слой трофобласта, состоящий из стволовых клеток , которые дают начало клеткам, включающим ворсинки хориона , плаценту и синцитиотрофобласт .
- После имплантации синцитиотрофобласт является самым внешним слоем трофобласта. Эти клетки секретируют протеолитические ферменты , разрушающие внеклеточный матрикс эндометрия и обеспечивающие имплантацию бластоцисты в стенку матки. [26]
Жидкостная полость бластоцеля содержит аминокислоты , факторы роста и другие молекулы, необходимые для клеточной дифференцировки . [27]
Спецификация ячейки
[ редактировать ]Множественные процессы контролируют спецификацию клеточных клонов в бластоцисте с образованием трофобласта, эпибласта и примитивной энтодермы. Эти процессы включают экспрессию генов, передачу сигналов между клетками, межклеточный контакт и позиционные взаимоотношения, а также эпигенетику .
Как только внутренняя клеточная масса сформировалась внутри бластоцисты, она готовится к дальнейшей спецификации в эпибласт и примитивную энтодерму. Этот процесс спецификации, известный как определение судьбы клеток , частично осуществляется с помощью передачи сигналов фактора роста фибробластов (FGF), который генерирует путь киназы MAP для изменения клеточных геномов. [28] Дальнейшая сегрегация бластомеров в трофэктодерму и внутреннюю клеточную массу регулируется гомеодоменным белком Cdx2 . Этот транскрипционный фактор подавляет экспрессию транскрипционных факторов Oct4 и Nanog в трофобласте . [29] Эти геномные изменения позволяют прогрессивную спецификацию как эпибластов, так и примитивной энтодермы в конце фазы развития бластоцисты, предшествующей гаструляции. Большая часть исследований, проводимых на этих ранних эмбриональных стадиях, проводится на эмбрионах мышей, и конкретные факторы могут различаться у разных млекопитающих.
Во время имплантации трофобласт дает начало внеэмбриональным мембранам и типам клеток , которые в конечном итоге образуют большую часть плаценты плода — специализированного органа, через который эмбрион получает материнское питание, необходимое для последующего экспоненциального роста. [30] Спецификация трофобласта контролируется комбинацией морфологических сигналов, возникающих из-за полярности клеток с дифференциальной активностью сигнальных путей, таких как Hippo и Notch, и ограничением внешних клеток спецификаторов клонов, таких как CDX2. [31]
У мышей первичные зародышевые клетки выделяются из клеток эпибласта , этот процесс сопровождается обширным генома перепрограммированием всего эпигенетическим . [32] Перепрограммирование включает глобальное деметилирование ДНК и реорганизацию хроматина клеток , что приводит к тотипотентности . [32] Процесс полногеномного ДНК деметилирования включает в себя путь эксцизионной репарации оснований . [33]
Трофобласты экспрессируют интегрин на поверхности своих клеток, что обеспечивает адгезию к внеклеточному матриксу стенки матки. Это взаимодействие делает возможным имплантацию и запускает дальнейшую спецификацию трех различных типов клеток, подготавливая бластоцисты к гаструляции. [34]
Клинические последствия
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( декабрь 2013 г. ) |
Тесты на беременность
[ редактировать ]Уровень хорионического гонадотропина человека (ХГЧ), секретируемого бластоцистой во время имплантации, является фактором, измеряемым при тесте на беременность . ХГЧ можно измерить как в крови, так и в моче, чтобы определить, беременна ли женщина. Больше ХГЧ выделяется при многоплодной беременности. Анализы крови на ХГЧ также можно использовать для проверки аномальной беременности.
Экстракорпоральное оплодотворение
[ редактировать ]Экстракорпоральное оплодотворение (ЭКО) является альтернативой традиционному оплодотворению in vivo, заключающемуся в оплодотворении яйцеклетки спермой и имплантации этого эмбриона в матку женщины. В течение многих лет эмбрион помещался в матку через два-три дня после оплодотворения. Однако на этой стадии развития очень сложно предсказать, какие эмбрионы будут развиваться лучше, и обычно имплантировали несколько эмбрионов. Несколько имплантированных эмбрионов увеличили вероятность развития плода, но также привели к развитию нескольких плодов. Это было серьезной проблемой и недостатком использования эмбрионов в ЭКО.
Использование бластоцист для ЭКО человека оказалось успешным. Бластоциста имплантируется через пять-шесть дней после оплодотворения яйцеклетки. [35] Через пять или шесть дней гораздо легче определить, из каких эмбрионов родятся здоровые живые существа. Знание того, какие эмбрионы будут успешными, позволяет имплантировать только одну бластоцисту, что резко снижает риск для здоровья и затраты на многоплодные роды. Теперь, когда определены потребности в питательных веществах для развития эмбриона и бластоцисты, гораздо легче давать эмбрионам правильные питательные вещества, необходимые для поддержания их в фазе бластоцисты.
Перенос эмбрионов после экстракорпорального оплодотворения — это процедура, при которой катетер вводится во влагалище, направляемый через шейку матки с помощью ультразвука, в полость матки, где бластоцисты вводятся в матку.
Бластоцисты также дают преимущество, поскольку их можно использовать для генетического тестирования клеток на наличие генетических проблем. В бластоцисте достаточно клеток, чтобы можно было удалить несколько клеток трофэктодермы, не повреждая развивающуюся бластоцисту. Эти клетки можно проверить на хромосом анеуплоидию с помощью преимплантационного генетического скрининга (ПГС) или специфических состояний, таких как муковисцидоз , часто известных как преимплантационная генетическая диагностика (ПГД). [36]
Процесс переноса эмбрионов
[ редактировать ]При процедуре переноса эмбрионов после первоначального ультразвукового исследования открываются с помощью зеркала стенки влагалища , и с помощью катетера эмбрион вводится через трубку для помещения в матку .
См. также
[ редактировать ]Ссылки
[ редактировать ]![]() Эта статья включает общедоступный текст из 20-го издания «Анатомии Грея» (1918 г.).
Эта статья включает общедоступный текст из 20-го издания «Анатомии Грея» (1918 г.).
- ^ «27.2C: Формирование бластоцисты» . Свободные тексты по медицине . 24 июля 2018 года . Проверено 11 октября 2022 г.
- ^ Jump up to: а б Стэндринг, Сьюзен (2016). Анатомия Грея: анатомические основы клинической практики (Сорок первое изд.). [Филадельфия]: Elsevier Limited. п. 167. ИСБН 9780702052309 .
- ^ Гилберт, Скотт Ф. (2000). «Раннее развитие млекопитающих» . Биология развития. 6-е издание . Проверено 13 мая 2022 г.
- ^ «трофобласт | эмбриология» . Британская энциклопедия . Проверено 1 ноября 2021 г.
- ^ Соломон, Эльдра (2018). Биология 11-е издание . Cengage Обучение. ISBN 978-1337392938 .
- ^ ВанПютте С (2020). Анатомия и физиология Сили . Нью-Йорк: МакГроу-Хилл. п. 1092. ИСБН 978-1-260-56596-6 . OCLC 1099344977 .
- ^ Джерри, Клаудия; Маккарти, Афшан; Аланис-Лобато, Грегорио; Демщенко, Андрей; Брюно, Александр; Луберсак, Софи; Фогарти, Нора М.Э.; Хэмпшир, Дэниел; Старейшина, Кей; Снелл, Фил; Кристи, Лейла; Дэвид, Лоран; Ван Де Вельде, Хильде; Фулади-Нашта, Али А.; Ниакан, Кэти К. (2020). «Инициирование консервативной программы трофэктодермы в эмбрионах человека, коровы и мыши» . Природа . 587 (7834): 443–447. Бибкод : 2020Natur.587..443G . дои : 10.1038/s41586-020-2759-x . ПМК 7116563 . ПМИД 32968278 .
- ^ Фирмин, Джули; Экер, Николас; Ривет Данон, Дайан; Озгюч, Озге; Барро Ланге, Вирджиния; Тюрлье, Эрве; Патрат, Кэтрин; Мастер Жан-Леон (2024 г.). «Механика уплотнения эмбриона человека» . Природа . 629 (8012): 646–651. Бибкод : 2024Natur.629..646F . дои : 10.1038/s41586-024-07351-x . ПМИД 38693259 .
- ^ Мэтр Жан-Леон; Ниваяма, Рицуя; Тюрлье, Эрве; Неделек, Франсуа; Хиираги, Такаши (2015). «Пульсирующая клеточно-автономная сократимость приводит к уплотнению эмбриона мыши» . Природная клеточная биология . 17 (7): 849–855. дои : 10.1038/ncb3185 . ПМИД 26075357 .
- ^ Фирмин, Джули; Экер, Николас; Ривет Данон, Дайан; Озгюч, Озге; Барро Ланге, Вирджиния; Тюрлье, Эрве; Патрат, Кэтрин; Мастер Жан-Леон (2024 г.). «Механика уплотнения эмбриона человека» . Природа . 629 (8012): 646–651. Бибкод : 2024Natur.629..646F . дои : 10.1038/s41586-024-07351-x . ПМИД 38693259 .
- ^ Мэтр Жан-Леон; Ниваяма, Рицуя; Тюрлье, Эрве; Неделек, Франсуа; Хиираги, Такаши (2015). «Пульсирующая клеточно-автономная сократимость приводит к уплотнению эмбриона мыши» . Природная клеточная биология . 17 (7): 849–855. дои : 10.1038/ncb3185 . ПМИД 26075357 .
- ^ Фирмин, Джули; Экер, Николас; Ривет Данон, Дайан; Озгюч, Озге; Барро Ланге, Вирджиния; Тюрлье, Эрве; Патрат, Кэтрин; Мастер Жан-Леон (2024 г.). «Механика уплотнения эмбриона человека» . Природа . 629 (8012): 646–651. Бибкод : 2024Natur.629..646F . дои : 10.1038/s41586-024-07351-x . ПМИД 38693259 .
- ^ Пельцер, Дайан; Де Платер, Людмила; Брэдбери, Пета; Эйхмюллер, Адриан; Бурдэ, Энн; Хале, Гийом; Мэтр, Жан-Леон (2023). «Фрагментация клеток в предимплантационных эмбрионах мышей, вызванная эктопической активацией пути экструзии полярных телец» . Журнал ЭМБО . 42 (17): e114415. дои : 10.15252/embj.2023114415 . ПМЦ 10476277 . ПМИД 37427462 .
- ^ Джерри, Клаудия; Маккарти, Афшан; Аланис-Лобато, Грегорио; Демщенко, Андрей; Брюно, Александр; Луберсак, Софи; Фогарти, Нора М.Э.; Хэмпшир, Дэниел; Старейшина, Кей; Снелл, Фил; Кристи, Лейла; Дэвид, Лоран; Ван Де Вельде, Хильде; Фулади-Нашта, Али А.; Ниакан, Кэти К. (2020). «Инициирование консервативной программы трофэктодермы в эмбрионах человека, коровы и мыши» . Природа . 587 (7834): 443–447. Бибкод : 2020Natur.587..443G . дои : 10.1038/s41586-020-2759-x . ПМК 7116563 . ПМИД 32968278 .
- ^ Мольнар, Чарльз; Гейр, Джейн (14 мая 2015 г.). «24.6. Оплодотворение и раннее эмбриональное развитие» . Проверено 19 октября 2022 г.
- ^ Ниссен С.Б., Перера М., Гонсалес Дж.М., Моргани С.М., Дженсен М.Х., Снеппен К. и др. (июль 2017 г.). «Четыре простых правила, которых достаточно для образования бластоцисты млекопитающих» . ПЛОС Биология . 15 (7): e2000737. дои : 10.1371/journal.pbio.2000737 . ПМК 5507476 . ПМИД 28700688 .
- ^ Шенвольф, Гэри К. (2015). Эмбриология человека Ларсена (Пятое изд.). Филадельфия, Пенсильвания. стр. 35–37. ISBN 9781455706846 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Дюмортье, Жюльен Г.; Ле Верж-Серандур, Матье; Торторелли, Анна Франческа; Мильке, Аннетт; Де Платер, Людмила; Тюрлье, Эрве; Мэтр, Жан-Леон (2019). «Гидравлический разрыв и активное огрубление позиционируют просвет бластоцисты мыши» . Наука . 365 (6452): 465–468. Бибкод : 2019Sci...365..465D . дои : 10.1126/science.aaw7709 . ПМИД 31371608 .
- ^ Дюмортье, Жюльен Г.; Ле Верж-Серандур, Матье; Торторелли, Анна Франческа; Мильке, Аннетт; Де Платер, Людмила; Тюрлье, Эрве; Мэтр, Жан-Леон (2019). «Гидравлический разрыв и активное огрубление позиционируют просвет бластоцисты мыши» . Наука . 365 (6452): 465–468. Бибкод : 2019Sci...365..465D . дои : 10.1126/science.aaw7709 . ПМИД 31371608 .
- ^ Гилберт С.Ф. (2000). «Раннее развитие млекопитающих» . Биология развития (6-е изд.). Сандерленд (Массачусетс): Sinauer Associates.
- ^ Чжан С., Линь Х., Конг С., Ван С., Ван Х., Ван Х., Армант Д.Р. (октябрь 2013 г.). «Физиологические и молекулярные детерминанты имплантации эмбриона» . Молекулярные аспекты медицины . 34 (5): 939–80. дои : 10.1016/j.mam.2012.12.011 . ПМЦ 4278353 . ПМИД 23290997 .
- ^ Шрисупарп С, Стракова З, Фазлеабас А.Т. (2001). «Роль хорионического гонадотропина (ХГ) в имплантации бластоцисты». Архивы медицинских исследований . 32 (6): 627–34. дои : 10.1016/S0188-4409(01)00330-7 . ПМИД 11750740 .
- ^ Гилберт С.Ф. (15 июля 2013 г.). Биология развития . Синауэр Ассошиэйтс, Инкорпорейтед. ISBN 978-1-60535-173-5 . [ нужна страница ]
- ^ Шенвольф Г.К., Ларсен В.Дж. (2009). Эмбриология человека Ларсена (4-е изд.). Филадельфия: Черчилль Ливингстон/Эльзевир. [ нужна страница ]
- ^ Джеймс Дж.Л., Stone PR, Чамли Л.В. (июль 2005 г.). «Дифференциация цитотрофобластов в первом триместре беременности: данные об отдельных предшественниках вневорсинчатых трофобластов и синцитиотрофобластов» . Размножение . 130 (1): 95–103. дои : 10.1530/rep.1.00723 . ПМИД 15985635 .
- ^ Вичовац Л., Аплин Дж.Д. (1996). «Эпителиально-мезенхимальный переход во время дифференцировки трофобласта». Акта Анатомика . 156 (3): 202–16. doi : 10.1159/000147847 (неактивен 9 июня 2024 г.). ПМИД 9124037 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на июнь 2024 г. ( ссылка ) - ^ Гасперович М., Натале Д.Р. (апрель 2011 г.). «Установить три линии бластоцисты — что потом?» . Биология размножения . 84 (4): 621–30. дои : 10.1095/biolreprod.110.085209 . ПМИД 21123814 .
- ^ Яманака Ю., Ланнер Ф., Россант Дж. (март 2010 г.). «Зависимая от сигнала FGF сегрегация примитивной энтодермы и эпибласта в бластоцисте мыши». Разработка . 137 (5): 715–24. дои : 10.1242/dev.043471 . ПМИД 20147376 . S2CID 28481311 .
- ^ Стрампф Д., Мао К.А., Яманака Ю., Ралстон А., Чавенгсаксофак К., Бек Ф., Россант Дж. (май 2005 г.). «Cdx2 необходим для правильной спецификации судеб клеток и дифференцировки трофэктодермы в бластоцисте мыши» . Разработка . 132 (9): 2093–102. дои : 10.1242/dev.01801 . hdl : 2381/1521 . ПМИД 15788452 .
- ^ Менчеро С., Сайнс де Ажа Х., Мансанарес М. (2018). «Наш первый выбор: клеточные и генетические основы идентичности и дифференциации трофэктодермы в эмбрионах млекопитающих». Актуальные темы биологии развития . 128 . Эльзевир: 59–80. дои : 10.1016/bs.ctdb.2017.10.009 . hdl : 20.500.12105/10490 . ISBN 978-0-12-804252-6 . ПМИД 29477171 .
- ^ Менчеро С., Роллан И., Лопес-Искьердо А., Андреу М.Дж., Сайнс де Ажа Дж., Канг М. и др. (апрель 2019 г.). «Переходы в активности клеток на ранних стадиях развития мышей обусловлены Notch» . электронная жизнь . 8 : е42930. doi : 10.7554/eLife.42930 . ПМК 6486152 . ПМИД 30958266 .
- ^ Jump up to: а б Хакетт Дж.А., Сенгупта Р., Зилич Дж.Дж., Мураками К., Ли С., Даун Т.А., Сурани М.А. (январь 2013 г.). «Динамика деметилирования зародышевой ДНК и стирание отпечатка с помощью 5-гидроксиметилцитозина» . Наука . 339 (6118): 448–52. Бибкод : 2013Sci...339..448H . дои : 10.1126/science.1229277 . ПМЦ 3847602 . ПМИД 23223451 .
- ^ Хайкова П., Джеффрис С.Дж., Ли С., Миллер Н., Джексон С.П., Сурани М.А. (июль 2010 г.). «Полногеномное перепрограммирование в зародышевой линии мыши влечет за собой путь эксцизионной репарации основания» . Наука . 329 (5987): 78–82. Бибкод : 2010Sci...329...78H . дои : 10.1126/science.1187945 . ПМЦ 3863715 . ПМИД 20595612 .
- ^ Дамский Ч., Либрах С., Лим К.Х., Фицджеральд М.Л., Макмастер М.Т., Джанатпур М. и др. (декабрь 1994 г.). «Переключение интегрина регулирует нормальную инвазию трофобласта» . Разработка . 120 (12): 3657–66. дои : 10.1242/dev.120.12.3657 . ПМИД 7529679 . Архивировано из оригинала 27 мая 2020 г. Проверено 10 декабря 2013 г.
- ^ Фонг С.И., Бонгсо А., Нг С.К., Анандакумар С., Траунсон А., Ратнам С. (март 1997 г.). «Продолжение нормальной беременности после переноса бластоцист без зоны: последствия переноса эмбрионов у человека» . Репродукция человека . 12 (3): 557–60. дои : 10.1093/humrep/12.3.557 . ПМИД 9130759 .
- ^ Ван Дж, Зауэр М.В. (декабрь 2006 г.). «Экстракорпоральное оплодотворение (ЭКО): обзор 3 десятилетий клинических инноваций и технологических достижений» . Терапия и управление клиническими рисками . 2 (4): 355–64. дои : 10.2147/tcrm.2006.2.4.355 . ЧВК 1936357 . ПМИД 18360648 .
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|