Клеточная кора
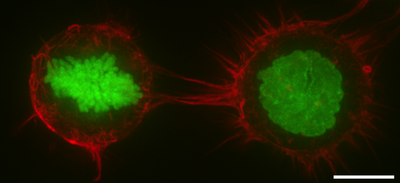
Клеточная кора , также известная как актиновая кора, кортикальный цитоскелет или актомиозиновая кора , представляет собой специализированный слой цитоплазматических белков на внутренней стороне клеточной мембраны . Он действует как модулятор поведения мембран и свойств клеточной поверхности. [1] [2] [3] В большинстве эукариотических клеток, лишенных клеточной стенки , кора представляет собой богатую актином сеть, состоящую из F-актиновых нитей, миозиновых моторов и актин-связывающих белков. [4] [5] Кора актомиозина прикрепляется к клеточной мембране с помощью мембрано-заякоренных белков, называемых белками ERM , которые играют центральную роль в контроле формы клеток. [1] [6] Белковые компоненты коры подвергаются быстрому обновлению, что делает кору одновременно механически жесткой и очень пластичной — два свойства, необходимые для ее функционирования. В большинстве случаев толщина коры составляет от 100 до 1000 нанометров .
В некоторых животных клетках белок спектрин может присутствовать в коре головного мозга. Спектрин помогает создать сеть из сшитых актиновых нитей. [3] Пропорции спектрина и актина варьируются в зависимости от типа клеток. [7] Спектриновые белки и актиновые микрофиламенты прикрепляются к трансмембранным белкам путем прикрепления белков между ними и трансмембранными белками. Кора клетки прикреплена к внутренней цитозольной поверхности плазматической мембраны в клетках, где белки-спектрины и микрофиламенты актина образуют сетчатую структуру, которая постоянно ремоделируется путем полимеризации , деполимеризации и разветвления.
Многие белки участвуют в регуляции и динамике коры головного мозга, в том числе формины , участвующие в полимеризации актина, комплексы Arp2/3 , которые приводят к разветвлению и кэпированию актина . Из-за процесса ветвления и плотности актиновой коры кортикальный цитоскелет может представлять собой очень сложную сеть, например фрактальную структуру. [8] Специализированные клетки обычно характеризуются очень специфическим кортикальным актиновым цитоскелетом. Например, в эритроцитах клеточная кора состоит из двумерной поперечно-сшитой эластичной сети с пятиугольной или гексагональной симметрией, привязанной к плазматической мембране и образованной в основном спектрином , актином и анкирином . [9] В аксонах нейронов актин или спектральный цитоскелет образует массив периодических колец. [10] а в спермия жгутике образует спиральную структуру. [11]
В растительных клетках клеточная кора усилена кортикальными микротрубочками, лежащими под плазматической мембраной. Направление этих кортикальных микротрубочек определяет, в какую сторону удлиняется клетка при росте.
Функции
[ редактировать ]Кора в основном функционирует, создавая напряжение под клеточной мембраной, позволяя клетке менять форму. [12] В первую очередь это достигается за счет моторов миозина II , которые тянут нити, создавая напряжение. [12] Эти изменения напряжения необходимы для того, чтобы клетка изменила свою форму, поскольку она подвергается клеточной миграции и клеточному делению . [12]
При митозе округлению F-актин и миозин II образуют высокосократительную и однородную кору головного мозга, способствующую митотических клеток . Поверхностное натяжение, создаваемое активностью коры актомиозина, создает внутриклеточное гидростатическое давление , способное смещать окружающие объекты, облегчая округление. [13] [14] Таким образом, клеточная кора служит для защиты веретена микротрубочек от внешнего механического разрушения во время митоза. [15] Когда к митотической клетке прикладывают внешние силы с достаточно большой скоростью и величиной, происходит потеря гомогенности кортикального F-актина, что приводит к образованию грыжи пузырей и временной потере способности защищать митотическое веретено. [16] [17] Генетические исследования показали, что кора клеток в митозе регулируется различными генами, такими как Rhoa, [18] WDR1, [19] белки ERM, [20] Экт2, [21] Pbl, Cdc42, aPKC, Par6, [22] DJ-1 и FAM134A. [23]
В цитокинезе кора клетки играет центральную роль, производя богатое миозином сократительное кольцо, сжимающее делящуюся клетку на две дочерние клетки. [24]
Сократимость коры клеток является ключевой для миграции клеток амебоидного типа, характерной для многих случаев метастазирования раковых клеток . [1] [25]
Помимо клеток кора также играет важную роль в формировании тканей, органов и организмов. Натягивая комплексы адгезии, кора способствует расширению контактов с другими клетками или с внеклеточным матриксом . Примечательно, что на ранних стадиях развития млекопитающих кора сближает клетки, вызывая уплотнение и образование морулы . [26] [27] Кроме того, различия в кортикальном напряжении способствуют сортировке внутренней клеточной массы и предшественников трофэктодермы во время формирования морулы . [28] сортировка зародышевого листка предшественников рыбок данио во время гаструляции , [29] [30] инвагинация мезодермы и удлинение зародышевого диска во время гаструляции дрозофилы. [31] [32]
Исследовать
[ редактировать ]Фундаментальные исследования клеточной коры проводятся с использованием иммортализованных клеточных линий , обычно клеток HeLa , клеток S2 , нормальных клеток почек крыс и клеток M2 . [12] В частности, в клетках М2 клеточные пузырьки , которые формируются без коры, а затем образуют ее по мере втягивания, часто используются для моделирования формирования и состава коры. [12]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Сальбре Г., Шаррас Г., Палюх Э. (октябрь 2012 г.). «Механика актиновой коры и клеточный морфогенез». Тенденции в клеточной биологии . 22 (10): 536–45. дои : 10.1016/j.tcb.2012.07.001 . ПМИД 22871642 .
- ^ Песен Д., Хо Дж.Х. (январь 2005 г.). «Микромеханическая архитектура коры эндотелиальных клеток» . Биофизический журнал . 88 (1): 670–9. Бибкод : 2005BpJ....88..670P . дои : 10.1529/biophysj.104.049965 . ПМК 1305044 . ПМИД 15489304 .
- ^ Перейти обратно: а б Альбертс, Брюс; Джонсон, Александр; Льюис, Джулиан; Рафф, Мартин; Робертс, Кейт; Уолтер, Питер (2002). «Сшивающие белки с разными свойствами организуют различные сборки актиновых нитей» . Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 0-8153-3218-1 .
- ^ Ганнинг П.В., Гошдастидер Ю., Уитакер С., Попп Д., Робинсон Р.К. (июнь 2015 г.). «Эволюция композиционно и функционально различных актиновых нитей» . Журнал клеточной науки . 128 (11): 2009–19. дои : 10.1242/jcs.165563 . ПМИД 25788699 .
- ^ Кларк А.Г., Вартлик О., Сальбре Г., Палух Е.К. (май 2014 г.). «Стрессы на поверхности клеток во время морфогенеза клеток животных» . Современная биология . 24 (10): Р484-94. Бибкод : 2014CBio...24.R484C . дои : 10.1016/j.cub.2014.03.059 . ПМИД 24845681 .
- ^ Фехон Р.Г., МакКлатчи А.И., Бретшер А. (апрель 2010 г.). «Организация клеточной коры: роль белков ERM» . Обзоры природы. Молекулярно-клеточная биология . 11 (4): 276–87. дои : 10.1038/nrm2866 . ПМК 2871950 . ПМИД 20308985 .
- ^ Махницка Б., Гроховальска Р., Богуславска Д.М., Сикорский А.Ф., Лекомт М.С. (январь 2012 г.). «Скелет на основе спектрина как участник клеточной сигнализации» . Клеточные и молекулярные науки о жизни . 69 (2): 191–201. дои : 10.1007/s00018-011-0804-5 . ПМЦ 3249148 . ПМИД 21877118 .
- ^ Садег С., Хиггинс Дж.Л., Маннион ПК, Тамкун М.М., Крапф Д. (2017). «Плазменная мембрана разделена самоподобной кортикальной актиновой сеткой» . Физический обзор X . 7 (1): 011031. arXiv : 1702.03997 . Бибкод : 2017PhRvX...7a1031S . дои : 10.1103/PhysRevX.7.011031 . ПМК 5500227 . ПМИД 28690919 .
- ^ Правительство Н.С. (январь 2007 г.). «Активная эластичная сеть: цитоскелет эритроцита». Физический обзор E . 75 (1 Pt 1): 011921. Бибкод : 2007PhRvE..75a1921G . дои : 10.1103/PhysRevE.75.011921 . ПМИД 17358198 .
- ^ Сюй К., Чжун Г, Чжуан Икс (январь 2013 г.). «Актин, спектрин и связанные с ними белки образуют периодическую структуру цитоскелета в аксонах» . Наука . 339 (6118): 452–6. Бибкод : 2013Sci...339..452X . дои : 10.1126/science.1232251 . ПМЦ 3815867 . ПМИД 23239625 .
- ^ Джерваси М.Г., Сюй X, Карбахал-Гонсалес Б., Буффоне М.Г., Висконти П.Е., Крапф Д. (июнь 2018 г.). «Актиновый цитоскелет жгутика сперматозоида мыши имеет спиральную структуру» . Журнал клеточной науки . 131 (11): jcs215897. дои : 10.1242/jcs.215897 . ПМК 6031324 . ПМИД 29739876 .
- ^ Перейти обратно: а б с д и Чуг П., Палуч Е.К. (июль 2018 г.). «Актиновая кора с первого взгляда» . J Cell Sci . 131 (14). дои : 10.1242/jcs.186254 . ПМК 6080608 . ПМИД 30026344 .
- ^ Стюарт М.П., Хелениус Дж., Тойода Ю., Раманатан С.П., Мюллер Д.Д., Хайман А.А. (январь 2011 г.). «Гидростатическое давление и кора актомиозина приводят к округлению митотических клеток». Природа . 469 (7329): 226–30. Бибкод : 2011Natur.469..226S . дои : 10.1038/nature09642 . ПМИД 21196934 . S2CID 4425308 .
- ^ Раманатан С.П., Хелениус Дж., Стюарт М.П., Кэттин С.Дж., Хайман А.А., Мюллер DJ (февраль 2015 г.). «Cdk1-зависимое митотическое обогащение кортикального миозина II способствует округлению клеток против удержания». Природная клеточная биология . 17 (2): 148–59. дои : 10.1038/ncb3098 . ПМИД 25621953 . S2CID 5208968 .
- ^ Ланкастер, Огайо (2013). «Митотическое округление изменяет геометрию клеток, обеспечивая эффективное формирование биполярного веретена» . Развивающая клетка . 25 (3): 270–283. дои : 10.1016/j.devcel.2013.03.014 . ПМИД 23623611 .
- ^ Шаррас, Гийом; Палух, Ева (сентябрь 2008 г.). «Пузыри прокладывают путь: как мигрировать без ламеллиподий». Nature Reviews Молекулярно-клеточная биология . 9 (9): 730–736. дои : 10.1038/nrm2453 . ПМИД 18628785 .
- ^ Каттин, Седрик (2015). «Механический контроль митотической прогрессии в отдельных клетках животных» . ПНАС . 112 (36): 11258–11263. Бибкод : 2015PNAS..11211258C . дои : 10.1073/pnas.1502029112 . ПМЦ 4568679 . ПМИД 26305930 .
- ^ Мэддокс, А. (2003). «RhoA необходим для ретракции коры и ригидности во время округления митотических клеток» . Дж. Клеточная Биол . 160 (2): 255–265. дои : 10.1083/jcb.200207130 . ПМК 2172639 . ПМИД 12538643 . S2CID 1491406 .
- ^ Фуджибучи, Т. (2005). «AIP1/WDR1 поддерживает округление митотических клеток». Биохим. Биофиз. Рез. Коммун . 327 (1): 268–275. дои : 10.1016/j.bbrc.2004.11.156 . ПМИД 15629458 .
- ^ Кунда, П. (2008). «Моезин контролирует жесткость коры, округление клеток и морфогенез веретена во время митоза» . Современная биология . 18 (2): 91–101. Бибкод : 2008CBio...18...91K . дои : 10.1016/j.cub.2007.12.051 . ПМИД 18207738 . S2CID 831851 .
- ^ Мэтьюз, Х (2013). «Изменения в локализации Ect2, пара актомиозин-зависимых изменений формы клеток с митотической прогрессией» . Развивающая клетка . 23 (2): 371–383. дои : 10.1016/j.devcel.2012.06.003 . ПМЦ 3763371 . ПМИД 22898780 . S2CID 1295956 .
- ^ Роза, А (2015). «Ect2/Pbl действует через Rho и белки полярности, направляя сборку изотропной коры актомиозина при входе в митоз» . Развивающая клетка . 32 (5): 604–616. дои : 10.1016/j.devcel.2015.01.012 . ПМК 4359025 . ПМИД 25703349 . S2CID 17482918 .
- ^ Тойода, Ю (2017). «Механическое фенотипирование отдельных клеток в масштабе генома выявляет гены, связанные с болезнями, участвующие в митотическом округлении» . Природные коммуникации . 8 (1): 1266. Бибкод : 2017NatCo...8.1266T . дои : 10.1038/s41467-017-01147-6 . ПМЦ 5668354 . ПМИД 29097687 . S2CID 19567646 .
- ^ Грин Р.А., Палух Э., Огема К. (ноябрь 2012 г.). «Цитокинез в животных клетках». Ежегодный обзор клеточной биологии и биологии развития . 28 : 29–58. doi : 10.1146/annurev-cellbio-101011-155718 . ПМИД 22804577 .
- ^ Олсон М.Ф., Сахай Э. (апрель 2009 г.). «Актиновый цитоскелет в подвижности раковых клеток» . Клинические и экспериментальные метастазы . 26 (4): 273–87. дои : 10.1007/s10585-008-9174-2 . ПМИД 18498004 .
- ^ Мэтр Жан-Леон; Ниваяма, Рицуя; Тюрлье, Эрве; Неделек, Франсуа; Хиираги, Такаши (июль 2015 г.). «Пульсирующая клеточно-автономная сократимость приводит к уплотнению эмбриона мыши». Природная клеточная биология . 17 (7): 849–855. дои : 10.1038/ncb3185 . ПМИД 26075357 .
- ^ Фирмин, Джули; Экер, Николас; Ривет Данон, Дайан; Озгюч, Озге; Барро Ланге, Вирджиния; Тюрлье, Эрве; Патрат, Кэтрин; Мастер Жан-Леон (16 мая 2024 г.). «Механика уплотнения эмбриона человека». Природа . 629 (8012): 646–651. Бибкод : 2024Natur.629..646F . дои : 10.1038/s41586-024-07351-x . ПМИД 38693259 .
- ^ Мастер Джон Леон; Тюрлье, Эрве; Иллюкумбура, Рукшала; Эйсманн, Медведь; Ниваяма, Рицуя; Неделек, Фрэнсис; Хиираги, Такаши (август 2016 г.). «Асимметричное деление сократительных доменов связывает расположение клеток и спецификацию судьбы» . Природа 536 (7616): 344–348. Бибкод : 2016Природа.536..344М . дои : 10.1038/nature18958 . ПМЦ 4998956 . ПМИД 27487217 .
- ^ Криг, М.; Арболеда-Эстудильо, Ю.; Пуэх, П.-Х.; Кафер, Дж.; Гранер, Ф.; Мюллер, диджей; Гейзенберг, К.-П. (апрель 2008 г.). «Силы растяжения управляют организацией зародышевого слоя у рыбок данио». Природная клеточная биология . 10 (4): 429–436. дои : 10.1038/ncb1705 . ПМИД 18364700 .
- ^ Мэтр Жан-Леон; Бертумье, Элен; Кренс, Саймон Фредерик Габриэль; Сальбре, Гийом; Юлихер, Франк; Палух, Ева; Гейзенберг, Карл-Филипп (12 октября 2012 г.). «Функции адгезии при сортировке клеток путем механического соединения кортикальных слоев прилипших клеток». Наука . 338 (6104): 253–256. Бибкод : 2012Sci...338..253M . дои : 10.1126/science.1225399 . ПМИД 22923438 .
- ^ Берте, Клэр; Сулак, Лоуренс; Лекуит, Томас (июнь 2004 г.). «Ремоделирование миозин-зависимого соединения контролирует интеркаляцию плоских клеток и удлинение оси». Природа . 429 (6992): 667–671. Бибкод : 2004Natur.429..667B . дои : 10.1038/nature02590 . ПМИД 15190355 .
- ^ Мартин, Адам С.; Кашубе, Матиас; Вишаус, Эрик Ф. (январь 2009 г.). «Импульсные сокращения актин-миозиновой сети вызывают апикальное сужение» . Природа . 457 (7228): 495–499. Бибкод : 2009Natur.457..495M . дои : 10.1038/nature07522 . ПМЦ 2822715 . ПМИД 19029882 .
Дальнейшее чтение
[ редактировать ]- Брей, Деннис (2000). «Актин и мембраны». Движения клеток . стр. 81–101. дои : 10.4324/9780203833582-7 . ISBN 978-0-203-83358-2 .