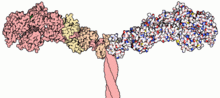
Часть структуры миозина II. Атомы тяжелой цепи окрашены в розовый цвет (слева); атомы в легких цепях окрашены в блекло-оранжевый и блекло-желтый цвет (также с левой стороны).
Хотя первоначально считалось, что миозин ограничен мышечными клетками (следовательно, мио- (s)+ -in ), единого «миозина» не существует; скорее, это очень большое суперсемейство генов, белковые продукты которых обладают общими основными свойствами связывания актина, гидролиза АТФ (активность фермента АТФазы) и силовой трансдукции. Практически все эукариотические клетки содержат изоформы миозина . Некоторые изоформы выполняют специализированные функции в определенных типах клеток (например, в мышцах), тогда как другие изоформы распространены повсеместно. Структура и функция миозина глобально консервативны у разных видов до такой степени, что миозин II мышц кролика связывается с актином амебы . [6] [7]
Большинство молекул миозина состоят из головки , шеи и хвоста.
Головной домен связывает нитчатый актин и использует АТФ гидролиз для создания силы и «прогулки» вдоль нити к зазубренному (+) концу (за исключением миозина VI, который движется к заостренному (-) концу).
шейный домен действует как линкер и как рычаг для передачи силы, генерируемой каталитическим моторным доменом. Шейковый домен также может служить местом связывания миозина легких цепей , которые представляют собой отдельные белки, образующие часть макромолекулярного комплекса и обычно выполняющие регуляторные функции.
Хвостовой домен обычно опосредует взаимодействие с молекулами-грузами и/или другими субъединицами миозина . В некоторых случаях хвостовой домен может играть роль в регуляции двигательной активности.
Множественные молекулы миозина II генерируют силу в скелетных мышцах посредством механизма силового удара, подпитываемого энергией, высвобождаемой в результате гидролиза АТФ. [8] Силовой удар возникает при высвобождении фосфата из молекулы миозина после гидролиза АТФ, когда миозин прочно связан с актином. Эффектом этого высвобождения является конформационное изменение молекулы, которая противодействует актину. Высвобождение молекулы АДФ приводит к так называемому строгому состоянию миозина. [9] Связывание новой молекулы АТФ высвободит миозин из актина. Гидролиз АТФ в миозине заставит его снова связаться с актином, чтобы повторить цикл. Комбинированный эффект множества мощных ударов заставляет мышцу сокращаться.
Широкое разнообразие генов миозина, обнаруженных во всех типах эукариот, было названо в соответствии с разными схемами по мере их открытия. Поэтому номенклатура может быть несколько запутанной при попытке сравнить функции белков миозина внутри и между организмами.
Миозин скелетных мышц, наиболее заметный представитель суперсемейства миозинов из-за его обилия в мышечных волокнах , был открыт первым. Этот белок составляет часть саркомера и образует макромолекулярные нити, состоящие из множества субъединиц миозина. Подобные миозиновые белки, образующие филаменты, были обнаружены в сердечной мышце , гладких мышцах и немышечных клетках. Однако начиная с 1970-х годов исследователи начали открывать новые гены миозина у простых эукариот. [5] кодирующие белки, которые действовали как мономеры и поэтому были названы миозинами класса I. Эти новые миозины получили общее название «нетрадиционные миозины». [10] и были обнаружены во многих тканях, помимо мышц. Эти новые члены суперсемейства были сгруппированы в соответствии с филогенетическими отношениями, полученными в результате сравнения аминокислотных последовательностей их головных доменов, при этом каждому классу присвоена римская цифра. [11] [12] [13] [14] (см. филогенетическое древо). Нетрадиционные миозины также имеют расходящиеся хвостовые домены, что предполагает уникальные функции. [15] Теперь разнообразный набор миозинов, вероятно, произошел от предкового предшественника (см. Рисунок).
Анализ аминокислотных последовательностей различных миозинов показывает большую вариабельность хвостовых доменов, но сильную консервативность последовательностей головных доменов. По-видимому, это происходит для того, чтобы миозины могли взаимодействовать через свои хвосты с большим количеством различных грузов, при этом цель в каждом случае – движение по актиновым нитям – остается той же самой и, следовательно, требует одних и тех же механизмов в моторе. Например, геном человека содержит более 40 различных генов миозина .
Эти различия в форме также определяют скорость, с которой миозины могут перемещаться по актиновым нитям. Гидролиз АТФ и последующее высвобождение фосфатной группы вызывает «силовой удар», при котором область «плеча рычага» или «шеи» тяжелой цепи тянется вперед. Поскольку рабочий ход всегда перемещает плечо рычага на один и тот же угол, длина плеча рычага определяет смещение груза относительно актиновой нити. Более длинное плечо рычага заставит груз пройти большее расстояние, хотя плечо рычага претерпевает такое же угловое смещение – точно так же, как человек с более длинными ногами может двигаться дальше с каждым отдельным шагом. Скорость миозинового мотора зависит от скорости, с которой он проходит полный кинетический цикл связывания АТФ с высвобождением АДФ.
Миозин I, вездесущий клеточный белок, действует как мономер и участвует в везикул . транспорте [16] Он имеет размер шага 10 нм и, как предполагается, отвечает за адаптационную реакцию стереоцилий во внутреннем ухе. [17]
Миозин II (также известный как обычный миозин) — это тип миозина, ответственный за сокращение мышц в мышечных клетках большинства типов клеток животных. Он также обнаружен в немышечных клетках сократительных пучков, называемых стрессовыми волокнами . [18]
Миозин II содержит две тяжелые цепи около 2000 аминокислот , каждая длиной , которые составляют головной и хвостовой домены. Каждая из этих тяжелых цепей содержит N-концевой головной домен, в то время как С-концевые хвосты принимают морфологию спиральной спирали , удерживая две тяжелые цепи вместе (представьте себе двух змей, обвивающих друг друга, как в кадуцее ). Таким образом, миозин II имеет две головки. Промежуточный домен шеи — это область, образующая угол между головой и хвостом. [19] В гладких мышцах один ген ( MYH11 ). [20] ) кодирует тяжелые цепи миозина II, но варианты сплайсинга этого гена приводят к образованию четырех различных изоформ. [19]
Он также содержит 4 легкие цепи миозина (MLC), в результате чего на головку приходится по 2, массой 20 (MLC 20 ) и 17 (MLC 17 ) кДа . [19] Они связывают тяжелые цепи в области «шеи» между головой и хвостом.
Самоингибирование миозина II. [21] [22] [23] Фильм начинается с Миозина II в конформации 10S со сложенным хвостовым доменом, заблокированной головкой и свободной головкой. [24] [25] В фильме схематически показано разворачивание хвоста и результирующее активное подтверждение 6S с последующим складыванием хвоста обратно в конформацию 10S. [26] [27] Иллюстрация концептуальная: переходные состояния и диффузионные движения, связанные со складыванием/разворачиванием, не показаны. [28] MLC 20 также известен как регуляторная легкая цепь и активно участвует в сокращении мышц . [19]
MLC 17 также известен как незаменимая легкая цепь . [19] Его точная функция неясна, но полагают, что он способствует структурной стабильности головки миозина вместе с MLC 20 . [19] Два варианта MLC 17 (MLC 17a/b ) существуют в результате альтернативного сплайсинга гена MLC 17 . [19]
В мышечных клетках длинные спиральные хвосты отдельных молекул миозина могут автоматически ингибировать активную функцию в конформации 10S или при фосфорилировании, переходить в конформацию 6S и соединяться, образуя толстые нити саркомера . [29] [30] Головные домены, производящие силу, выступают со стороны толстой нити, готовые идти вдоль соседних тонких нитей, основанных на актине, в ответ на соответствующие химические сигналы и могут находиться либо в автоингибированной, либо в активной конформации. Баланс/переход между активным и неактивным состояниями подлежит обширной химической регуляции.
Миозин IV имеет единственный мотив IQ и хвост, в котором отсутствует какая-либо последовательность, образующая спиральную спираль. Он имеет гомологию, аналогичную хвостовым доменам миозина VII и XV. [33]
Миозин V представляет собой нетрадиционный миозиновый мотор, который является процессивным как димер и имеет размер шага 36 нм. [34] Он перемещается (ходит) вдоль актиновых нитей, направляясь к зазубренному концу (+ концу) нитей. Миозин V участвует в транспортировке грузов (например, РНК, везикул, органелл, митохондрий) из центра клетки на периферию, но, кроме того, было показано, что он действует как динамическая привязь, удерживая везикулы и органеллы в богатых актином клетках. периферия клеток. [35] [36] Недавнее исследование восстановления одиночной молекулы in vitro при сборке актиновых нитей предполагает, что миозин V перемещается дальше при вновь собирающемся (богатом ADP-Pi) F-актине, в то время как процессивные длины короче на более старом (богатом ADP) F-актине. [37]
Ленточная диаграмма молекулярного двигателя Myosin V. [38] псевдоцветной для иллюстрации основных поддоменов. В целях наглядности важные петли (которые в литературе часто обозначаются отдельно) не выделяются. Эта перспектива подчеркивает сайт связывания нуклеотидов и разделение субдоменов U50 и L50, которые образуют расщелину сайта связывания актина.
Моторную головку Myosin V можно разделить на следующие функциональные области: [38]
Нуклеотид -связывающий сайт. Эти элементы вместе координируют катионы двухвалентных металлов (обычно магния ) и катализируют гидролиз:
Переключатель I — содержит высококонсервативный мотив SSR. Изомеризуется в присутствии АТФ .
Switch II — это киназа-GTPase-версия мотива Walker B DxxG. Изомеризуется в присутствии АТФ.
P-петля - содержит мотив Уокера A GxxxxGK(S,T). Это основной сайт связывания АТФ.
Преобразователь — семь β-нитей , лежащих в основе структуры моторной головки. [39]
U50 и L50. Верхний (U50) и нижний (L50) домены имеют массу около 50 кДа каждый . Их пространственное разделение [40] образует щель, необходимую для связывания с актином и некоторыми регуляторными соединениями.
Спираль SH1 и реле. Эти элементы вместе обеспечивают важный механизм связи ферментативного состояния моторного домена с областью, производящей силовой удар (конвертерный домен, плечо рычага и легкие цепи). [41] [42]
Преобразователь - преобразует изменение конформации головки двигателя в угловое смещение плеча рычага (в большинстве случаев усиленного легкими цепями). [42]
Состояние миозина VI PDB 2V26 перед силовым ударом [43]
Миозин VI представляет собой нетрадиционный миозиновый двигатель, который в основном является процессивным как димер, но также действует как непроцессивный мономер. Он идет вдоль актиновых нитей, направляясь к заостренному концу (-концу) нитей. [44] Считается, что миозин VI транспортирует эндоцитарные пузырьки в клетку. [45]
Миозин VIII — это специфичный для растений миозин, связанный с делением клеток; [48] в частности, он участвует в регуляции потока цитоплазмы между клетками. [49] и в локализации везикул во фрагмопласте . [50]
Миозин IX — группа одноголовых моторных белков. Впервые было показано, что он направлен минус-концом. [51] но более позднее исследование показало, что оно направлено на плюс-конец. [52] Механизм движения этого миозина плохо изучен.
Миозин X — это нетрадиционный миозиновый мотор, который функционирует как димер . Считается, что димеризация миозина X происходит антипараллельно. [53] Такое поведение не наблюдалось у других миозинов. Обнаружено, что в клетках млекопитающих мотор локализуется в филоподиях . Миозин X приближается к зазубренным концам нитей. Некоторые исследования показывают, что он предпочитает ходить по пучкам актина, а не по одиночным нитям. [54] Это первый миозиновый мотор, демонстрирующий такое поведение.
Миозин XI направляет движение органелл, таких как пластиды и митохондрии , в растительных клетках. [55] Он отвечает за светонаправленное движение хлоропластов в зависимости от интенсивности света и образование стромул, соединяющих разные пластиды. Миозин XI также играет ключевую роль в росте кончиков полярных корней и необходим для правильного удлинения корневых волосков . [56] Было обнаружено, что специфический миозин XI, обнаруженный в Nicotiana tabacum, является самым быстрым из известных процессивных молекулярных двигателей , движущихся со скоростью 7 мкм/с с шагом 35 нм вдоль актиновой нити. [57]
Эта группа миозина была обнаружена в типе Apicomplexa . [58] Миозины локализуются на плазматических мембранах внутриклеточных паразитов и затем могут участвовать в процессе клеточной инвазии. [59]
Этот миозин также обнаружен у реснитчатых простейших Tetrahymena thermaphila . Известные функции включают: транспортировку фагосом в ядро и нарушение регулируемого развитием элиминации макронуклеуса во время конъюгации.
Миозин XV необходим для развития актиновой основной структуры неподвижных стереоцилий, расположенных во внутреннем ухе. Считается, что он функционален как мономер.
MYO18A Ген на хромосоме 17q11.2, который кодирует моторные молекулы на основе актина с АТФазной активностью, которые могут участвовать в поддержании каркаса стромальных клеток, необходимого для поддержания межклеточного контакта.
Легкие цепи миозина различны и обладают своими свойствами. Они не считаются «миозинами», но являются компонентами макромолекулярных комплексов, составляющих функциональные ферменты миозина.
Парамиозин представляет собой крупный мышечный белок массой 93–115 кДа , который был описан у ряда различных типов беспозвоночных . [61] Считается, что толстые нити беспозвоночных состоят из внутреннего ядра парамиозина, окруженного миозином. Миозин взаимодействует с актином , что приводит к сокращению волокон. [62] Парамиозин обнаружен у многих различных видов беспозвоночных, например, у Brachiopoda , Sipunculidea , Nematoda , Annelida , Mollusca , Arachnida и Insecta . [61] Парамиозин отвечает за механизм «захвата», который обеспечивает устойчивое сокращение мышц с очень небольшими затратами энергии, так что моллюск может оставаться закрытым в течение длительного времени.
^ Гудсон Х.В. (1994). «Молекулярная эволюция суперсемейства миозина: применение филогенетических методов к биологическим вопросам клетки». Серия Общества общих физиологов . 49 : 141–157. ПМИД 7939893 .
^ Мацуока Р., Ёсида М.К., Фурутани Ю., Имамура С., Канда Н., Янагисава М. и др. (апрель 1993 г.). «Ген тяжелой цепи миозина гладкой мускулатуры человека, картированный в хромосомной области 16q12». Американский журнал медицинской генетики . 46 (1): 61–67. дои : 10.1002/ajmg.1320460110 . ПМИД 7684189 .
^ Петтерсен Э.Ф., Годдард Т.Д., Хуанг CC, Коуч Г.С., Гринблатт Д.М., Мэн ЕС и др. (октябрь 2004 г.). «UCSF Chimera - система визуализации для поисковых исследований и анализа». Журнал вычислительной химии . 25 (13): 1605–1612. дои : 10.1002/jcc.20084 . ПМИД 15264254 .
^ Хаммер Дж. А., Селлерс-младший (декабрь 2011 г.). «Поход на работу: роль миозинов класса V как переносчиков грузов». Обзоры природы. Молекулярно-клеточная биология . 13 (1): 13–26. дои : 10.1038/nrm3248 . ПМИД 22146746 . S2CID 11853457 .
^ Иноуэ А., Сайто Дж., Икебе Р., Икебе М. (апрель 2002 г.). «Миозин IXb представляет собой одноголовый процессивный двигатель, направленный на отрицательный конец». Природная клеточная биология . 4 (4): 302–306. дои : 10.1038/ncb774 . ПМИД 11901422 . S2CID 12158370 .
^ О'Коннелл CB, Мусекер MS (февраль 2003 г.). «Нативный миозин-IXb представляет собой двигатель, направленный на плюс, а не на минус». Природная клеточная биология . 5 (2): 171–172. дои : 10.1038/ncb924 . ПМИД 12563277 . S2CID 687308 .
Arc.Ask3.Ru Номер скриншота №: 65161e6cad681de6a8289bbe42b64868__1722589800 URL1:https://arc.ask3.ru/arc/aa/65/68/65161e6cad681de6a8289bbe42b64868.html Заголовок, (Title) документа по адресу, URL1: Myosin - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)




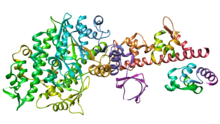
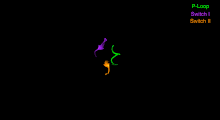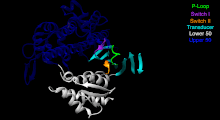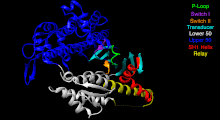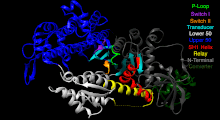

 Этап 1
Этап 1 Этап 2
Этап 2 Этап 3
Этап 3 Этап 4
Этап 4


