МИО10
| МИО10 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | MYO10 , миозин X, MyoX | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 601481 ; МГИ : 107716 ; Гомологен : 36328 ; Генные карты : MYO10 ; ОМА : MYO10 – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




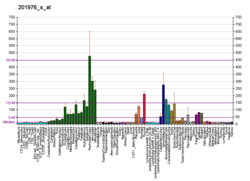
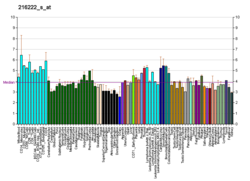
Миозин X , также известный как MYO10 , представляет собой белок , который у человека кодируется MYO10 геном . [5] [6] [7] [8]
Myo10 — это актина на основе моторный белок , который может локализоваться на кончиках пальцеобразных клеточных выступов, известных как филоподии . [9] [10] Myo10 широко экспрессируется в тканях млекопитающих, хотя и на относительно низких уровнях. [8] Исследования на нокаутных мышах показывают, что Myo10 выполняет важные функции в эмбриональных процессах, таких как закрытие нервной трубки и развитие глаз. [11] [12] [13] Myo10 также выполняет важные функции при инвазии и росте рака. [14] [15] [9] [12] [16] [17]
Myo10 не следует путать с Myh10 , который кодирует тяжелую цепь миозина класса II, известного как немышечный миозин 2b.
Структура и функции
[ редактировать ]
человека Ген MYO10 занимает ~274 т.п.н. и расположен на полосе 5p15.1 хромосомы 5 (выпуск 89 GRCh Ensembl). Он производит полноразмерный транскрипт РНК с 41 экзоном, кодирующий тяжелую цепь MYO10, рассчитанная последовательность которой имеет 2058 аминокислот и прогнозируемую молекулярную массу ~ 237 кДа. Как и многие моторные белки, полноразмерный белок Myo10 можно считать состоящим из головки, шеи и хвоста. [8] [21] N-концевая головка или моторный домен миозина может связываться с актиновой нитью, гидролизовать АТФ и создавать силу. [22] [23] Связывающий домен шеи или легкой цепи состоит из 3 мотивов IQ , каждый из которых обеспечивает сайт связывания для одной молекулы кальмодулина , кальций-связывающего белка ~16,5 кДа. [22] В отличие от большинства сайтов связывания кальмодулина, которые связываются с кальмодулином только в присутствии кальция, мотивы IQ в Myo10 могут связываться с кальмодулином в отсутствие кальция. Сообщалось также, что мотивы Myo10 IQ связывают CALML3 , кальмодулинподобный белок, экспрессируемый в эпителиальных клетках, поэтому в некоторых ситуациях CALML3 может служить легкой цепью Myo10 вместо кальмодулина. [24] Хвост Myo10 начинается с альфа-спиральной области, проксимальная часть которой образует единственную стабильную альфа-спираль (домен SAH), которая удлиняет плечо рычага, образованное шейным доменом. [25] [26] Дистальная часть альфа-спиральной области может самоассоциироваться с Kd ~0,6 мкм с образованием антипараллельной спиральной спирали , что позволяет двум тяжелым цепям Myo10 образовывать антипараллельный димер, уникальную структуру среди известных миозинов. [26] [27]
Хвост Myo10 включает в себя несколько областей помимо SAH и спиральной катушки. К ним относятся область с тремя последовательностями PEST — последовательностями, обогащенными аминокислотами пролин (P), глутамин (E), серин (S) и T (треонин), которые часто связаны с расщеплением протеазами, такими как кальпаин . [8] Хвост Myo10 уникален среди известных миозинов, поскольку содержит 3 домена PH (домен гомологичности плекстрина), домен, часто участвующий в связывании с мембранами. Последовательность первого домена PH Myo10 несколько необычна тем, что она разделена наличием поверхностной петли, содержащей второй домен PH. [8] [19] Второй домен PH связывается с важным сигнальным липидом фосфатидилинозитол (3,4,5)-трифосфатом [PI(3,4,5)P 3 ] и, как сообщается, в некоторых ситуациях связывается с фосфатидилинозитол (4,5)-бисфосфатом. [ПИ(4,5)Р 2 ]. [28] [19] [29] Считается, что 3 домена PH Myo10 работают вместе, рекрутируя его на плазматическую мембрану. Хвост Myo10 заканчивается супрамодулем, состоящим из домена MyTH4 (гомология хвоста миозина 4) и домена FERM (полоса 4.1, эзрин, радиксин, моэзин). [18] [20] Домен MyTH4 Myo10 может связываться с микротрубочками с заявленной аффинностью ~0,24 мкМ и дает полноразмерному Myo10 важную способность связывать актиновую нить, связанную его головкой, с микротрубочкой, связанной его хвостом. [30] [18] [31] Домен Myo10 FERM может связываться с цитоплазматическими доменами нескольких β- интегринов , основного класса рецепторов клеточной адгезии, а также с цитоплазматическими доменами рецепторов нетрина , удаленных при колоректальном раке ( DCC ) и неогенина ( Neo1 ). [32] [33] Хотя полноразмерный белок Myo10, по-видимому, экспрессируется на относительно низких уровнях, его можно обнаружить в большинстве тканей млекопитающих, включая мозг, семенники, почки, легкие, желудок и поджелудочную железу. [8]
Нативная полноразмерная тяжелая цепь Myo10 может существовать в виде мономера с 3 кальмодулин/кальмодулинподобными легкими цепями или в виде антипараллельного димера с 6 кальмодулин/кальмодулинподобными легкими цепями. Таким образом, антипараллельный димер Myo10 со всеми 6 легкими цепями будет иметь 8 субъединиц и нативную молекулярную массу ~574 кДа. Важно отметить, что хвост мономера Myo10 может загибаться назад на голову, подавляя двигательную активность головы. [29] Предполагается, что повышение уровня PI(3,4,5)P3 в плазматической мембране приводит к привлечению мономеров Myo10 к плазматической мембране через их PH-домены, активируя их двигательную активность и увеличивая их локальную концентрацию, что приводит к образованию активных антипараллельных димеров, которые способны перемещаться по актиновым нитям. Myo10, как и все известные миозины, кроме Myo6 , движется к зазубренному концу актиновой нити. [34] Myo10 способен гидролизовать ~10-20 АТФ/с на голову и, как сообщается, генерирует движение со скоростью ~300-1500 нм/с. [23] [27] Исследования одиночных молекул показывают, что нативные димеры Myo10 могут совершать шаги длиной до ~55 нм, что является одним из самых больших шагов, зарегистрированных для моторного белка. [27] Большой размер шага Myo10 отчасти обусловлен длинным плечом рычага, образованным его шейкой и стабильной альфа-спиралью, а частично из-за чрезвычайно большого поворота (около 120°), которому подвергается плечо рычага Myo10 во время рабочего хода. [27] Существует большой интерес к механизмам, которые нацеливают Myo10 на филоподиальные пучки актина, а также к способности Myo10 переходить от одной актиновой нити в пучке к другой. [35] [27] [36] В дополнение к полноразмерному Myo10, описанному выше, использование альтернативных сайтов начала транскрипции, расположенных в интронах 19-20 полноразмерного транскрипта, приводит к образованию «безголовых» транскриптов Myo10, в которых отсутствует большая часть головного домена миозина, но включают остальную часть тяжелой цепи Myo10. [37] [38] [39] Предполагается, что основные безголовые транскрипты у человека будут включать экзоны 20-41 полноразмерного MYO10, а инициация трансляции на M644 приведет к образованию безголового белка из 1415 аминокислот с прогнозируемой молекулярной массой ~ 163 кДа, который будет идентичен аминокислотам 644. -2058 полнометражного MYO10. [37] Поскольку у безголового Myo10 отсутствует большая часть головного домена, ему не хватает двигательной активности, но он сохраняет все другие домены Myo10 и, таким образом, ожидается, что он сохранит способность связываться с легкими цепями суперсемейства кальмодулина, с мембранами, содержащими PI(3,4, 5)P 3 или PI(4,5)P 3 , к микротрубочкам и к белкам, которые связывают хвост Myo10, таким как DCC, неогенин и β-интегрины. Было высказано предположение, что безголовый Myo10 действует как каркасный белок для своих различных партнеров по связыванию и/или как «естественный» доминантный негатив, который может ингибировать действие полноразмерного Myo10. [37] [38]
Эволюционные отношения
[ редактировать ]Myo10 является членом эволюционно древней группы миозинов, чьи хвосты содержат домены MyTH4-FERM и которые, как было показано, выполняют важные функции в клеточных выпячиваниях, основанных на пучках актина, таких как филоподии, микроворсинки и стереоцилии внутреннего уха . [40] [41] Слизевики Dictyostelium экспрессируют миозин MyTH4-FERM, известный как миозин-7, который участвует в формировании филоподий и имеет 2 супрамодуля MyTH4-FERM, но не имеет доменов PH. [40] Myo10, по-видимому, произошел от предкового миозин-7-подобного белка примерно миллиард лет назад в результате нескольких изменений, включая потерю 1 супрамодуля MyTH4-FERM и добавление 3 доменов PH. Ген Myo10 присутствует во многих организмах: от филозоев и хоанофлагеллят (группы простейших, наиболее тесно связанных с многоклеточными животными) до человека. [40] Myo10 был потерян в линиях беспозвоночных, ведущих к таким организмам, как плодовые мухи и нематоды, хотя эти линии действительно экспрессируют другие миозины MyTH4-FERM, такие как миозин-7. В дополнение к Myo10 люди экспрессируют 3 миозина MyTH4-FERM: MYO7A , ген, который мутирует при синдроме Ашера 1b, слепоглухости; MYO7B, компонент адгезионного комплекса на кончиках микроворсинок; и MYO15A , миозин, который локализуется на кончиках стереоцилий внутреннего уха и мутирует при глухоте DFNB3. [41] Головные домены других миозинов MyTH4-FERM, экспрессируемых у человека, демонстрируют не более 45% общей идентичности аминокислотных последовательностей с Myo10, а каждый из их хвостовых доменов содержит 2 домена MyTH4-FERM вместо 3 доменов PH и 1 домена MyTH4-FERM в Myo10. . [8] [41]
Клеточная функция
[ редактировать ]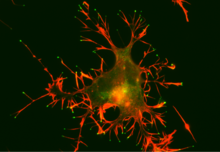
Myo10 может локализоваться на кончиках филоподий, чего нет у большинства других миозинов. Когда Myo10 был помечен зеленым флуоресцентным белком (GFP) и экспрессирован в клетках, наблюдалось движение небольших точек GFP-Myo10 вперед внутри филоподий к кончику со скоростью ~ 100 нм/с. [42] Визуализация с чувствительностью к одиночным молекулам выявила сходные движения отдельных димеров Myo10 со скоростью ~ 600-1400 нм/с. [43] [44] [45] GFP-Myo10 также движется назад в филоподиях со скоростью ретроградного потока ~15 нм/с. Эти наблюдения привели к гипотезе, что молекулы Myo10 используют свою двигательную активность для быстрого перемещения вперед вдоль филоподиальных актиновых филаментов и могут связываться с филоподиальными актиновыми нитями, которые медленно переносятся назад ретроградным потоком актина. [42] Эта «внутрифилоподиальная подвижность» Myo10 привела к предположению, что Myo10 функционирует как моторный белок для транспортировки грузов внутри филоподий. Myo10 также выполняет важные функции в формировании и/или стабилизации филоподий, при этом сверхэкспрессия Myo10 увеличивает количество и длину филоподий, тогда как нокдаун или нокаут Myo10 уменьшает филоподии. [42] [46] [47] Myo10 также выполняет важные функции в делении клеток митотического веретена . , особенно в ориентации [30] [48] [49] [50] Myo10 также необходим для кластеризации избыточных центросом , которые являются отличительной чертой раковых клеток. [49] процесс представляет большой интерес, поскольку раковым клеткам для успешного деления необходимо группировать свои центросомы.
Роль в болезни
[ редактировать ]Все больше данных показывает, что Myo10 играет важную роль в развитии рака. [9] Помимо своей роли в кластеризации избыточных центросом раковых клеток, [49] Myo10 является ключевым компонентом инвадоподий , выступов, связанных с филоподиями, которые раковые клетки используют для вторжения в окружающую среду. [51] Сообщалось также, что несколько микроРНК, которые подавляют инвазию раковых клеток, частично действуют путем нацеливания на мРНК Myo10. [52] [53] Сообщается, что нокаут или нокдаун Myo10 подавляет инвазию или распространение раковых клеток в экспериментальных моделях рака молочной железы. [14] [15] рак легких, [17] и глиома , [16] где нокаут Myo10 также увеличил эффективность химиотерапевтического агента, который в противном случае был бы неэффективен. Эти результаты, а также исследования, показывающие, что нокаут Myo10 увеличивает время выживания на 260% в мышиной модели меланомы . [12] сделать Myo10 потенциальной мишенью для борьбы с раком.
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Ensembl выпуск 89: ENSG00000145555 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000022272 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ген Энтрез: миозин X MYO10» .
- ^ Бемент В.М., Хассон Т., Вирт Дж.А., Чейни Р.Э., Мусекер М.С. (июль 1994 г.). «Идентификация и перекрывающаяся экспрессия множества нетрадиционных генов миозина в типах клеток позвоночных» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (14): 6549–53. Бибкод : 1994PNAS...91.6549B . дои : 10.1073/pnas.91.14.6549 . ПМК 44240 . ПМИД 8022818 .
- ^ Хэссон Т., Скоурон Дж.Ф., Гилберт Д.Д., Авраам К.Б., Перри В.Л., Бемент В.М. и др. (сентябрь 1996 г.). «Картирование нетрадиционных миозинов у мышей и человека» . Геномика . 36 (3): 431–9. дои : 10.1006/geno.1996.0488 . ПМИД 8884266 .
- ^ Jump up to: а б с д и ж г час Берг Дж.С., Дерфлер Б.Х., Пенниси К.М., Кори Д.П., Чейни Р.Э. (октябрь 2000 г.). «Миозин-X, новый миозин с доменами, гомологичными плекстрину, ассоциируется с областями динамического актина» . Журнал клеточной науки . 113. 113 Пт 19 (19): 3439–51. дои : 10.1242/jcs.113.19.3439 . ПМИД 10984435 .
- ^ Jump up to: а б с Курсон Д.С., Чейни Р.Э. (май 2015 г.). «Миозин-X и болезнь» . Экспериментальные исследования клеток . 334 (1): 10–5. дои : 10.1016/j.yexcr.2015.03.014 . ПМЦ 4433855 . ПМИД 25819274 .
- ^ Токуо Х (2020). «Миозин Х». Миозины . Достижения экспериментальной медицины и биологии. Том. 1239. стр. 391–403. дои : 10.1007/978-3-030-38062-5_17 . ISBN 978-3-030-38061-8 . ПМИД 32451868 . S2CID 218892501 .
- ^ Хеймсат Э.Г., Йим Й.И., Мустафа М., Хаммер Дж.А., Чейни Р.Э. (декабрь 2017 г.). «Нокаут миозина-X является полулетальным и демонстрирует, что миозин-X участвует в закрытии нервной трубки, пигментации, регрессии гиалоидных сосудов и формировании филоподий» . Научные отчеты . 7 (1): 17354. Бибкод : 2017NatSR...717354H . дои : 10.1038/s41598-017-17638-x . ПМЦ 5725431 . ПМИД 29229982 .
- ^ Jump up to: а б с Токуо Х., Бхаван Дж., Колуччо Л.М. (июль 2018 г.). «Миозин X необходим для эффективной миграции меланобластов, возникновения и метастазирования меланомы» . Научные отчеты . 8 (1): 10449. Бибкод : 2018NatSR...810449T . дои : 10.1038/s41598-018-28717-y . ПМК 6041326 . ПМИД 29993000 .
- ^ Бахг А.С., Хорстемке М., Скрябин Б.В., Класен Т., Нагельманн Н., Фабер С. и др. (январь 2019 г.). «Фенотипический анализ нокаута Myo10 (Myo10 тм2/тм2 ) мыши, у которых отсутствует полноразмерный (моторизованный), но не специфичный для мозга безголовый миозин X" . Scientific Reports . 9 (1): 597. Bibcode : 2019NatSR...9..597B . doi : 10.1038/s41598-018-37160- у . ПМЦ 6345916 .
- ^ Jump up to: а б Арйонен А., Кауконен Р., Маттила Е., Роухи П., Хёгнес Г., Сихто Х. и др. (март 2014 г.). «Повышение регуляции миозина-X, связанное с мутантом p53, способствует инвазии и метастазированию рака молочной железы» . Журнал клинических исследований . 124 (3): 1069–82. дои : 10.1172/JCI67280 . ПМЦ 3934176 . ПМИД 24487586 .
- ^ Jump up to: а б Цао Р., Чен Дж., Чжан Икс, Чжай Ю, Цин Икс, Син В и др. (июль 2014 г.). «Повышенная экспрессия миозина X в опухолях способствует агрессивности рака молочной железы и метастазированию» . Британский журнал рака . 111 (3): 539–50. дои : 10.1038/bjc.2014.298 . ПМК 4119973 . ПМИД 24921915 .
- ^ Jump up to: а б Кенчаппа Р.С., Мистриотис П., Вишневски Е., Бхаттачарья С., Кулкарни Т., Вест Р. и др. (декабрь 2020 г.). «Миозин 10 регулирует инвазию, митоз и метаболическую передачу сигналов при глиобластоме» . iScience . 23 (12): 101802. Бибкод : 2020iSci...23j1802K . doi : 10.1016/j.isci.2020.101802 . ПМК 7702012 . ПМИД 33299973 .
- ^ Jump up to: а б Саммербелл Э.Р., Моу Дж.К., Белл Дж.С., Книпплер С.М., Педро Б., Арнст Дж.Л. и др. (июль 2020 г.). «Эпигенетически гетерогенные опухолевые клетки направляют коллективную инвазию посредством микропаттерна фибронектина, управляемого филоподиями» . Достижения науки . 6 (30): eaaz6197. Бибкод : 2020SciA....6.6197S . дои : 10.1126/sciadv.aaz6197 . ПМК 7439406 . ПМИД 32832657 .
- ^ Jump up to: а б с Хирано Ю., Хатано Т., Такахаши А., Торияма М., Инагаки Н. и Хакосима Т. (2011). Структурная основа распознавания груза доменом миозина-X MyTH4-FERM. Журнал EMBO 30 , 2734–2747.
- ^ Jump up to: а б с Лу, К., Ю, Дж., Ян, Дж., Вэй, З. и Чжан, М. (2011). Структурная основа тандема миозина X PH1(N)-PH2-PH1(C) как специфического и острого клеточного сенсора PI(3,4,5)P(3). Мол Биол Ячейка 22 , 4268-4278.
- ^ Jump up to: а б Вэй З, Ян Дж, Лу Ц, Пан Л, Чжан М (март 2011 г.). «Механизм распознавания грузов миозина X, выявленный по структуре его хвоста MyTH4-FERM в комплексе с доменом DCC P3» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (9): 3572–7. Бибкод : 2011PNAS..108.3572W . дои : 10.1073/pnas.1016567108 . ПМК 3048157 . ПМИД 21321230 .
- ^ Кербер М.Л., Чейни Р.Э. (ноябрь 2011 г.). «Миозин-X: миозин MyTH-FERM на кончиках филоподий» . Журнал клеточной науки . 124 (Часть 22): 3733–41. дои : 10.1242/jcs.023549 . ПМЦ 3225264 . ПМИД 22124140 .
- ^ Jump up to: а б Хомма К., Сайто Дж., Икебе Р., Икебе М. (сентябрь 2001 г.). «Двигательная функция и регуляция миозина Х» . Журнал биологической химии . 276 (36): 34348–54. дои : 10.1074/jbc.M104785200 . ПМИД 11457842 . S2CID 46029622 .
- ^ Jump up to: а б Такаги Ю., Фэрроу Р.Э., Биллингтон Н., Надь А., Баттерс С., Ян Ю. и др. (май 2014 г.). «Миозин-10 производит свой рабочий ход в две фазы и последовательно движется по одной актиновой нити при низкой нагрузке» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (18): E1833-42. Бибкод : 2014PNAS..111E1833T . дои : 10.1073/pnas.1320122111 . ПМК 4020102 . ПМИД 24753602 .
- ^ Карид Эй.Дж., Беннетт Р.Д., Стрелер Э.Э. (сентябрь 2010 г.). «Кинетический анализ выявляет различия в механизме связывания кальмодулина и кальмодулинподобного белка с мотивами IQ миозина-10» . Биохимия . 49 (37): 8105–16. дои : 10.1021/bi100644q . ПМЦ 3818122 . ПМИД 20731332 .
- ^ Найт П.Дж., Тирумуруган К., Сюй Ю., Ван Ф., Калверда А.П., Стаффорд В.Ф. и др. (октябрь 2005 г.). «Предсказанный спиральный домен миозина 10 образует новый удлиненный домен, который удлиняет голову» . Журнал биологической химии . 280 (41): 34702–8. дои : 10.1074/jbc.M504887200 . ПМИД 16030012 . S2CID 22503825 .
- ^ Jump up to: а б Лу Ц, Е Ф, Вэй Цз, Вэнь Цз, Чжан М (октябрь 2012 г.). «Антипараллельная спирально-опосредованная димеризация миозина X» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (43): 17388–93. дои : 10.1073/pnas.1208642109 . ПМЦ 3491486 . ПМИД 23012428 .
- ^ Jump up to: а б с д и Ропарс В., Ян З., Изабет Т., Блан Ф., Чжоу К., Лин Т. и др. (сентябрь 2016 г.). «Мотор миозина X оптимизирован для движения по актиновым пучкам» . Природные коммуникации . 7 : 12456. Бибкод : 2016NatCo...712456R . дои : 10.1038/ncomms12456 . ПМК 5025751 . ПМИД 27580874 .
- ^ Плантар Л., Арйонен А., Лок Дж.Г., Нурани Г., Иваска Дж., Стрёмблад С. (октябрь 2010 г.). «PtdIns(3,4,5)P₃ является регулятором локализации миозина-X и образования филоподий» . Журнал клеточной науки . 123 (Часть 20): 3525–34. дои : 10.1242/jcs.069609 . ПМИД 20930142 . S2CID 2070375 .
- ^ Jump up to: а б Умеки Н., Юнг Х.С., Сакаи Т., Сато О., Икебе Р., Икебе М. (июнь 2011 г.). «Фосфолипидзависимая регуляция двигательной активности миозина Х». Структурная и молекулярная биология природы . 18 (7): 783–8. дои : 10.1038/nsmb.2065 . ПМИД 21666676 . S2CID 9755324 .
- ^ Jump up to: а б Вебер К.Л., Сокач А.М., Берг Дж.С., Чейни Р.Э., Бемент В.М. (сентябрь 2004 г.). «Миозин, связывающий микротрубочки, необходимый для ядерного закрепления и сборки веретена». Природа . 431 (7006): 325–9. Бибкод : 2004Natur.431..325W . дои : 10.1038/nature02834 . ПМИД 15372037 . S2CID 4342360 .
- ^ Планельес-Эрреро В.Дж., Блан Ф., Сиригу С., Сиркиа Х., Клауз Дж., Суриг И. и др. (май 2016 г.). «Структуры миозина MyTH4-FERM подчеркивают важные принципы конвергентной эволюции» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (21): E2906-15. Бибкод : 2016PNAS..113E2906P . дои : 10.1073/pnas.1600736113 . ПМЦ 4889382 . ПМИД 27166421 .
- ^ Чжан Х., Берг Дж.С., Ли З., Ван Ю., Лонг П., Соуза А.Д. и др. (июнь 2004 г.). «Миозин-X обеспечивает моторную связь между интегринами и цитоскелетом». Природная клеточная биология . 6 (6): 523–31. дои : 10.1038/ncb1136 . ПМИД 15156152 . S2CID 24260820 .
- ^ Чжу XJ, Ван ЧЗ, Дай П.Г., Се Ю, Сун Н.Н., Лю Ю и др. (февраль 2007 г.). «Миозин X регулирует рецепторы нетрина и участвует в поиске аксонального пути». Природная клеточная биология . 9 (2): 184–92. дои : 10.1038/ncb1535 . ПМИД 17237772 . S2CID 7012749 .
- ^ Хомма К., Икебе М. (август 2005 г.). «Миозин X — двигатель с высоким коэффициентом полезного действия» . Журнал биологической химии . 280 (32): 29381–91. дои : 10.1074/jbc.M504779200 . ПМИД 15961399 . S2CID 22330459 .
- ^ Броули CM, Rock RS (июнь 2009 г.). «Нетрадиционный транспорт миозина в клетках обнаруживает селективный актиновый цитоскелет» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (24): 9685–90. Бибкод : 2009PNAS..106.9685B . дои : 10.1073/pnas.0810451106 . ПМК 2701044 . ПМИД 19478066 .
- ^ Вавра К.С., Ся Ю, Rock RS (июнь 2016 г.). «Конкуренция между спирально-спиральными структурами и влияние на выбор пучка миозина-10» . Биофизический журнал . 110 (11): 2517–2527. Бибкод : 2016BpJ...110.2517V . дои : 10.1016/j.bpj.2016.04.048 . ПМК 4906270 . ПМИД 27276269 .
- ^ Jump up to: а б с Соуза А.Д., Берг Дж.С., Робертсон Б.В., Микер Р.Б., Чейни Р.Э. (январь 2006 г.). «Myo10 в мозге: регуляция развития, идентификация безголовой изоформы и динамика в нейронах» . Журнал клеточной науки . 119 (Часть 1): 184–94. дои : 10.1242/jcs.02726 . ПМИД 16371656 . S2CID 3085103 .
- ^ Jump up to: а б Рейнс А.Н., Нагдас С., Кербер М.Л., Чейни Р.Э. (июль 2012 г.). «Безголовый Myo10 является негативным регулятором полноразмерного Myo10 и подавляет рост аксонов в корковых нейронах» . Журнал биологической химии . 287 (30): 24873–83. дои : 10.1074/jbc.M112.369173 . ПМК 3408153 . ПМИД 22661706 .
- ^ Лай М, Ли Л, Цзюй Х, Ю Х, Чжу Икс (2013). «Клонирование, характеристика и анализ промотора мышиного гена Myo10». Нуклеозиды, нуклеотиды и нуклеиновые кислоты . 32 (7): 354–65. дои : 10.1080/15257770.2013.790551 . ПМИД 23742061 . S2CID 1942016 .
- ^ Jump up to: а б с Петерсен К.Дж., Гудсон Х.В., Артур А.Л., Люкстон Г.В., Худусс А., Титус М.А. (декабрь 2016 г.). «Миозины MyTH4-FERM играют древнюю и консервативную роль в формировании филопод» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (50): Е8059–Е8068. Бибкод : 2016PNAS..113E8059P . дои : 10.1073/pnas.1615392113 . ПМК 5167205 . ПМИД 27911821 .
- ^ Jump up to: а б с Век М.Л., Грега-Ларсон Н.Е., Тыска М.Дж. (февраль 2017 г.). «Миозины MyTH4-FERM в сборке и поддержании выступов на основе актина» . Современное мнение в области клеточной биологии . 44 : 68–78. дои : 10.1016/j.ceb.2016.10.002 . ПМК 5403704 . ПМИД 27836411 .
- ^ Jump up to: а б с Берг Дж.С., Чейни Р.Э. (март 2002 г.). «Миозин-X — это нетрадиционный миозин, обладающий внутрифилоподиальной подвижностью». Природная клеточная биология . 4 (3): 246–50. дои : 10.1038/ncb762 . ПМИД 11854753 . S2CID 22683743 .
- ^ Кербер М.Л., Джейкобс Д.Т., Кампаньола Л., Данн Б.Д., Инь Т., Соуза А.Д. и др. (июнь 2009 г.). «Новая форма подвижности филоподий, обнаруженная при визуализации миозина-X на уровне одиночных молекул» . Современная биология . 19 (11): 967–73. Бибкод : 2009CBio...19..967K . дои : 10.1016/j.cub.2009.03.067 . ПМК 2817954 . ПМИД 19398338 .
- ^ Ватанабэ Т.М., Токуо Х., Гонда К., Хигучи Х., Икебе М. (июнь 2010 г.). «Миозин-X индуцирует филоподии за счет механизма множественного удлинения» . Журнал биологической химии . 285 (25): 19605–14. дои : 10.1074/jbc.M109.093864 . ПМЦ 2885239 . ПМИД 20392702 .
- ^ Бабулал Т.Г., Машанов Г.И., Ненашева Т.А., Пекхэм М., Моллой Дж.Е. (октябрь 2016 г.). «Сочетание диффузии и активной транслокации локализует миозин 10 на кончике филоподий» . Журнал биологической химии . 291 (43): 22373–22385. дои : 10.1074/jbc.M116.730689 . ПМК 5077179 . ПМИД 27566544 .
- ^ Бохил А.Б., Робертсон Б.В., Чейни Р.Э. (август 2006 г.). «Миозин-X — это молекулярный мотор, который участвует в формировании филоподий» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (33): 12411–6. Бибкод : 2006PNAS..10312411B . дои : 10.1073/pnas.0602443103 . ПМЦ 1567893 . ПМИД 16894163 .
- ^ Токуо Х., Мабути К., Икебе М. (октябрь 2007 г.). «Двигательная активность миозина-X способствует сближению актиновых волокон на периферии клетки, что инициирует образование филоподий» . Журнал клеточной биологии . 179 (2): 229–38. дои : 10.1083/jcb.200703178 . ПМК 2064759 . ПМИД 17954606 .
- ^ Тоёсима Ф., Нисида Э. (март 2007 г.). «Интегрино-опосредованная адгезия ориентирует веретено параллельно субстрату EB1- и миозин-X-зависимым образом» . Журнал ЭМБО . 26 (6): 1487–98. дои : 10.1038/sj.emboj.7601599 . ПМЦ 1829369 . ПМИД 17318179 .
- ^ Jump up to: а б с Квон М., Багонис М., Данузер Г., Пеллман Д. (август 2015 г.). «Прямое связывание микротрубочек миозином-10 ориентирует центросомы к ретракционным волокнам и подкорковым актиновым облакам» . Развивающая клетка . 34 (3): 323–37. дои : 10.1016/j.devcel.2015.06.013 . ПМЦ 4672950 . ПМИД 26235048 .
- ^ Сандквист Дж.К., Ларсон М.Э., Вулнер С., Дин З., Бемент В.М. (март 2018 г.). «Взаимодействие между миозином-10 и регулятором клеточного цикла Wee1 связывает динамику веретена с митотической прогрессией в эпителии» . Журнал клеточной биологии . 217 (3): 849–859. дои : 10.1083/jcb.201708072 . ПМЦ 5839792 . ПМИД 29321170 .
- ^ Шумахер М., Гольдман Р.Д., Лувард Д., Виньевич Д.М. (май 2010 г.). «Актин, микротрубочки и промежуточные филаменты виментина взаимодействуют для удлинения инвадоподий» . Журнал клеточной биологии . 189 (3): 541–56. дои : 10.1083/jcb.200909113 . ПМЦ 2867303 . ПМИД 20421424 .
- ^ Сунь Ю, Ай Икс, Шен С, Лу С (апрель 2015 г.). «NF-κB-опосредованная миР-124 подавляет метастазирование немелкоклеточного рака легких путем нацеливания на MYO10» . Онкотаргет . 6 (10): 8244–54. дои : 10.18632/oncotarget.3135 . ПМЦ 4480748 . ПМИД 25749519 .
- ^ Ван X, Ли Дж, Сюй X, Чжэн Дж, Ли Ц (июль 2018 г.). «МиР-129 ингибирует рост опухоли и усиливает химиочувствительность нейробластомы путем нацеливания на MYO10». Биомедицина и фармакотерапия . 103 : 1312–1318. дои : 10.1016/j.biopha.2018.04.153 . ПМИД 29864913 . S2CID 46926904 .
Дальнейшее чтение
[ редактировать ]- Хэссон Т., Скоурон Дж.Ф., Гилберт Д.Д., Авраам К.Б., Перри В.Л., Бемент В.М. и др. (сентябрь 1996 г.). «Картирование нетрадиционных миозинов у мышей и человека» . Геномика . 36 (3): 431–9. дои : 10.1006/geno.1996.0488 . ПМИД 8884266 .
- Нагасе Т., Исикава К., Суяма М., Кикуно Р., Миядзима Н., Танака А. и др. (октябрь 1998 г.). «Прогнозирование кодирующих последовательностей неопознанных генов человека. XI. Полные последовательности 100 новых клонов кДНК головного мозга, которые кодируют большие белки in vitro» . Исследование ДНК . 5 (5): 277–86. дои : 10.1093/dnares/5.5.277 . ПМИД 9872452 .
- Рохас К., Серрано де ла Пенья Л., Галлардо Т., Симмонс А., Найс К., МакГрат Р. и др. (декабрь 1999 г.). «Физическая карта и характеристика транскриптов в интервале-кандидате семейного хондрокальциноза на хромосоме 5p15.1». Геномика . 62 (2): 177–83. дои : 10.1006/geno.1999.5997 . ПМИД 10610710 .
- Роджерс М.С., Стрелер Э.Э. (апрель 2001 г.). «Опухолечувствительный кальмодулинподобный белок представляет собой специфическую легкую цепь нетрадиционного человеческого миозина X» . Журнал биологической химии . 276 (15): 12182–9. дои : 10.1074/jbc.M010056200 . ПМИД 11278607 .
- Кокс Д., Берг Дж.С., Каммер М., Чинегвундо Д.О., Дейл Б.М., Чейни Р.Э., Гринберг С. (июль 2002 г.). «Миозин X является нижестоящим эффектором PI (3) K во время фагоцитоза». Природная клеточная биология . 4 (7): 469–77. дои : 10.1038/ncb805 . ПМИД 12055636 . S2CID 24127183 .
- Наварро-Лерида I, Мартинес Морено М, Ронкаль Ф, Гавиланес Ф, Альбар ХП, Родригес-Креспо I (февраль 2004 г.). «Протеомная идентификация белков головного мозга, которые взаимодействуют с легкой цепью динеина LC8». Протеомика . 4 (2): 339–46. дои : 10.1002/pmic.200300528 . ПМИД 14760703 . S2CID 8868600 .
- Чжан Х., Берг Дж.С., Ли З., Ван Ю., Лонг П., Соуза А.Д. и др. (июнь 2004 г.). «Миозин-X обеспечивает моторную связь между интегринами и цитоскелетом». Природная клеточная биология . 6 (6): 523–31. дои : 10.1038/ncb1136 . ПМИД 15156152 . S2CID 24260820 .
- Токуо Х., Икебе М. (июнь 2004 г.). «Миозин X транспортирует Mena/VASP к кончику филоподий». Связь с биохимическими и биофизическими исследованиями . 319 (1): 214–20. дои : 10.1016/j.bbrc.2004.04.167 . ПМИД 15158464 .
- Кимура К., Вакамацу А., Судзуки Й., Ота Т., Нишикава Т., Ямашита Р. и др. (январь 2006 г.). «Диверсификация транскрипционной модуляции: крупномасштабная идентификация и характеристика предполагаемых альтернативных промоторов генов человека» . Геномные исследования . 16 (1): 55–65. дои : 10.1101/гр.4039406 . ПМЦ 1356129 . ПМИД 16344560 .
- Соуза А.Д., Берг Дж.С., Робертсон Б.В., Микер Р.Б., Чейни Р.Э. (январь 2006 г.). «Myo10 в мозге: регуляция развития, идентификация безголовой изоформы и динамика в нейронах» . Журнал клеточной науки . 119 (Часть 1): 184–94. дои : 10.1242/jcs.02726 . ПМИД 16371656 .
- Бохил А.Б., Робертсон Б.В., Чейни Р.Э. (август 2006 г.). «Миозин-X — это молекулярный мотор, который участвует в формировании филоподий» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (33): 12411–6. Бибкод : 2006PNAS..10312411B . дои : 10.1073/pnas.0602443103 . ПМЦ 1567893 . ПМИД 16894163 .
- Олсен Дж.В., Благоев Б., Гнад Ф., Мачек Б., Кумар С., Мортенсен П., Манн М. (ноябрь 2006 г.). «Глобальная, in vivo и сайт-специфическая динамика фосфорилирования в сигнальных сетях» . Клетка . 127 (3): 635–48. дои : 10.1016/j.cell.2006.09.026 . ПМИД 17081983 . S2CID 7827573 .