Виментин
| ПОЧЕМУ | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | ВИМ , CTRCT30, HEL113, виментин | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 193060 ; МГИ : 98932 ; Гомологен : 2538 ; Генные карты : ВИМ ; ОМА : ВИМ – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




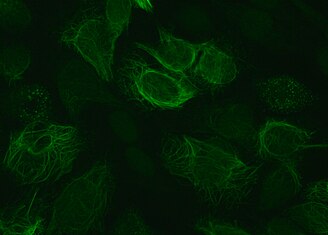
Виментин — структурный белок , который у человека кодируется VIM геном . Его название происходит от латинского vimentum , обозначающего набор гибких стержней. [5]

Виментин представляет собой белок промежуточных филаментов (ПФ) III типа , который экспрессируется в мезенхимальных клетках. Белки IF присутствуют во всех клетках животных. [6] а также бактерии . [7] Промежуточные филаменты, наряду с тубулина на основе микротрубочками и актина на основе микрофиламентами , составляют цитоскелет . Все белки IF экспрессируются строго регулируемым в процессе развития способом; Виментин является основным компонентом цитоскелета мезенхимальных клеток. По этой причине виментин часто используется в качестве маркера клеток мезенхимального происхождения или клеток, подвергающихся эпителиально-мезенхимальному переходу (ЕМТ) как во время нормального развития, так и в ходе метастатического прогрессирования.
Структура
[ редактировать ]Сборка фиброзной нити виментина, образующей цитоскелет, происходит постепенно. Мономер виментина имеет центральный α-спиральный домен , кэпированный с каждого конца неспиральными амино- (головным) и карбоксильным (хвостовым) доменами. [8] Два мономера, вероятно, котрансляционно экспрессируются таким образом, что облегчает их взаимодействие с образованием спирально-спирального димера, который является основной субъединицей сборки виментина. [9] Пара спиральных димеров соединяются антипараллельно, образуя тетрамер. Восемь тетрамеров соединяются, образуя так называемую нить единичной длины (ULF), затем ULF прилипают друг к другу и удлиняются с последующим уплотнением с образованием волокнистых белков. [10]
α-спиральные последовательности содержат набор гидрофобных аминокислот, которые способствуют образованию «гидрофобного уплотнения» на поверхности спирали. [8] Кроме того, существует периодическое распределение кислых и основных аминокислот , которое, по-видимому, играет важную роль в стабилизации спиральных димеров. [8] Расстояние между заряженными остатками оптимально для ионных солевых мостиков , что позволяет стабилизировать структуру α-спирали. Хотя этот тип стабилизации интуитивно понятен для внутрицепных взаимодействий, а не для межцепных взаимодействий, ученые предположили, что, возможно, переход от внутрицепочечных солевых мостиков, образованных кислотными и основными остатками, к межцепочечным ионным ассоциациям способствует сборке нити. [8]
Функция
[ редактировать ]Виментин играет значительную роль в поддержании и закреплении положения органелл в цитозоле . Виментин прикрепляется к ядру , эндоплазматической сети и митохондриям латерально или терминально. [11]
Динамическая природа виментина важна для обеспечения гибкости клетки. Ученые обнаружили, что виментин обеспечивает клеткам устойчивость, отсутствующую в сетях микротрубочек или актиновых филаментов, при механическом стрессе in vivo . Поэтому в целом принято считать, что виментин является компонентом цитоскелета, ответственным за поддержание целостности клеток. (Было обнаружено, что клетки без виментина чрезвычайно чувствительны при повреждении микропунктурой). [12] Трансгенные мыши, у которых отсутствует виментин, выглядели нормальными и не имели функциональных различий. [13] Возможно, сеть микротрубочек компенсировала отсутствие промежуточной сети. Этот результат подтверждает тесное взаимодействие между микротрубочками и виментином. Более того, когда присутствовали деполимеризаторы микротрубочек, происходила реорганизация виментина, что еще раз указывает на связь между двумя системами. [12] С другой стороны, раненые мыши, у которых отсутствует ген виментина, заживают медленнее, чем их собратья дикого типа. [14]
По сути, виментин отвечает за поддержание формы клеток, целостности цитоплазмы и стабилизацию цитоскелетных взаимодействий. Было показано, что виментин устраняет токсичные белки из JUNQ и IPOD телец включения при асимметричном делении клеточных линий млекопитающих . [15]
Кроме того, обнаружено, что виментин контролирует транспорт , полученного из липопротеинов низкой плотности (ЛПНП), холестерина из лизосомы к месту этерификации. [16] Было обнаружено, что при блокировании транспорта холестерина, полученного из ЛПНП, клетки хранят гораздо меньший процент липопротеинов, чем нормальные клетки с виментином. Эта зависимость, по-видимому, является первым процессом биохимической функции любой клетки, который зависит от сети клеточных промежуточных филаментов. Этот тип зависимости имеет последствия для клеток надпочечников, которые полагаются на эфиры холестерина, полученные из ЛПНП. [16]
Виментин играет роль в формировании агресомы , где он образует клетку, окружающую ядро агрегированного белка. [17]
Помимо традиционной внутриклеточной локализации, виментин можно обнаружить и внеклеточно. Виментин может экспрессироваться как белок клеточной поверхности и, как предполагается, играет роль в иммунных реакциях. Он также может высвобождаться в фосфорилированных формах во внеклеточное пространство активированными макрофагами , известно также, что астроциты выделяют виментин. [18]
Клиническое значение
[ редактировать ]Его использовали в качестве саркомы опухолевого маркера для идентификации мезенхимы . [19] [20] Его специфичность как биомаркера оспаривается Джерадом Гарднером. [21] Виментин присутствует при веретеноклеточном плоскоклеточном раке. [22] [23]
Метилирование гена виментина было установлено как биомаркер рака толстой кишки, и это используется при разработке фекальных тестов на рак толстой кишки. Статистически значимые уровни метилирования гена виментина также наблюдались при некоторых патологиях верхних отделов желудочно-кишечного тракта, таких как пищевод Барретта , аденокарцинома пищевода и рак желудка кишечного типа. [24] Высокие уровни метилирования ДНК в промоторной области также были связаны со значительным снижением выживаемости при гормонопозитивном раке молочной железы. [25] было выявлено снижение уровня виментина при кистозном варианте папиллярного рака щитовидной железы . С помощью протеомного подхода [26] См. также антитела к цитруллинированному белку для использования в диагностике ревматоидного артрита .
обнаружили, что виментин является фактором прикрепления SARS-CoV-2 . Надер Рахими и его коллеги [27]
Взаимодействия
[ редактировать ]Было показано, что виментин взаимодействует с:
- ЦСП [28]
- МУЖЧИНЫ1 [29]
- МИСТ2 [30] [31]
- ПКН1 [32]
- ПРККИ [33] [34]
- ОСТАВЛЯТЬ [35] [36]
- СПТАН1 [36]
- УПП1 [37]
- ЯВАЗ [38]
кДа . Было обнаружено, что 3'-UTR мРНК виментина связывает белок массой 46 [39]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000026025 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000026728 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Франке В.В., Шмид Э., Осборн М., Вебер К. (октябрь 1978 г.). «Различные нити среднего размера, выявленные с помощью иммунофлуоресцентной микроскопии» . Труды Национальной академии наук Соединенных Штатов Америки . 75 (10): 5034–5038. Бибкод : 1978PNAS...75.5034F . дои : 10.1073/pnas.75.10.5034 . ПМК 336257 . ПМИД 368806 .
- ^ Эрикссон Дж.Э., Дечат Т., Грин Б., Хелфанд Б., Мендес М., Паллари Х.М., Голдман Р.Д. (июль 2009 г.). «Введение промежуточных филаментов: от открытия к болезни» . Журнал клинических исследований . 119 (7): 1763–1771. дои : 10.1172/JCI38339 . ПМК 2701876 . ПМИД 19587451 .
- ^ Кабин М.Т., Джейкобс-Вагнер С. (2010). «Бактериальный цитоскелет». Ежегодный обзор генетики . 44 : 365–392. doi : 10.1146/annurev-genet-102108-134845 . ПМИД 21047262 .
- ^ Перейти обратно: а б с д Фукс Э, Вебер К (1994). «Промежуточные нити: структура, динамика, функции и заболевания». Ежегодный обзор биохимии . 63 : 345–382. дои : 10.1146/annurev.bi.63.070194.002021 . ПМИД 7979242 .
- ^ Чанг Л., Шав-Тал Ю., Трчек Т., Сингер Р.Х., Голдман Р.Д. (февраль 2006 г.). «Сборка сети промежуточных филаментов путем динамической котрансляции» . Журнал клеточной биологии . 172 (5): 747–758. дои : 10.1083/jcb.200511033 . ПМК 2063706 . ПМИД 16505169 .
- ^ Дэниэлссон Ф, Петерсон МК, Кальдейра Араужо Х, Лаутеншлегер Ф, Гад АК (сентябрь 2018 г.). «Разнообразие Виментина в здоровье и болезнях» . Клетки . 7 (10): 147. doi : 10.3390/cells7100147 . ПМК 6210396 . ПМИД 30248895 .
- ^ Кацумото Т., Мицусима А., Куримура Т. (1990). «Роль промежуточных филаментов виментина в клетках 3Y1 крысы, выясненная с помощью иммуноэлектронной микроскопии и компьютерной графической реконструкции». Биология клетки . 68 (2): 139–146. дои : 10.1016/0248-4900(90)90299-I . ПМИД 2192768 . S2CID 29019928 .
- ^ Перейти обратно: а б Гольдман Р.Д., Хуон С., Чоу Ю.Х., Опал П., Штайнерт П.М. (август 1996 г.). «Функция промежуточных филаментов в форме клеток и целостности цитоскелета» . Журнал клеточной биологии . 134 (4): 971–983. дои : 10.1083/jcb.134.4.971 . ПМК 2120965 . ПМИД 8769421 .
- ^ Колуччи-Гийон Э., Портье М.М., Дуния И., Полен Д., Пурнен С., Бабине С. (ноябрь 1994 г.). «Мыши, лишенные виментина, развиваются и размножаются без явного фенотипа». Клетка . 79 (4): 679–694. дои : 10.1016/0092-8674(94)90553-3 . ПМИД 7954832 . S2CID 28146121 .
- ^ Экес Б., Колуччи-Гийон Э., Смола Х., Ноддер С., Бабине С., Криг Т., Мартин П. (июль 2000 г.). «Нарушение заживления ран у эмбриональных и взрослых мышей, лишенных виментина». Журнал клеточной науки . 113 (13): 2455–2462. дои : 10.1242/jcs.113.13.2455 . ПМИД 10852824 .
- ^ Огродник М., Салмонович Х., Браун Р., Турковска Дж., Среднява В., Паттабираман С. и др. (июнь 2014 г.). «Динамические тельца включения JUNQ асимметрично наследуются в клеточных линиях млекопитающих за счет асимметричного разделения виментина» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (22): 8049–8054. Бибкод : 2014PNAS..111.8049O . дои : 10.1073/pnas.1324035111 . ПМК 4050583 . ПМИД 24843142 .
- ^ Перейти обратно: а б Саррия А.Дж., Панини С.Р., Эванс Р.М. (сентябрь 1992 г.). «Функциональная роль промежуточных филаментов виментина в метаболизме холестерина, полученного из липопротеинов, в клетках SW-13 человека» . Журнал биологической химии . 267 (27): 19455–19463. дои : 10.1016/S0021-9258(18)41797-8 . ПМИД 1527066 .
- ^ Джонстон Дж. А., Уорд К. Л., Копито Р. Р. (декабрь 1998 г.). «Агресомы: клеточный ответ на неправильно свернутые белки» . Журнал клеточной биологии . 143 (7): 1883–1898. дои : 10.1083/jcb.143.7.1883 . ПМК 2175217 . ПМИД 9864362 .
- ^ Полин Д., Лилиенбаум А., Карджян С., Агбулут О., Ли З. (июнь 2022 г.). «Виментин: Регуляция и патогенез» . Биохимия . 197 : 96–112. дои : 10.1016/j.biochi.2022.02.003 . ПМИД 35151830 . S2CID 246778685 .
- ^ Лидер М., Коллинз М., Патель Дж., Генри К. (январь 1987 г.). «Виментин: оценка его роли как онкомаркера». Гистопатология . 11 (1): 63–72. дои : 10.1111/j.1365-2559.1987.tb02609.x . ПМИД 2435649 . S2CID 34804720 .
- ^ «Иммуногистохимия Вашингтонской лаборатории диагностики болезней животных (WADDL) Колледжа ветеринарной медицины Вашингтонского государственного университета» . Архивировано из оригинала 1 декабря 2008 г. Проверено 14 марта 2009 г.
- ^ Гарднер Дж. (23 сентября 2010 г.). «Как интерпретировать иммуноокрашивание виментином» . Ютуб . Архивировано из оригинала 12 декабря 2021 г.
- ^ Дотто, Дж. Э.; Глусак, Э.Дж. (2006). «Р63 является полезным маркером веретеноклеточного плоскоклеточного рака кожи» . Журнал кожной патологии . 33 (6): 413–417. дои : 10.1111/j.0303-6987.2006.00477.x . ПМИД 16776716 . S2CID 38273016 .
- ^ Барретт, К.; Ван Кельст, С.; Воутерс, Дж.; Марасиган, В.; Фьювс, С.; Агостинис, П.; Ван Ден Оорд, Дж.; Гармин, М. (2014). «Эпителиально-мезенхимальный переход во время инвазии плоскоклеточного рака кожи сопровождается активацией АКТ» . Британский журнал дерматологии . 171 (5): 1014–1021. дои : 10.1111/bjd.12967 . ПМИД 24628329 . S2CID 42440330 .
- ^ Мойнова Х., Лейднер Р.С., Рави Л., Латтербо Дж., Барнхольц-Слоан Дж.С., Чен Ю. и др. (апрель 2012 г.). «Аберрантное метилирование виментина характерно для патологий верхних отделов желудочно-кишечного тракта» . Эпидемиология рака, биомаркеры и профилактика . 21 (4): 594–600. дои : 10.1158/1055-9965.EPI-11-1060 . ПМЦ 3454489 . ПМИД 22315367 .
- ^ Улирш Дж., Фан С., Кнафл Г., Ву М.Дж., Коулман Б., Перу С.М., Свифт-Сканлан Т. (январь 2013 г.). «Метилирование ДНК виментина предсказывает выживаемость при раке молочной железы» . Исследование и лечение рака молочной железы . 137 (2): 383–396. дои : 10.1007/s10549-012-2353-5 . ПМЦ 3838916 . ПМИД 23239149 .
- ^ Динец А., Пернемальм М., Челлин Х., Святоха В., Софиадис А., Юлин CC и др. (2015). «Дифференциальные профили экспрессии белка в кистозной жидкости папиллярной карциномы щитовидной железы и доброкачественных поражений щитовидной железы» . ПЛОС ОДИН . 10 (5): e0126472. Бибкод : 2015PLoSO..1026472D . дои : 10.1371/journal.pone.0126472 . ПМЦ 4433121 . ПМИД 25978681 .
- ^ Амраи Р., Ся С., Олейник Дж., Уайт М.Р., Наполеон М.А., Лотфоллахзаде С. и др. (февраль 2022 г.). «Внеклеточный виментин является фактором прикрепления, который облегчает проникновение SARS-CoV-2 в эндотелиальные клетки человека» . Труды Национальной академии наук Соединенных Штатов Америки . 119 (6). Бибкод : 2022PNAS..11913874A . дои : 10.1073/pnas.2113874119 . ПМЦ 8833221 . ПМИД 35078919 . e2113874119.
- ^ Мэн Дж.Дж., Борнслагер Э.А., Грин К.Дж., Штайнерт П.М., Ип В. (август 1997 г.). «Двугибридный анализ выявляет фундаментальные различия в прямых взаимодействиях между десмоплакином и промежуточными филаментами, специфичными для типа клеток» . Журнал биологической химии . 272 (34): 21495–21503. дои : 10.1074/jbc.272.34.21495 . ПМИД 9261168 .
- ^ Лопес-Эгидо Дж., Каннингем Дж., Берг М., Оберг К., Бонгкам-Рудлофф Э., Гобль А. (август 2002 г.). «Взаимодействие менина с глиальным фибриллярным кислым белком и виментином предполагает роль сети промежуточных филаментов в регуляции активности менина». Экспериментальные исследования клеток . 278 (2): 175–183. дои : 10.1006/excr.2002.5575 . ПМИД 12169273 .
- ^ Руал Дж. Ф., Венкатесан К., Хао Т., Хиродзан-Кисикава Т., Дрико А., Ли Н. и др. (октябрь 2005 г.). «К карте сети белок-белковых взаимодействий человека в масштабе протеома». Природа . 437 (7062): 1173–1178. Бибкод : 2005Natur.437.1173R . дои : 10.1038/nature04209 . ПМИД 16189514 . S2CID 4427026 .
- ^ Стельцль У., Ворм У., Лаловски М., Хениг С., Брембек Ф.Х., Гёлер Х. и др. (сентябрь 2005 г.). «Сеть белок-белкового взаимодействия человека: ресурс для аннотирования протеома». Клетка . 122 (6): 957–968. дои : 10.1016/j.cell.2005.08.029 . hdl : 11858/00-001M-0000-0010-8592-0 . ПМИД 16169070 . S2CID 8235923 .
- ^ Мацудзава К., Косако Х., Инагаки Н., Сибата Х., Мукаи Х., Оно Ю. и др. (май 1997 г.). «Доменоспецифическое фосфорилирование виментина и глиального фибриллярного кислого белка с помощью PKN». Связь с биохимическими и биофизическими исследованиями . 234 (3): 621–625. дои : 10.1006/bbrc.1997.6669 . ПМИД 9175763 .
- ^ Ратнаяке В.С., Апостолатос А.Х., Остров Д.А., Асеведо-Дункан М. (ноябрь 2017 г.). «Два новых атипичных ингибитора ПКС: ACPD и DNDA эффективно уменьшают пролиферацию клеток и эпителиально-мезенхимальный переход метастатической меланомы, одновременно индуцируя апоптоз» . Международный журнал онкологии . 51 (5): 1370–1382. дои : 10.3892/ijo.2017.4131 . ПМЦ 5642393 . ПМИД 29048609 .
- ^ Ратнаяке В.С., Апостолатос К.А., Апостолатос А.Х., Шутте Р.Дж., Хюнь М.А., Остров Д.А., Асеведо-Дункан М. (2018). «Онкогенная PKC-ι активирует виментин во время эпителиально-мезенхимального перехода при меланоме; исследование, основанное на специфических ингибиторах PKC-ι и PKC-ζ» . Адгезия и миграция клеток . 12 (5): 447–463. дои : 10.1080/19336918.2018.1471323 . ПМК 6363030 . ПМИД 29781749 .
- ^ Херрманн Х., Виче Г. (январь 1987 г.). «Плектин и IFAP-300K являются гомологичными белками, связывающимися с белками 1 и 2, связанными с микротрубочками, а также с субъединицей спектрина массой 240 килодальтон» . Журнал биологической химии . 262 (3): 1320–1325. дои : 10.1016/S0021-9258(19)75789-5 . ПМИД 3027087 .
- ^ Перейти обратно: а б Браун М.Дж., Халлам Дж.А., Лю Ю., Ямада К.М., Шоу С. (июль 2001 г.). «Авангард: интеграция цитоскелета Т-лимфоцитов человека с помощью цитолинкера плектина» . Журнал иммунологии . 167 (2): 641–645. дои : 10.4049/jimmunol.167.2.641 . ПМИД 11441066 .
- ^ Рассел Р.Л., Цао Д., Чжан Д., Хандшумахер Р.Э., Пиццорно Дж. (апрель 2001 г.). «Ассоциация уридинфосфорилазы с виментином. Внутриклеточное распределение и локализация» . Журнал биологической химии . 276 (16): 13302–13307. дои : 10.1074/jbc.M008512200 . ПМИД 11278417 .
- ^ Цивион Г., Луо З.Дж., Авруч Дж. (сентябрь 2000 г.). «Каликулин А-индуцированное фосфорилирование виментина секвестрирует 14-3-3 и вытесняет других партнеров 14-3-3 in vivo» . Журнал биологической химии . 275 (38): 29772–29778. дои : 10.1074/jbc.M001207200 . ПМИД 10887173 .
- ^ Зенер З.Е., Шеперд Р.К., Габришук Дж., Фу Т.Ф., Аль-Али М., Холмс В.М. (август 1997 г.). «РНК-белковые взаимодействия внутри 3'-нетранслируемой области мРНК виментина» . Исследования нуклеиновых кислот . 25 (16): 3362–3370. дои : 10.1093/нар/25.16.3362 . ПМК 146884 . ПМИД 9241253 .
Дальнейшее чтение
[ редактировать ]- Снасель Й, Пихова I (1997). «Расщепление белков клетки-хозяина протеазой ВИЧ-1». Фолиа биологическая . 42 (5): 227–230. дои : 10.1007/BF02818986 . ПМИД 8997639 . S2CID 7617882 .
- Лейк Дж.А., Карр Дж., Фэн Ф., Манди Л., Баррелл С., Ли П. (февраль 2003 г.). «Роль Vif во время инфекции ВИЧ-1: взаимодействие с новыми клеточными факторами хозяина». Журнал клинической вирусологии . 26 (2): 143–152. дои : 10.1016/S1386-6532(02)00113-0 . ПМИД 12600646 .
Внешние ссылки
[ редактировать ]галерея PDB |
|---|

