Цитоскелет может выполнять множество функций. Его основная функция — придать клетке форму и механическую устойчивость к деформации, а за счет ассоциации с внеклеточной соединительной тканью и другими клетками он стабилизирует целые ткани. [4] [5] Цитоскелет также может сокращаться, тем самым деформируя клетку и ее среду и позволяя клеткам мигрировать . [6] Более того, он участвует во многих клеточных сигнальных путях и в поглощении внеклеточного материала ( эндоцитоз ). [7] расслоение хромосом при делении клетки , [4] стадия цитокинеза деления клеток, [8] как каркас для организации содержимого клетки в пространстве [6] и во внутриклеточном транспорте (например, перемещение везикул и органелл внутри клетки) [4] и может служить шаблоном для построения клеточной стенки . [4] Кроме того, он может образовывать специализированные структуры, такие как жгутики , реснички , ламеллиподии и подосомы . Структура, функции и динамическое поведение цитоскелета могут сильно различаться в зависимости от организма и типа клеток. [4] [9] [8] Даже внутри одной клетки цитоскелет может меняться за счет ассоциации с другими белками и предыдущей истории сети. [6]
Масштабным примером действия цитоскелета является сокращение мышц . Это осуществляется совместными усилиями групп узкоспециализированных клеток. Основным компонентом цитоскелета, который помогает показать истинную функцию мышечного сокращения, являются микрофиламенты . Микрофиламенты состоят из наиболее распространенного клеточного белка, известного как актин. [10] Во время сокращения мышцы внутри каждой мышечной клетки молекулярные моторы миозина коллективно воздействуют на параллельные актиновые нити. Сокращение мышц начинается с нервных импульсов, которые затем вызывают высвобождение повышенного количества кальция из саркоплазматической сети . Увеличение содержания кальция в цитозоле позволяет начать сокращение мышц с помощью двух белков, тропомиозина и тропонина . [10] Тропомиозин ингибирует взаимодействие между актином и миозином, тогда как тропонин ощущает увеличение кальция и снимает это торможение. [11] Это действие сокращает мышечную клетку, а благодаря синхронному процессу во многих мышечных клетках – всю мышцу.
В 1903 году Николай К. Кольцов предположил, что форма клеток определяется сетью канальцев, которую он назвал цитоскелетом. Идея белковой мозаики, динамически координирующей биохимию цитоплазмы, была предложена Рудольфом Петерсом в 1929 году. [12] в то время как термин ( цитоскелетт , по-французски) был впервые введен французским эмбриологом Полем Винтребертом в 1931 году. [13]
Когда цитоскелет был впервые представлен, считалось, что это неинтересное гелеобразное вещество, которое помогает органеллам оставаться на месте. [14] Было проведено много исследований, чтобы попытаться понять назначение цитоскелета и его компонентов.
Первоначально считалось, что цитоскелет свойственен только эукариотам, но в 1992 году было обнаружено, что он присутствует и у прокариот. Это открытие произошло после осознания того, что бактерии обладают белками, гомологичными тубулину и актину; Основные компоненты цитоскелета эукариот. [15]
Исследования нейродегенеративных заболеваний , таких как болезнь Паркинсона , болезнь Альцгеймера , болезнь Хантингтона и боковой амиотрофический склероз (БАС), показывают, что при этих заболеваниях поражается цитоскелет. [19] Болезнь Паркинсона характеризуется деградацией нейронов, что приводит к тремору, ригидности и другим немоторным симптомам. Исследования показали, что сборка и стабильность микротрубочек в цитоскелете нарушаются, что приводит к деградации нейронов с течением времени. [20] При болезни Альцгеймера тау-белки , которые стабилизируют микротрубочки, нарушаются при прогрессировании заболевания, вызывая патологию цитоскелета. [21] Предполагается, что избыток глютамина в белке Хантингтона, участвующем в связывании везикул с цитоскелетом, является фактором развития болезни Хантингтона. [22] Боковой амиотрофический склероз приводит к потере движения, вызванной деградацией мотонейронов, а также включает дефекты цитоскелета. [23]
Вспомогательные белки, включая моторные белки, регулируют и связывают филаменты с другими клеточными соединениями и друг с другом и необходимы для контролируемой сборки цитоскелетных филаментов в определенных местах. [26]
ряд низкомолекулярных цитоскелетных препаратов Обнаружен , взаимодействующих с актином и микротрубочками. Эти соединения оказались полезными при изучении цитоскелета, а некоторые из них нашли клиническое применение.
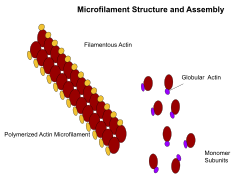
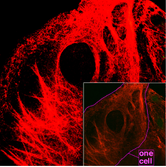
Микрофиламенты, также известные как актиновые нити, состоят из линейных полимеров белков G-актина и генерируют силу, когда растущий (плюсовой) конец нити прижимается к барьеру, например клеточной мембране. Они также действуют как треки для движения молекул миозина , которые прикрепляются к микрофиламентам и «ходят» вдоль них. Как правило, основным компонентом или белком микрофиламентов является актин. Мономер G-актина объединяется с образованием полимера, который продолжает формировать микрофиламент (актиновую нить). Эти субъединицы затем собираются в две цепи, которые переплетаются в так называемые цепи F-актина . [27] Движение миозина по нитям F-актина генерирует сократительные силы в так называемых актомиозиновых волокнах, как в мышцах, так и в большинстве типов немышечных клеток. [28] Структуры актина контролируются семейством Rho небольших GTP-связывающих белков, таких как сам Rho для сократительных нитей актомиозина («стрессовые волокна»), Rac для ламеллиподий и Cdc42 для филоподий.
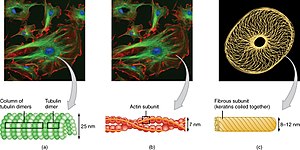
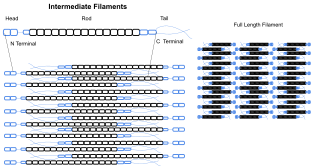
Промежуточные филаменты входят в состав цитоскелета многих эукариотических клеток. Эти нити, имеющие средний диаметр 10 нанометров, более стабильны (прочно связаны), чем микрофиламенты и гетерогенные составляющие цитоскелета. Подобно актиновым нитям, они поддерживают форму клеток, выдерживая напряжение ( микротрубочки , напротив, сопротивляются сжатию, но также могут выдерживать напряжение во время митоза и во время позиционирования центросомы). Промежуточные нити организуют внутреннюю трехмерную структуру клетки, закрепляют органеллы и служат структурными компонентами ядерной пластинки . Они также участвуют в некоторых межклеточных и межклеточных соединениях. Ядерная пластинка существует у всех животных и всех тканей. У некоторых животных, таких как плодовая мушка, нет цитоплазматических промежуточных нитей. У тех животных, которые экспрессируют цитоплазматические промежуточные филаменты, они тканеспецифичны. [5] Кератиновые промежуточные нити в эпителиальных клетках обеспечивают защиту от различных механических воздействий, которым может подвергаться кожа. Они также обеспечивают защиту органов от метаболических, окислительных и химических стрессов. Укрепление эпителиальных клеток этими промежуточными нитями может предотвратить начало апоптоза или гибель клеток за счет снижения вероятности стресса. [29]
Промежуточные филаменты чаще всего известны как система поддержки или «каркас» для клетки и ядра, а также играют роль в некоторых клеточных функциях. В сочетании с белками и десмосомами промежуточные нити образуют межклеточные связи и закрепляют соединения клетка-матрикс, которые используются для передачи сообщений между клетками, а также для жизненно важных функций клетки. Эти соединения позволяют клетке взаимодействовать через десмосому с несколькими клетками, чтобы корректировать структуры ткани на основе сигналов из окружающей среды клетки. Было показано, что мутации в белках IF вызывают серьезные медицинские проблемы, такие как преждевременное старение, мутации десмина, поражающие органы, болезнь Александера и мышечную дистрофию . [5]
Различные промежуточные нити:
из виментинов . Промежуточные филаменты виментина обычно присутствуют в мезенхимальных клетках.
изготовлен из кератина . Кератин в основном присутствует в эпителиальных клетках.
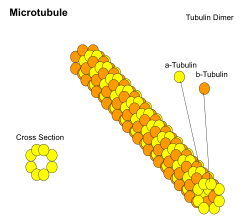
Микротрубочки представляют собой полые цилиндры диаметром около 23 нм (диаметр просвета около 15 нм), чаще всего состоящие из 13 протофиламентов , которые, в свою очередь, представляют собой полимеры альфа- и бета- тубулина . Они имеют очень динамичное поведение, связывая GTP для полимеризации. Они обычно организованы центросомой .
В девяти тройных наборах (звездчатых) они образуют центриоли , а в девяти дублетах, ориентированных вокруг двух дополнительных микротрубочек (колесообразных), они образуют реснички и жгутики. Последнее образование обычно называют структурой «9+2», в которой каждый дублет соединен с другим белком динеином . Поскольку и жгутики, и реснички являются структурными компонентами клетки и поддерживаются микротрубочками, их можно считать частью цитоскелета. Существует два типа ресничек: подвижные и неподвижные. Реснички короткие и более многочисленные, чем жгутики. Подвижные реснички совершают ритмичные волнистые или пульсирующие движения по сравнению с неподвижными ресничками, которые получают сенсорную информацию для клетки; обработка сигналов от других клеток или окружающих их жидкостей. Кроме того, микротрубочки контролируют биение (движение) ресничек и жгутиков. [31] Кроме того, динеиновые плечи, прикрепленные к микротрубочкам, действуют как молекулярные моторы. Движение ресничек и жгутиков создается за счет скольжения микротрубочек друг мимо друга, для чего требуется АТФ. [31] Они играют ключевую роль в:
Септины представляют собой группу высококонсервативных GTP- связывающих белков, обнаруженных у эукариот . Различные септины образуют белковые комплексы друг с другом . Они могут собираться в нити и кольца. Следовательно, септины можно рассматривать как часть цитоскелета. [36] Функция септинов в клетках включает в себя роль локализованного места прикрепления других белков и предотвращение диффузии определенных молекул из одного клеточного отсека в другой. [36] В дрожжевых клетках они строят каркас, обеспечивающий структурную поддержку во время клеточного деления и компартментализации частей клетки. Недавние исследования клеток человека показывают, что септины создают клетки вокруг бактериальных патогенов, обездвиживая вредные микробы и предотвращая их проникновение в другие клетки. [37]
У почкующихся дрожжей (важного модельного организма ) актин образует кортикальные участки, актиновые кабели, а также цитокинетическое кольцо и колпачок. Кортикальные участки представляют собой отдельные актиновые тельца на мембране и жизненно важны для эндоцитоза , особенно для рециркуляции глюкансинтазы, которая важна для синтеза клеточной стенки . Актиновые кабели представляют собой пучки актиновых нитей и участвуют в транспортировке везикул к колпачку (который содержит ряд различных белков, поляризующих клеточный рост), а также в расположении митохондрий. Цитокинетическое кольцо формируется и сужается вокруг места деления клеток . [39]
До работы Jones et al., 2001 считалось, что клеточная стенка является решающим фактором для многих форм бактериальных клеток, включая палочки и спирали. При изучении было обнаружено, что многие деформированные бактерии имеют мутации, связанные с развитием клеточной оболочки . [40] Когда-то считалось, что цитоскелет свойственен только эукариотическим клеткам, но были обнаружены гомологи всех основных белков эукариотического цитоскелета у прокариот . [41] Гарольд Эриксон отмечает, что до 1992 года считалось, что только эукариоты имеют компоненты цитоскелета. Однако исследования начала 90-х годов показали, что бактерии и археи имеют гомологи актина и тубулина и что они лежат в основе эукариотических микротрубочек и микрофиламентов. [42] Хотя эволюционные взаимоотношения настолько далеки, что не очевидны только из сравнения последовательностей белков, сходство их трехмерных структур и схожие функции по поддержанию формы и полярности клеток дают убедительные доказательства того, что эукариотические и прокариотические цитоскелеты действительно гомологичны. [43] Три лаборатории независимо друг от друга обнаружили, что FtsZ, белок, уже известный как ключевой игрок в бактериальном цитокинезе, имеет «последовательность сигнатуры тубулина», присутствующую во всех α-, β- и γ-тубулинах. [42] Однако некоторые структуры бактериального цитоскелета, возможно, еще не идентифицированы. [28] [44]
FtsZ был первым идентифицированным белком цитоскелета прокариот. Подобно тубулину, FtsZ образует нити в присутствии гуанозинтрифосфата (GTP), но эти нити не группируются в канальцы. Во время деления клеток FtsZ является первым белком, который перемещается к месту деления, и необходим для привлечения других белков, которые синтезируют новую клеточную стенку между делящимися клетками.
Прокариотические актиноподобные белки, такие как MreB , участвуют в поддержании формы клеток. Все несферические бактерии имеют гены, кодирующие актиноподобные белки, и эти белки образуют спиральную сеть под клеточной мембраной, которая направляет белки, участвующие в биосинтезе клеточной стенки . [45]
Бактерия Caulobacter crescentus содержит третий белок — кресцентин , родственный промежуточным нитям эукариотических клеток. Кресцентин также участвует в поддержании формы клеток, таких как спиральные и вибриоидные формы бактерий, но механизм, с помощью которого он это делает, в настоящее время неясен. [47] Кроме того, искривление можно описать смещением серповидных нитей после нарушения синтеза пептидогликана. [48]
Цитоскелет представляет собой высокоанизотропную и динамическую сеть, постоянно ремоделирующуюся в ответ на изменение клеточного микроокружения. Сеть влияет на механику и динамику клетки путем дифференциальной полимеризации и деполимеризации составляющих ее филаментов (в первую очередь актина и миозина, но также играют роль микротрубочки и промежуточные филаменты). [49] Это генерирует силы, которые играют важную роль в информировании клетки о ее микроокружении. В частности, было показано, что такие силы, как напряжение, жесткость и силы сдвига, влияют на судьбу клеток, дифференцировку, миграцию и подвижность. [49] Посредством процесса, называемого «механотрансдукцией», клетка реконструирует свой цитоскелет, чтобы чувствовать эти силы и реагировать на них.
Механотрансдукция в значительной степени зависит от фокальных спаек , которые по существу соединяют внутриклеточный цитоскелет с внеклеточным матриксом (ECM). Посредством фокальных спаек клетка способна интегрировать внеклеточные силы во внутриклеточные, поскольку белки, присутствующие в фокальных спайках, претерпевают конформационные изменения, чтобы инициировать сигнальные каскады. Было показано, что такие белки, как киназа фокальной адгезии (FAK) и Src, передают силовые сигналы в ответ на клеточную активность, такую как пролиферация и дифференцировка, и предположительно являются ключевыми сенсорами на пути механотрансдукции. [50] В результате механотрансдукции цитоскелет меняет свой состав и/или ориентацию, чтобы приспособиться к силовому стимулу и обеспечить соответствующую реакцию клетки.
Цитоскелет меняет механику клетки в ответ на обнаруженные силы. Например, увеличение напряжения внутри плазматической мембраны повышает вероятность открытия ионных каналов, что увеличивает ионную проводимость и значительно повышает вероятность клеточных изменений притока или оттока ионов. [50] Более того, механические свойства клеток определяют, насколько далеко и в каком направлении сила будет распространяться по клетке и как она будет изменять ее динамику. [51] Например, мембранный белок, который не связан тесно с цитоскелетом, не будет оказывать существенного влияния на корковую актиновую сеть, если на него будет воздействовать специально направленная сила. Однако мембранные белки, более тесно связанные с цитоскелетом, будут вызывать более значительный ответ. [50] Таким образом, анизотропия цитоскелета служит для более четкого направления клеточных ответов на внутри- или внеклеточные сигналы.
Конкретные пути и механизмы, с помощью которых цитоскелет воспринимает силы и реагирует на них, все еще изучаются. Однако известно, что дальний порядок, генерируемый цитоскелетом, способствует механотрансдукции. [52] Клетки диаметром около 10–50 мкм в несколько тысяч раз больше молекул, находящихся в цитоплазме, которые необходимы для координации клеточной деятельности. Поскольку клетки настолько велики по сравнению с основными биомолекулами, из-за отсутствия организующей сети различным частям цитоплазмы трудно взаимодействовать. [53] Более того, биомолекулы должны полимеризоваться до длины, сравнимой с длиной клетки, но образующиеся полимеры могут быть сильно дезорганизованы и неспособны эффективно передавать сигналы из одной части цитоплазмы в другую. Таким образом, необходимо иметь цитоскелет для организации полимеров и обеспечения их эффективной связи по всей клетке.
По определению, цитоскелет состоит из белков, которые могут образовывать продольные массивы (волокна) у всех организмов. Эти белки, образующие нити, были разделены на 4 класса. Тубулиноподобные , актиноподобные , цитоскелетные АТФазы Уолкера А (WACA-белки) и промежуточные филаменты . [8] [28]
Тубулиноподобные белки — это тубулин у эукариот и FtsZ , TubZ, RepX у прокариот. Актиноподобными белками являются актин у эукариот и MreB , FtsA у прокариот. Примером WACA-белков, которые в основном встречаются у прокариот, является MinD . Примерами промежуточных филаментов, которые встречаются почти исключительно у животных (т.е. у эукариот), являются ламины , кератины , виментин , нейрофиламенты и десмин . [8]
Цитоскелетные белки обычно коррелируют с формой клеток, сегрегацией ДНК и делением клеток у прокариот и эукариот. Какие белки выполняют какую задачу – это очень разные вещи. Например, сегрегация ДНК у всех эукариот происходит за счет использования тубулина, но у прокариот могут использоваться либо белки WACA, либо актиноподобные, либо тубулиноподобные белки. Деление клеток опосредовано у эукариот актином, но у прокариотов обычно тубулиноподобными (часто FtsZ-кольцо) белками и иногда ( термопротеотами ) ESCRT-III , который у эукариот все еще играет роль на последней стадии деления. [8]
Цитоплазматический поток , также известный как циклоз, представляет собой активное движение содержимого клетки по компонентам цитоскелета. Хотя этот процесс в основном наблюдается у растений, все типы клеток используют этот процесс для транспортировки отходов, питательных веществ и органелл в другие части клетки. [54] Клетки растений и водорослей обычно крупнее многих других клеток; поэтому поток цитоплазмы важен для этих типов клеток. Это связано с тем, что дополнительный объем клетки требует потока цитоплазмы для перемещения органелл по всей клетке. [55] Органеллы движутся вдоль микрофиламентов в цитоскелете, управляемые миозиновыми моторами, связывающими и толкающими пучки актиновых филаментов. [54]
^ В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (8 июня 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 3.2. Цитоплазма и клеточные органеллы. ISBN 978-1-947172-04-3 .
^ МакКинли, Майкл; Дин О'Локлин, Валери; Пеннефатер-О'Брайен, Элизабет; Харрис, Рональд (2015). Анатомия человека (4-е изд.). Нью-Йорк: Образование Макгроу Хилл. п. 29. ISBN 978-0-07-352573-0 .
^ Гели М.И., Ризман Х. (апрель 1998 г.). «Эндоцитарная интернализация в дрожжевых и животных клетках: сходство и различие». Журнал клеточной науки . 111 (Пт 8) (8): 1031–7. дои : 10.1242/jcs.111.8.1031 . ПМИД 9512499 .
^ Петерс РА. «Лекции Харбена, 1929 г. Перепечатано в: Питерс, Р.А. (1963) Биохимические поражения и летальный синтез, стр. 216. Pergamon Press, Оксфорд». {{cite journal}}: Для цитирования журнала требуется |journal= ( помощь )
^ Полин Д., Ли З. (ноябрь 2004 г.). «Десмин: основной белок промежуточных филаментов, необходимый для структурной целостности и функции мышц». Экспериментальные исследования клеток . 301 (1): 1–7. doi : 10.1016/j.yexcr.2004.08.004 . ПМИД 15501438 .
Arc.Ask3.Ru Номер скриншота №: 25f9b1d1807ff43dc540084629708b8c__1716791640 URL1:https://arc.ask3.ru/arc/aa/25/8c/25f9b1d1807ff43dc540084629708b8c.html Заголовок, (Title) документа по адресу, URL1: Cytoskeleton - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)



 В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (8 июня 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 3.2. Цитоплазма и клеточные органеллы. ISBN 978-1-947172-04-3 .
В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (8 июня 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 3.2. Цитоплазма и клеточные органеллы. ISBN 978-1-947172-04-3 .