Ядрышко
| Клеточная биология | |
|---|---|
| Диаграмма клеток животных | |
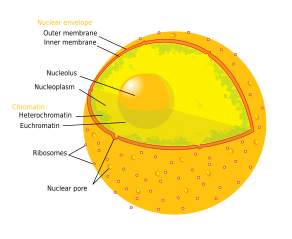
 Компоненты типичной животной клетки:
|
Ядрышко nj ( / nj uː ˈ k l iː ə l ə s , ˌ в uː k l i ˈ oʊ l ə s / ; мн .: ядрышки / -l aɪ / ) самая крупная структура ядре эукариотических клеток — . [1] Он наиболее известен как место биогенеза рибосом , то есть синтеза рибосом. Ядрышко также участвует в формировании частиц распознавания сигналов и играет роль в реакции клетки на стресс. [2] Ядрышки состоят из белков , ДНК и РНК и формируются вокруг определенных хромосомных областей, называемых ядрышковыми организующими областями . Нарушение функции ядрышек может быть причиной ряда заболеваний человека, называемых «нуклеопатиями». [3] [4] и ядрышко исследуется как мишень для рака химиотерапии . [5] [6]
История [ править ]
Ядрышко было идентифицировано с помощью светлопольной микроскопии в 1830-х годах. [7] Теодор Шванн в своем трактате 1939 года описывает, что Шлейден обнаружил в ядрах небольшие тельца, и называет эту структуру «Kernkörperchen». В переводе работы на английский язык в 1947 году структура получила название «ядрышко». [8] [9]
В дополнение к этим особенностям цитобласта, уже известным Брауном и Мейеном, Шлейден обнаружил внутри него небольшое тельце (см. табл. I, рис. 1, 4), которое в полностью развитом цитобласте выглядит как толстое кольцо или толстостенная полая глобула. Однако, по-видимому, он имеет различный вид в разных цитобластах. Иногда можно различить только внешний резко очерченный круг этого кольца с темной точкой в центре, а иногда, а чаще всего, лишь резко очерченное пятно. В других случаях это пятно очень маленькое, а иногда его вообще невозможно распознать. Поскольку об этом теле часто придется говорить в следующем трактате, я для краткости назову его «ядрышком» (Kernkorperchen, «ядро-тельце»).
- Теодор Шванн, перевод Генри Смита, «Микроскопические исследования соответствия структуры и роста животных и растений», стр. 3.
Мало что было известно о функции ядрышка до 1964 года, когда было проведено исследование. [10] Исследования ядрышек, проведенные Джоном Гердоном и Дональдом Брауном у африканской шпорцевой лягушки Xenopus laevis, вызвали растущий интерес к функции и детальной структуре ядрышка. Они обнаружили, что 25% яиц лягушек не имели ядрышка и что такие яйца нежизнеспособны. Половина яиц имела одно ядрышко, а 25% — два. Они пришли к выводу, что ядрышко выполняет функцию, необходимую для жизни. В 1966 году Макс Л. Бирнстил показали и его коллеги с помощью экспериментов по гибридизации нуклеиновых кислот , что ДНК в ядрышках кодирует рибосомальную РНК . [11] [12]
Структура [ править ]
Выделяют три основных компонента ядрышка: фибриллярный центр (FC), плотный фибриллярный компонент (DFC) и зернистый компонент (GC). [1] Транскрипция рДНК происходит в ФК. [13] DFC содержит белок фибрилларин . [13] что важно для процессинга рРНК. ГК содержит белок нуклеофосмин . [13] (В23 на внешнем изображении), который также участвует в биогенезе рибосом .
Однако было высказано предположение, что эта конкретная организация наблюдается только у высших эукариот и что она развилась из двудольной организации при переходе от анамниот к амниотам . Отражая существенное увеличение межгенной области ДНК , исходный фибриллярный компонент должен был разделиться на FC и DFC. [14]
Другая структура, обнаруженная во многих ядрышках (особенно у растений), представляет собой четкую область в центре структуры, называемую ядрышковой вакуолью. [15] Было показано, что ядрышки различных видов растений содержат очень высокие концентрации железа. [16] в отличие от ядрышек клеток человека и животных.
ядрышка Ультраструктуру можно увидеть с помощью электронного микроскопа , а организацию и динамику можно изучить с помощью флуоресцентного мечения белков и восстановления флуоресценции после фотообесцвечивания ( FRAP ). Антитела против белка PAF49 также можно использовать в качестве маркера ядрышка в экспериментах по иммунофлуоресценции. [17]
Хотя обычно можно увидеть только одно или два ядрышка, диплоидная человеческая клетка имеет десять областей организатора ядрышка (ЯОР) и может иметь больше ядрышек. Чаще всего в каждом ядрышке участвуют несколько ЯОР. [18]
Функция и сборка рибосом [ править ]

Для биогенеза рибосом необходимы две из трех эукариотических РНК-полимераз ( Pol I и Pol III ), и они функционируют скоординировано. На начальной стадии гены рРНК транскрибируются как единая единица внутри ядрышка с помощью РНК-полимеразы I. Для того чтобы произошла эта транскрипция, необходимы несколько факторов, связанных с pol I, и ДНК-специфичных транс-действующих факторов. У дрожжей наиболее важными являются: UAF ( вышестоящий активирующий фактор ), TBP (белок, связывающий ТАТА-бокс) и основной связывающий фактор (CBF), которые связывают элементы промотора и образуют преинициативный комплекс (PIC), который, в свою очередь, распознается с помощью РНК-полимеразы. У людей аналогичный PIC собирается с SL1 , фактором селективности промотора (состоящим из TBP и TBP-ассоциированных факторов , или TAF), факторами инициации транскрипции и UBF (восходящим фактором связывания). РНК-полимераза I транскрибирует большинство транскриптов рРНК (28S, 18S и 5,8S), но субъединица 5S рРНК (компонент рибосомальной субъединицы 60S) транскрибируется РНК-полимеразой III. [19]
Транскрипция рРНК приводит к образованию длинной молекулы-предшественника (45S пре-рРНК), которая все еще содержит ITS и ETS. Для создания молекул 18S РНК, 5,8S и 28S РНК необходим дальнейший процессинг. У эукариот ферменты, модифицирующие РНК, доставляются к соответствующим сайтам узнавания за счет взаимодействия с направляющими РНК, которые связывают эти специфические последовательности. Эти направляющие РНК относятся к классу малых ядрышковых РНК ( мякРНК ), которые находятся в комплексе с белками и существуют в виде малых ядрышково- рибонуклеопротеинов ( мякРНП ). После процессинга субъединиц рРНК они готовы к сборке в более крупные рибосомальные субъединицы. Однако необходима также дополнительная молекула рРНК, 5S рРНК. У дрожжей последовательность 5S рДНК локализована в межгенном спейсере и транскрибируется в ядрышке РНК-полимеразой.
У высших эукариот и растений ситуация более сложная, поскольку 5S-последовательность ДНК лежит вне ЯОР и транскрибируется РНК Пол III в нуклеоплазме , после чего попадает в ядрышко для участия в сборке рибосомы. В этой сборке участвуют не только рРНК, но и рибосомальные белки. Гены, кодирующие эти r-белки, транскрибируются Pol II в нуклеоплазме по «традиционному» пути синтеза белка (транскрипция, процессинг пре-мРНК, ядерный экспорт зрелой мРНК и трансляция на цитоплазматических рибосомах). Зрелые r-белки затем импортируются в ядро и, наконец, в ядрышко. Ассоциация и созревание рРНК и р-белков приводят к образованию 40S (малых) и 60S (больших) субъединиц полной рибосомы. Они экспортируются через комплексы ядерных пор в цитоплазму, где остаются свободными или связываются с эндоплазматической сетью , образуя шероховатый эндоплазматический ретикулум (RER). [20] [21]
В клетках эндометрия человека иногда образуется сеть ядрышковых каналов. Происхождение и функции этой сети до сих пор четко не определены. [22]
Секвестрация белков [ править ]
Помимо своей роли в биогенезе рибосом, ядрышко, как известно, захватывает и иммобилизует белки - процесс, известный как ядрышковое задержание. Белки, задержанные в ядрышке, не способны диффундировать и взаимодействовать со своими партнерами по связыванию. Мишенью этого механизма посттрансляционной регуляции являются VHL , PML , MDM2 , POLD1 , RelA , HAND1 и hTERT , среди многих других. Сейчас известно, что за это явление ответственны длинные некодирующие РНК, происходящие из межгенных участков ядрышка. [23]
См. также [ править ]
Ссылки [ править ]
- ^ Перейти обратно: а б О'Салливан Дж. М., Пай Д. А., Cridge AG, Энгельке Д. Р., Гэнли А. Р. (июнь 2013 г.). «Ядрышко: плот, дрейфующий в ядерном море, или краеугольный камень ядерной структуры?» . Биомолекулярные концепции . 4 (3): 277–86. дои : 10.1515/bmc-2012-0043 . ПМК 5100006 . ПМИД 25436580 .
- ^ Олсон М.О., Дундр М. (16 февраля 2015 г.). «Ядрышко: строение и функция». Энциклопедия наук о жизни (eLS) . дои : 10.1002/9780470015902.a0005975.pub3 . ISBN 978-0-470-01617-6 .
- ^ Гетман М (июнь 2014 г.). «Роль ядрышка в заболеваниях человека. Предисловие» . Биохимика и биофизика Acta . 1842 (6): 757. doi : 10.1016/j.bbadis.2014.03.004 . ПМИД 24631655 .
- ^ Бахадори, М; Азизи, Миннесота; Дабири, С; Бахадори, Н. (2022). «Влияние ядрышка человека на вирусную жизнь гостей, с акцентом на инфекцию COVID-19: мини-обзор» . Иранский журнал патологии . 17 (1): 1–7. дои : 10.30699/IJP.2021.540305.2744 . ПМЦ 8794558 . ПМИД 35096082 .
- ^ Куин Дж.Э., Девлин Дж.Р., Кэмерон Д., Ханнан К.М., Пирсон Р.Б., Ханнан Р.Д. (июнь 2014 г.). «Нацеливание на ядрышко при раковом вмешательстве» . Biochimica et Biophysica Acta (BBA) - Молекулярные основы болезней . 1842 (6): 802–16. дои : 10.1016/j.bbadis.2013.12.009 . hdl : 11343/44176 . ПМИД 24389329 .
- ^ Вудс С.Дж., Ханнан К.М., Пирсон Р.Б., Ханнан Р.Д. (июль 2015 г.). «Ядрышко как фундаментальный регулятор реакции р53 и новая мишень для терапии рака». Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1849 (7): 821–9. дои : 10.1016/j.bbagrm.2014.10.007 . ПМИД 25464032 .
- ^ Педерсон Т. (март 2011 г.). «Ядрышко» . Перспективы Колд-Спринг-Харбор в биологии . 3 (3): а000638. doi : 10.1101/cshperspect.a000638 . ПМК 3039934 . ПМИД 21106648 .
- ^ «Микроскопические исследования согласия в строении и росте животных и растений / Т. Шванн. С четырьмя медными пластинками» . Приветственная коллекция . Проверено 12 марта 2024 г.
- ^ Шванн, Теодор; Шванн, Теодор; Смит, Генри; Шлейден, MJ (1847). Микроскопические исследования соответствия строения и роста животных и растений . Лондон: Сиденхэмское общество. п. 3.
- ^ Браун Д.Д., Гердон Дж.Б. (январь 1964 г.). «Отсутствие синтеза рибосомальной РНК у безъядерного мутанта xenopus laevis» . Труды Национальной академии наук Соединенных Штатов Америки . 51 (1): 139–46. Бибкод : 1964PNAS...51..139B . дои : 10.1073/pnas.51.1.139 . ПМК 300879 . ПМИД 14106673 .
- ^ Бирнстил М.Л., Уоллес Х., Сирлин Дж.Л., Фишберг М. (декабрь 1966 г.). «Локализация комплементов рибосомальной ДНК в области ядрышкового организатора Xenopus laevis». Монография Национального института рака . 23 : 431–47. ПМИД 5963987 .
- ^ Уоллес Х., Бирнстил М.Л. (февраль 1966 г.). «Рибосомальные цистроны и ядрышковый организатор». Biochimica et Biophysical Acta (BBA) - Нуклеиновые кислоты и синтез белка . 114 (2): 296–310. дои : 10.1016/0005-2787(66)90311-x . ПМИД 5943882 .
- ^ Перейти обратно: а б с Сирри В., Уркуки-Инчима С., Руссель П., Эрнандес-Верден Д. (январь 2008 г.). «Ядрышко: удивительное ядерное тело» . Гистохимия и клеточная биология . 129 (1): 13–31. дои : 10.1007/s00418-007-0359-6 . ПМК 2137947 . ПМИД 18046571 .
- ^ Тири М., Лафонтен Д.Л. (апрель 2005 г.). «Рождение ядрышка: эволюция ядрышковых компартментов». Тенденции в клеточной биологии . 15 (4): 194–9. дои : 10.1016/j.tcb.2005.02.007 . ПМИД 15817375 . в формате PDF. Архивировано 17 декабря 2008 г. в Wayback Machine.
- ^ Бевен А.Ф., Ли Р., Разаз М., Лидер-ди-джей, Браун Дж.В., Шоу П.Дж. (июнь 1996 г.). «Организация процессинга рибосомальной РНК коррелирует с распределением ядрышковых мяРНК» . Журнал клеточной науки . 109 (Часть 6) (6): 1241–51. дои : 10.1242/jcs.109.6.1241 . ПМИД 8799814 .
- ^ Рошцтардц Х., Грийе Л., Изаур М.П., Конехеро Г., Ортега Р., Кюри К., Мари С. (август 2011 г.). «Ядрышко растительной клетки как горячая точка для железа» . Журнал биологической химии . 286 (32): 27863–6. дои : 10.1074/jbc.C111.269720 . ПМК 3151030 . ПМИД 21719700 .
- ^ Антитело PAF49 | ДжинТекс Инк . Genetex.com. Проверено 18 июля 2019 г.
- ^ фон Кнебель Дебериц М., Вентценсен Н. (2008). «Клетка: основная структура и функция». Комплексная цитопатология (третье изд.).
- ^ Чамп ПК, Харви Р.А., Ферье Д.Р. (2005). Иллюстрированные обзоры Липпинкотта: биохимия . Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-2265-0 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. стр. 331–3. ISBN 978-0-8153-3218-3 .
- ^ Купер ГМ, Хаусман Р.Э. (2007). Клетка: молекулярный подход (4-е изд.). Синауэр Ассошиэйтс. стр. 371–9. ISBN 978-0-87893-220-7 .
- ^ Ван Т., Шнайдер Дж. (1 июля 1992 г.). «Происхождение и судьба системы ядрышковых каналов нормального эндометрия человека» . Клеточные исследования . 2 (2): 97–102. дои : 10.1038/cr.1992.10 .
- ^ Аудас Т.Э., Джейкоб, доктор медицинских наук, Ли С. (январь 2012 г.). «Иммобилизация белков в ядрышке рибосомальным межгенным спейсером, некодирующим РНК» . Молекулярная клетка . 45 (2): 147–57. doi : 10.1016/j.molcel.2011.12.012 . ПМИД 22284675 .
Дальнейшее чтение [ править ]
- Купер GM (2000). «Ядрышко» . Клетка: молекулярный подход (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-106-4 .
- Тику В., Антеби А. (август 2018 г.). «Функция ядрышка в регуляции продолжительности жизни». Тенденции в клеточной биологии . 28 (8): 662–672. дои : 10.1016/j.tcb.2018.03.007 . ПМИД 29779866 . S2CID 29167518 .
- ДжоАнна Кляйн (20 мая 2018 г.). «Вещь внутри ваших клеток, которая может определить, как долго вы проживете» . Нью-Йорк Таймс .
Внешние ссылки [ править ]
- Ядрышко под электронным микроскопом II на uni-mainz.de
- База данных ядерных белков – поиск по отсеку
- Клетка + ядрышко в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Гистологическое изображение: 20104loa - Система обучения гистологии в Бостонском университете.
| Базы данных органов управления : Национальные |
|---|