Конденсин

Конденсины — это крупные белковые комплексы , которые играют центральную роль в хромосом сборке и сегрегации во время митоза и мейоза (рис. 1). [ 1 ] [ 2 ] Их субъединицы первоначально были идентифицированы как основные компоненты митотических хромосом, собранных в Xenopus экстрактах яиц . [ 3 ]
Субъединичный состав
[ редактировать ]Эукариотические типы
[ редактировать ]
Многие эукариотические клетки обладают двумя разными типами конденсиновых комплексов, известными как конденсин I и конденсин II , каждый из которых состоит из пяти субъединиц (рис. 2). [ 4 ] Конденсины I и II имеют одну и ту же пару основных субъединиц, SMC2 и SMC4, которые принадлежат к большому семейству хромосомных АТФаз , известных как белки SMC (SMC означает «структурное поддержание хромосом»). [ 5 ] [ 6 ] Каждый из комплексов содержит отдельный набор регуляторных субъединиц, не относящихся к SMC ( клейзина ). субъединица [ 7 ] и пара субъединиц повтора HEAT ). [ 8 ] Оба комплекса крупные, имеют общую молекулярную массу 650-700 кДа.
| Сложный | Субъединица | Классификация | Позвоночные животные | Д. меланогастер | К. Элеганс | С. cerevisiae | С. насос | А. Талиана | С. меролаэ | Т. термофила |
|---|---|---|---|---|---|---|---|---|---|---|
| конденсин I и II | СМК2 | SMC-АТФаза | CAP-E/SMC2 | СМК2 | МИКС-1 | смк2 | Вырезать14 | CAP-E1&-E2 | СМК2 | Скм2 |
| конденсин I и II | SMC4 | SMC-АТФаза | CAP-C/SMC4 | SMC4/Глюон | СМК-4 | смк4 | Вырезать3 | КАП-С | SMC4 | смк4 |
| конденсин I | КАП-Д2 | ТЕПЛО -IA | КАП-Д2 | КАП-Д2 | ДПЯ-28 | Ycs4 | Кнд1 | CAB72176 | КАП-Д2 | Комп.1 и 2 |
| конденсин I | КАП-Г | ТЕПЛО -IB | КАП-Г | КАП-Г | КАП-G1 | Ycg1 | Кнд3 | БАБ08309 | КАП-Г | Cpg1 |
| конденсин I | CAP-H | клейзин | CAP-H | CAP-H/Бесплодный | ДПИ-26 | Брн1 | Кнд2 | ААС25941 | CAP-H | Cph1,2,3,4&5 |
| конденсин II | КАП-Д3 | ТЕПЛО - ТАМ | КАП-Д3 | КАП-Д3 | HCP-6 | - | - | At4g15890.1 | КАП-Д3 | - |
| конденсин II | КАП-G2 | ТЕПЛО -IIB | КАП-G2 | - | КАП-G2 | - | - | CAP-G2/HEB1 | КАП-G2 | - |
| конденсин II | КАП-H2 | клейзин | КАП-H2 | КАП-H2 | КЛЮЧ-2 | - | - | CAP-H2/HEB2 | КАП-H2 | - |
| конденсин I округ Колумбия | вариант SMC4 | SMC-АТФаза | - | - | ДПЯ-27 | - | - | - | - | - |
Конденсины основных субъединиц (SMC2 и SMC4) консервативны среди всех видов эукариот , изученных на сегодняшний день. Не-SMC-субъединицы, уникальные для конденсина I, также консервативны среди эукариот, но встречаемость не-SMC-субъединиц, уникальных для конденсина II, сильно варьирует среди видов.
- Например, у плодовой мушки Drosophila melanogaster нет гена субъединицы CAP-G2 конденсина II. [ 9 ] У других видов насекомых также часто отсутствуют гены субъединиц CAP-D3 и/или CAP-H, что указывает на то, что субъединицы, отличные от SMC, уникальные для конденсина II, находились под высоким давлением отбора во время эволюции насекомых. [ 10 ]
- Нематода . Caenorhabditis elegans обладает как конденсинами I, так и II Этот вид, однако, уникален в том смысле, что у него есть третий комплекс (тесно связанный с конденсином I), который участвует в регуляции генов во всей хромосоме , т. е. дозовой компенсации . [ 11 ] В этом комплексе, известном как конденсин I округ Колумбия аутентичная субъединица SMC4 заменяется ее вариантом DPY-27 (рис. 2).
- У некоторых видов, таких как грибы (например, почкующиеся дрожжи Saccharomyces cerevisiae и делящиеся дрожжи Schizosaccharomyces pombe ), отсутствуют все регуляторные субъединицы, уникальные для конденсина II. [ 12 ] [ 13 ] С другой стороны, одноклеточная примитивная красная водоросль Cyanidioschyzon merolae , размер генома которой сравним с размером генома дрожжей, имеет как конденсины I, так и II. [ 14 ] Таким образом, не существует очевидной связи между наличием конденсина II и размером геномов эукариот.
- Инфузория имеет только Tetrahymena thermophila конденсин I. Тем не менее, существует множество паралогов для двух его регуляторных субъединиц (CAP-D2 и CAP-H), и некоторые из них специфически локализуются либо в макронуклеусе (отвечающем за экспрессию генов), либо в микроядре (отвечающем за размножение). [ 15 ] Таким образом, этот вид имеет множественные комплексы конденсина I, которые имеют разные регуляторные субъединицы и демонстрируют четкую ядерную локализацию. [ 16 ] Это очень уникальное свойство, которого нет у других видов.
Прокариотические типы
[ редактировать ]У прокариотических видов также есть конденсиноподобные комплексы, которые играют важную роль в организации и сегрегации хромосом ( нуклеоидов ). Прокариотические конденсины можно разделить на два типа: SMC-ScpAB. [ 17 ] и МукБЭФ . [ 18 ] Многие виды эубактерий и архей имеют SMC-ScpAB, тогда как подгруппа эубактерий (известных как Gammaproteobacteria ), включая Escherichia coli, имеет MukBEF. ScpA и MukF принадлежат к семейству белков, называемых «клезинами». [ 7 ] тогда как ScpB и MukE недавно были отнесены к новому семейству белков, названному «воздушный змей». [ 19 ]
| Сложный | Субъединица | Классификация | Б. субтилис | Каулобактер | кишечная палочка |
|---|---|---|---|---|---|
| SMC-ScpAB | СМК | АТФаза | СМК/БсСМК | СМК | - |
| SMC-ScpAB | ScpA | клейзин | ScpA | ScpA | - |
| SMC-ScpAB | ScpB | видеть | ScpB | ScpB | - |
| МукБЭФ | МукБ | АТФаза | - | - | МукБ |
| МукБЭФ | МукЭ | видеть | - | - | МукЭ |
| МукБЭФ | МукФ | клейзин | - | - | МукФ |
Несмотря на сильно расходящиеся первичные структуры соответствующих субъединиц между SMC-ScpAB и MukBEF, разумно предположить, что эти два комплекса играют схожие, если не идентичные функции в организации и динамике прокариотических хромосом, основываясь на их молекулярной архитектуре и их дефектных клеточных фенотипах. Поэтому оба комплекса часто называют прокариотическими (или бактериальными) конденсинами. Недавние исследования сообщают о возникновении третьего комплекса, связанного с MukBEF (названного MksBEF), у некоторых видов бактерий. [ 20 ]
Молекулярные механизмы
[ редактировать ]Молекулярные структуры
[ редактировать ]
Димеры SMC, которые действуют как основные субъединицы конденсинов, имеют весьма характерную V-образную форму, каждое плечо которой состоит из антипараллельных спиральных спиралей (рис. 3; см. в белках SMC ). подробности [ 21 ] [ 22 ] Длина каждого спирального плеча достигает ~50 нм, что соответствует длине ~150 п.н. двухцепочечной ДНК (дцДНК). В эукариотических комплексах конденсина I и II субъединица клейзина соединяет два головных домена димера SMC и связывается с двумя субъединицами повтора HEAT (рис. 1). [ 23 ] [ 24 ]
Ранние исследования выяснили структуру частей бактериальных конденсинов, таких как MukBEF. [ 25 ] [ 26 ] и SMC-ScpA. [ 27 ] [ 28 ] Сообщалось о нескольких структурах подкомплексов и субдоменов эукариотических комплексов, включая шарнирный и плечевой домены димера SMC2-SMC4, [ 29 ] [ 30 ] субкомплекс CAP-G(ycg1)/CAP-H(brn1), [ 31 ] [ 32 ] и подкомплекс CAP-D2(ycs4)/CAP-H(brn1). [ 24 ] Недавнее крио-ЭМ исследование показало, что конденсин претерпевает большие конформационные изменения, которые связаны со связыванием АТФ и гидролизом его субъединиц SMC. [ 33 ] С другой стороны, высокоскоростная атомно-силовая микроскопия продемонстрировала, что плечи димера SMC гораздо более гибкие, чем ожидалось. [ 34 ]
Молекулярная деятельность
[ редактировать ]Конденсин I, очищенный из Xenopus яиц , представляет собой ДНК -стимулируемую АТФазу и демонстрирует способность придавать положительное сверхспиральное натяжение дцДНК экстрактов активность АТФ -гидролиз-зависимым способом (положительная суперспирализации ). [ 35 ] [ 36 ] Аналогичная активность была обнаружена в конденсинах других организмов. [ 37 ] [ 38 ] Положительная активность суперспирализации активируется in vitro посредством Cdk1 фосфорилирования , что указывает на то, что это, вероятно, одна из физиологических активностей, непосредственно участвующих в сборке митотических хромосом. [ 39 ] Предполагается, что эта активность конденсина I помогает сворачивать ДНК и способствует опосредованному топоизомеразой II разрешению сестринских хроматид. [ 40 ] Ранние эксперименты с одиночными молекулами ДНК также продемонстрировали в реальном времени, что конденсин I способен уплотнять ДНК зависимым от АТФ-гидролиза образом. [ 41 ]
Совсем недавно эксперименты на одиночных молекулах продемонстрировали, что конденсин I почкующихся дрожжей способен перемещаться по дцДНК ( двигательная активность). [ 42 ] и «экструдировать» петли ДНК ( деятельность по экструзии петель ) [ 43 ] в зависимости от гидролиза АТФ. В последних экспериментах активность отдельных комплексов конденсина на ДНК была визуализирована с помощью флуоресцентной визуализации в реальном времени , что показало, что конденсин I действительно является двигателем быстрой экструзии петель и что одиночный комплекс конденсина I может экструдировать 1500 п.о. ДНК в секунду. строго АТФ-зависимым образом. Было высказано предположение, что конденсин I закрепляет ДНК между субъединицами Ycg1-Brn1. [ 31 ] и асимметрично тянет ДНК, образуя большие петли. Более того, было показано, что конденсиновые комплексы могут пересекать друг друга, образуя динамические петлевые структуры и изменяя свои размеры. [ 44 ]
Неизвестно, как конденсины могут действовать на нуклеосомную ДНК . Недавняя разработка системы восстановления выявила, что гистоновый шаперон FACT является важным компонентом конденсина I-опосредованной сборки хромосом in vitro , что дает важный ключ к разгадке этой проблемы. [ 45 ] Также было показано, что конденсины могут собирать хромосомоподобные структуры в бесклеточных экстрактах даже в условиях, когда сборка нуклеосом в значительной степени подавлена. [ 46 ] Это наблюдение указывает на то, что конденсины могут воздействовать, по крайней мере частично, на ненуклеосомную ДНК в физиологических условиях.
Насколько схожи и насколько различны молекулярные активности конденсина I и конденсина II? Оба имеют две общие субъединицы SMC, но каждая имеет три уникальные субъединицы, не относящиеся к SMC (рис. 2). Точный баланс между действиями этих субъединиц, не относящихся к SMC, может определять различия в скорости экструзии петель. [ 47 ] и активность сборки митотических хромосом [ 48 ] [ 49 ] [ 50 ] [ 51 ] из двух комплексов. Вводя различные мутации, можно превратить конденсин I в комплекс с активностью, подобной конденсину II, и наоборот. [ 51 ]
Математическое моделирование
[ редактировать ]нескольких попытках математического и компьютерного моделирования Сообщалось о сборки митотических хромосом на основе молекулярной активности конденсинов. К показательным относятся моделирование на основе выдавливания петель, [ 52 ] стохастические парные контакты [ 53 ] и сочетание циклических и межконденсинных притяжений. [ 54 ]
Функции сборки и разделения хромосом
[ редактировать ]Митоз
[ редактировать ]
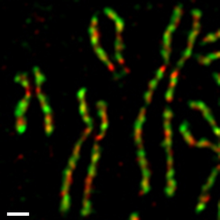
В клетках культуры тканей человека два конденсиновых комплекса регулируются по-разному во время митотического клеточного цикла (рис. 4). [ 55 ] [ 56 ] Конденсин II присутствует в ядре клетки во время интерфазы и участвует в ранней стадии конденсации хромосом в профазном ядре. С другой стороны, конденсин I присутствует в цитоплазме во время интерфазы и получает доступ к хромосомам только после разрушения ядерной оболочки (NEBD) в конце профазы. Во время прометафазы и метафазы конденсин I и конденсин II взаимодействуют, собирая палочковидные хромосомы, в которых две сестринские хроматиды полностью разделяются. Такая дифференциальная динамика двух комплексов наблюдается в Xenopus экстрактах яиц . [ 57 ] мышиные ооциты, [ 58 ] и нервные стволовые клетки, [ 59 ] что указывает на то, что это часть фундаментального регуляторного механизма, консервативного среди разных организмов и типов клеток. Скорее всего, этот механизм обеспечивает упорядоченное действие двух комплексов: сначала конденсина II, а затем конденсина I. [ 60 ]
На метафазных хромосомах конденсины I и II обогащены центральной осью неперекрывающимся образом (рис. 5). Эксперименты по истощению in vivo [ 4 ] [ 59 ] [ 61 ] и эксперименты по истощению иммунитета с Xenopus экстрактами яиц [ 57 ] демонстрируют, что эти два комплекса имеют разные функции при сборке метафазных хромосом. Клетки с дефицитом функций конденсина не задерживаются на определенной стадии клеточного цикла, демонстрируя дефекты сегрегации хромосом (т.е. анафазные мосты) и прогрессируя посредством аномального цитокинеза. [ 62 ] [ 63 ]
Относительный вклад конденсинов I и II в митоз варьируется у разных видов эукариот. Например, каждый из конденсинов I и II играет важную роль в эмбриональном развитии мышей. [ 59 ] Они выполняют как перекрывающиеся, так и неперекрывающиеся функции во время митотического клеточного цикла. С другой стороны, конденсин II не является необходимым для митоза у примитивной водоросли C. merolae. [ 14 ] и наземное растение A. thaliana . [ 64 ] Любопытно, что конденсин II играет доминантную роль над конденсином I у ранних эмбрионов C. elegans . [ 11 ] Эта особенность может быть связана с тем, что C. elegans имеет специализированную структуру хромосом, известную как голоцентрические хромосомы . Грибы, такие как S. cerevisiae. [ 13 ] и С. помбе [ 12 ] не имеют конденсина II из первого. Эти различия между видами эукариот имеют важное значение для эволюции архитектуры хромосом (см. раздел «Эволюционные последствия» ниже).
| разновидность | М. мышца | Д. меланогастер | К. Элеганс | С. cerevisiae | С. насос | А. Талиана | С. меролаэ |
|---|---|---|---|---|---|---|---|
| размер генома | ~2500 МБ | 140 Мб | 100 Мб | 12 Мб | 14 Мб | 125 Мб | 16 Мб |
| конденсин I | существенный | существенный | незначительный | существенный | существенный | существенный | существенный |
| конденсин II | существенный | несущественный | существенный | - | - | несущественный | несущественный |
Недавно стало возможным, что структурные изменения хромосом, зависящие от клеточного цикла, можно отслеживать с помощью основанного на геномике метода, известного как Hi-C (высокопроизводительный захват конформации хромосом ). [ 65 ] Влияние дефицита конденсина на конформацию хромосом изучалось у почкующихся дрожжей. [ 66 ] [ 67 ] делящиеся дрожжи, [ 68 ] [ 69 ] и куриные клетки DT40. [ 70 ] Результаты этих исследований убедительно подтверждают представление о том, что конденсины играют решающую роль в сборке митотических хромосом и что конденсины I и II выполняют разные функции в этом процессе. Более того, количественный анализ изображений позволяет исследователям подсчитать количество конденсиновых комплексов, присутствующих на метафазных хромосомах человека. [ 71 ]
Мейоз
[ редактировать ]Конденсины также играют важную роль в сборке и сегрегации хромосом при мейозе . Сообщалось о генетических исследованиях S. cerevisiae , [ 72 ] Д. меланогастер , [ 73 ] [ 74 ] и C. elegans . [ 75 ] На мышах потребность в субъединицах конденсина в мейозе была решена с помощью экспериментов по блокированию, опосредованному антителами. [ 58 ] и анализ условного нокаута генов . [ 76 ] При мейозе I млекопитающих функциональный вклад конденсина II оказывается больше, чем вклад конденсина I. Как было показано при митозе, [ 59 ] однако два конденсиновых комплекса также имеют как перекрывающиеся, так и неперекрывающиеся функции в мейозе. В отличие от когезина , до сих пор не идентифицировано мейоз-специфичных субъединиц конденсинов.
Хромосомные функции вне митоза или мейоза
[ редактировать ]Недавние исследования показали, что конденсины участвуют в широком спектре функций хромосом, помимо митоза или мейоза . [ 60 ]
- У почкующихся дрожжей конденсин I (единственный конденсин в этом организме) участвует в регуляции числа копий рДНК. повтора [ 77 ] а также в кластеризации генов тРНК . [ 78 ]
- У делящихся дрожжей конденсин I участвует в регуляции репликативной контрольной точки. [ 79 ] и кластеризация генов, транскрибируемых РНК-полимеразой III. [ 80 ]
- У C. elegans третий конденсиновый комплекс (конденсин I округ Колумбия ), связанный с конденсином I, регулирует структуру Х-хромосом высшего порядка как основной регулятор дозовой компенсации . [ 81 ]
- У D. melanogaster субъединицы конденсина II способствуют растворению политенных хромосом. [ 82 ] и образование хромосомных территорий [ 83 ] в питающих клетках яичников . Имеются доказательства того, что они отрицательно регулируют трансвекцию в диплоидных клетках. Также сообщалось, что компоненты конденсина I необходимы для обеспечения правильной экспрессии генов в нейронах после выхода из клеточного цикла. [ 84 ]
- У A. thaliana конденсин II необходим для толерантности к избыточному борному стрессу, возможно, за счет облегчения повреждения ДНК. [ 64 ]
- В клетках млекопитающих вполне вероятно, что конденсин II участвует в регуляции архитектуры и функции интерфазных хромосом. Например, в клетках человека конденсин II участвует в инициации разрешения сестринских хроматид во время S-фазы, задолго до митотической профазы, когда сестринские хроматиды становятся цитологически видимыми. [ 85 ]
- В интерфазных ядрах мыши прицентромерный гетерохроматин на разных хромосомах связывается друг с другом, образуя большую структуру, известную как хромоцентры. Клетки с дефицитом конденсина II, но не конденсина I, демонстрируют гиперкластеризацию хромцентров, что указывает на то, что конденсин II играет специфическую роль в подавлении кластеризации хромоцентров. [ 59 ]
- Хотя ранние исследования предполагали возможность того, что конденсины могут напрямую участвовать в регуляции экспрессии генов , некоторые недавние исследования опровергают эту гипотезу. [ 86 ] [ 87 ]
- Были получены мутанты делящихся дрожжей Schizosaccharomyces pombe , которые имели фенотип, чувствительный к температуре и/или повреждению ДНК . [ 88 ] Некоторые из этих мутантов были дефектными в субъединицах HEAT конденсина, что указывает на то, что субъединицы HEAT необходимы для репарации ДНК . [ 88 ]
Посттрансляционные модификации и регуляция клеточного цикла
[ редактировать ]Субъединицы конденсина подвергаются различным посттрансляционным модификациям , зависящим от клеточного цикла . [ 89 ] Среди них фосфорилирование при митозе. лучше всего изучено [ 90 ]
Фосфорилирование с помощью Cdk1 необходимо для сверхспиральной активности конденсина I. [ 39 ] [ 38 ] и активность сборки хромосом [ 45 ] in vitro. Однако целевые субъединицы и сайты (и количество) фосфорилирования, необходимые для активации, неизвестны. Последовательности S/TP, основные мишени Cdk1, имеют тенденцию быть обогащенными внутренне неупорядоченными областями (IDR), расположенными на концах субъединиц конденсина. [ 90 ] но их распределение и вклад в регуляцию конденсина широко варьируются у разных видов. Например, у делящихся дрожжей фосфорилирование N-конца субъединицы SMC4 регулирует ядерную транслокацию конденсина во время митоза. [ 12 ] У почкующихся дрожжей конденсин локализуется в ядре на протяжении всего клеточного цикла, а фосфорилирование N-конца субъединицы SMC4 участвует в регуляции динамики хромосомных ассоциаций конденсина. [ 91 ] [ 92 ] Было высказано предположение, что у позвоночных животных N-концевое фосфорилирование субъединицы CAP-H способствует специфической для митоза загрузке конденсина I. [ 93 ] Помимо Cdk1, положительная регуляция со стороны Aurora B [ 94 ] [ 95 ] и поло [ 38 ] и негативная регуляция CK2 (казеинкиназа 2) [ 96 ] было сообщено.
Несколько митотических киназ Cdk1 , [ 97 ] [ 98 ] [ 50 ] [ 51 ] курица [ 99 ] и Мпс1 [ 100 ] участвуют в регуляции конденсина II. Было показано, что С-концевой хвост субъединицы CAP-D3 является основной мишенью фосфорилирования Cdk1 в комплексе конденсина II человека. [ 51 ] Более того, CAP-D3 был идентифицирован как субстрат протеинфосфатазы PP2A -B55. [ 101 ]
Сообщалось, что субъединица CAP-H2 конденсина II разрушается у дрозофилы под действием убиквитинлигазы SCFSlimb . [ 102 ]
Связь с болезнями
[ редактировать ]Было продемонстрировано, что MCPH1, один из белков, ответственных за первичную микроцефалию человека , обладает способностью отрицательно регулировать конденсин II. [ 103 ] В клетках пациента mcph1 конденсин II (но не конденсин I) гиперактивируется, что приводит к преждевременной конденсации хромосом в фазе G2 (т.е. перед вступлением в митоз). [ 104 ] Однако нет доказательств того, что неправильная регуляция конденсина II напрямую связана с этиологией микроцефалии mcph1 . Совсем недавно сообщалось, что гипоморфные мутации в субъединицах конденсина I или II вызывают микроцефалию у людей. [ 105 ] У мышей гипоморфные мутации в субъединицах конденсина II вызывают специфические дефекты развития Т-клеток . [ 106 ] приводит к Т-клеточной лимфоме . [ 107 ] Интересно отметить, что типы клеток со специализированными способами клеточного деления, такие как нейральные стволовые клетки и Т-клетки , особенно восприимчивы к мутациям в субъединицах конденсина.
Эволюционные последствия
[ редактировать ]Прокариоты имеют примитивные типы конденсинов, [ 17 ] [ 18 ] что указывает на то, что эволюционное происхождение конденсинов предшествует происхождению гистонов. Тот факт, что конденсины I и II широко консервативны среди современных видов эукариот, убедительно указывает на то, что последний общий предок эукариот ( LECA ) имел оба комплекса. [ 60 ] Поэтому разумно предположить, что некоторые виды, например грибы, утратили конденсин II в ходе эволюции .
Тогда почему у многих эукариот есть два разных конденсиновых комплекса? Как обсуждалось выше, относительный вклад конденсинов I и II в митоз варьируется у разных организмов. Они играют одинаково важную роль в митозе млекопитающих, тогда как у многих других видов конденсин I играет преобладающую роль по сравнению с конденсином II. У этих видов конденсин II мог быть адаптирован для выполнения различных второстепенных функций, кроме митоза . [ 64 ] [ 82 ] Хотя очевидной связи между наличием конденсина II и размером генома нет, кажется, что функциональный вклад конденсина II становится большим по мере увеличения размера генома. [ 14 ] [ 59 ] Недавнее всестороннее исследование Hi-C утверждает с эволюционной точки зрения, что конденсин II действует как детерминант, который преобразует постмитотические конфигурации Рабля в интерфазные хромосомные территории. [ 108 ] Относительный вклад двух конденсиновых комплексов в архитектуру митотических хромосом также меняется во время развития, оказывая влияние на морфологию митотических хромосом. [ 57 ] Таким образом, балансирование конденсинов I и II, по-видимому, тонко настроено как в эволюции, так и в развитии.
Родственники
[ редактировать ]Эукариотические клетки имеют два дополнительных класса белковых комплексов SMC . Когезин содержит SMC1 и SMC3 и участвует в слипании сестринских хроматид. Комплекс SMC5/6 содержит SMC5 и SMC6 и участвует в рекомбинационной репарации.
См. также
[ редактировать ]- хромосома
- нуклеоид
- митоз
- мейоз
- клеточный цикл
- когезин
- SMC-белок
- АТФаза
- ТЕПЛО повтор
- Топоизомераза II
- Суперспираль ДНК
Ссылки
[ редактировать ]- ^ Хирано Т (2016). «Организация хромосом на основе конденсина от бактерий до позвоночных» . Клетка . 164 (5): 847–857. дои : 10.1016/j.cell.2016.01.033 . ПМИД 26919425 .
- ^ Калицыс П., Чжан Т., Маршалл К.М., Нильсен К.Ф., Хадсон Д.Ф. (2017). «Конденсин, главный организатор генома». Хромосома Res . 25 (1): 61–76. дои : 10.1007/s10577-017-9553-0 . ПМИД 28181049 . S2CID 28241964 .
- ^ Хирано Т., Кобаяши Р., Хирано М. (1997). «Конденсины, комплекс конденсации хромосом, содержащий XCAP-C, XCAP-E и гомолог Xenopus белка Drosophila Barren» . Клетка . 89 (4): 511–21. дои : 10.1016/S0092-8674(00)80233-0 . ПМИД 9160743 . S2CID 15061740 .
- ^ Jump up to: а б Оно Т., Лосада А., Хирано М., Майерс М.П., Нойвальд А.Ф., Хирано Т. (2003). «Дифференциальный вклад конденсина I и конденсина II в архитектуру митотических хромосом в клетках позвоночных» . Клетка . 115 (1): 109–21. дои : 10.1016/s0092-8674(03)00724-4 . ПМИД 14532007 . S2CID 18811084 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ульманн Ф (2016). «Комплексы SMC: от ДНК к хромосомам». Нат. Преподобный мол. Клеточная Биол . 17 (7): 399–412. дои : 10.1038/номер.2016.30 . ПМИД 27075410 . S2CID 20398243 .
- ^ Яцкевич С., Роудс Дж., Нэсмит К. (2019). «Организация хромосомной ДНК комплексами ГМК» . Анну. Преподобный Жене . 53 : 445–482. doi : 10.1146/annurev-genet-112618-043633 . ПМИД 31577909 .
- ^ Jump up to: а б Шлейффер А., Кейтна С., Маурер-Штро С., Глотцер М., Нэсмит К., Эйзенхабер Ф. (2003). «Клейзины: суперсемейство бактериальных и эукариотических белков-партнеров SMC» . Мол. Клетка . 11 (3): 571–5. дои : 10.1016/S1097-2765(03)00108-4 . ПМИД 12667442 .
- ^ Нойвальд А.Ф., Хирано Т. (2000). «Повторы HEAT, связанные с конденсинами, когезинами и другими комплексами, участвующими в функциях, связанных с хромосомами» . Геном Рез . 10 (10): 1445–52. дои : 10.1101/гр.147400 . ПМК 310966 . ПМИД 11042144 .
- ^ Херцог С., Нагаркар Джайсвал С., Урбан Э., Ример А., Фишер С., Хайдманн С.К. (2013). «Функциональное вскрытие субъединицы конденсина Cap-G Drosophila melanogaster выявило ее исключительную связь с конденсином I» . ПЛОС Генет . 9 (4): e1003463. дои : 10.1371/journal.pgen.1003463 . ПМК 3630105 . ПМИД 23637630 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кинг, Томас Д.; Леонард, Кристофер Дж; Купер, Джейкоб С; Нгуен, Сон; Джойс, Эрик Ф; Фаднис, Нитин; Такахаси, Ая (октябрь 2019 г.). «Периодические потери и быстрая эволюция комплекса конденсина II у насекомых» . Молекулярная биология и эволюция . 36 (10): 2195–2204. дои : 10.1093/molbev/msz140 . ПМК 6759200 . ПМИД 31270536 .
- ^ Jump up to: а б Чанковски Г, Коллетт К, Спал К, Кэри Дж, Снайдер М, Петти Е, Патель Ю, Табучи Т, Лю Х, Маклеод И, Томпсон Дж, Саркешик А, Йейтс Дж, Мейер Б.Дж., Хагстром К. (2009). «Три различных комплекса конденсина контролируют динамику хромосом C. elegans» . Курс. Биол . 19 (1): 9–19. дои : 10.1016/j.cub.2008.12.006 . ПМЦ 2682549 . ПМИД 19119011 .
- ^ Jump up to: а б с Сутани Т., Юаса Т., Томонага Т., Домае Н., Такио К., Янагида М. (1999). «Конденсиновый комплекс делящихся дрожжей: важная роль субъединиц, не относящихся к SMC, для конденсации и фосфорилирования Cdc2 Cut3 / SMC4» . Генс Дев . 13 (17): 2271–83. дои : 10.1101/gad.13.17.2271 . ПМК 316991 . ПМИД 10485849 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Фриман Л., Арагон-Алкайд Л., Струнников А. (2000). «Комплекс конденсина управляет конденсацией хромосом и митотической передачей рДНК» . Дж. Клеточная Биол . 149 (4): 811–824. дои : 10.1083/jcb.149.4.811 . ПМК 2174567 . ПМИД 10811823 .
- ^ Jump up to: а б с Фудзивара Т., Танака К., Куроива Т., Хирано Т. (2013). «Пространственно-временная динамика конденсинов I и II: эволюционные идеи примитивной красной водоросли Cyanidioschyzon merolae» . Мол. Биол. Клетка . 24 (16): 2515–27. дои : 10.1091/mbc.E13-04-0208 . ПМЦ 3744952 . ПМИД 23783031 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ховард-Тилль Р., Лойдл Дж. (2018). «Конденсины способствуют индивидуализации и сегрегации хромосом во время митоза, мейоза и амитоза у Tetrahymena thermophila» . Мол. Биол. Клетка . 29 (4): 466–478. doi : 10.1091/mbc.E17-07-0451 . ПМК 6014175 . ПМИД 29237819 .
- ^ Ховард-Тилль, Рэйчел; Тянь, Мяо; Лойдл, Йозеф; Коэн-Фикс, Орна (15 мая 2019 г.). «Специализированный конденсиновый комплекс участвует в созревании соматического ядра» . Молекулярная биология клетки . 30 (11): 1326–1338. doi : 10.1091/mbc.E18-08-0487 . ПМК 6724606 . ПМИД 30893010 .
- ^ Jump up to: а б Маскаренхас Дж., Соппа Дж., Струнников А.В., Грауманн П.Л. (2002). «Зависимая от клеточного цикла локализация двух новых прокариотических белков сегрегации и конденсации хромосом в Bacillus subtilis, которые взаимодействуют с белком SMC» . ЭМБО Дж . 21 (12): 3108–18. дои : 10.1093/emboj/cdf314 . ПМК 126067 . ПМИД 12065423 .
- ^ Jump up to: а б Ямадзоэ М., Оноги Т., Сунако Ю., Ники Х., Яманака К., Ичимура Т., Хирага С. (1999). «Комплексное образование белков MukB, MukE и MukF, участвующих в разделении хромосом у Escherichia coli» . ЭМБО Дж. 18 (21): 5873–8 дои : 10.1093/emboj/18.21.5873 . ПМЦ 1171653 . ПМИД 10545099 .
- ^ Палечек Дж. Дж., Грубер С. (2015). «Белки воздушного змея: суперсемейство партнеров SMC/клейзина, консервативное у бактерий, архей и эукариот» . Структура . 23 (12): 2183–2190. дои : 10.1016/j.str.2015.10.004 . ПМИД 26585514 .
- ^ Петрушенко З.М., Ше Ш, Рыбенков В.В. (2011). «Новое семейство бактериальных конденсинов» . Мол. Микробиол . 81 (4): 881–896. дои : 10.1111/j.1365-2958.2011.07763.x . ПМК 3179180 . ПМИД 21752107 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мелби Т.Э., Чампальо К.Н., Бриско Дж., Эриксон Х.П. (1998). «Симметричная структура структурного обеспечения хромосом (SMC) и белков MukB: длинные антипараллельные спиральные спирали, сложенные гибким шарниром» . Дж. Клеточная Биол . 142 (6): 1595–1604. дои : 10.1083/jcb.142.6.1595 . ПМК 2141774 . ПМИД 9744887 .
- ^ Андерсон Д.Е., Лосада А., Эриксон Х.П., Хирано Т. (2002). «Конденсин и когезин демонстрируют различную конформацию плеч с характерными шарнирными углами» . Дж. Клеточная Биол . 156 (6): 419–424. дои : 10.1083/jcb.200111002 . ПМК 2173330 . ПМИД 11815634 .
- ^ Онн И, Аоно Н, Хирано М, Хирано Т (2007). «Восстановление и геометрия субъединиц конденсиновых комплексов человека» . ЭМБО Дж . 26 (4): 1024–1034. дои : 10.1038/sj.emboj.7601562 . ПМЦ 1852836 . ПМИД 17268547 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Хасслер М., Шалтиэль И.А., Кшонсак М., Саймон Б., Меркель Ф., Терихен Л., Бэйли Х.Дж., Макошек Дж., Браво С., Мец Дж., Хенниг Дж., Херинг Х.Х. (2019). «Структурные основы асимметричного конденсинового АТФазного цикла» . Мол Клетка . 74 (6): 1175–1188.e24. doi : 10.1016/j.molcel.2019.03.037 . ПМК 6591010 . ПМИД 31226277 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Феннелл-Феззи Р., Градиа С.Д., Эйки Д., Бергер Дж.М. (2005). «Субъединица MukF конденсина Escherichia coli: архитектура и функциональное отношение к клейзинам» . ЭМБО Дж . 24 (11): 1921–1930. дои : 10.1038/sj.emboj.7600680 . ПМЦ 1142612 . ПМИД 15902272 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ву Дж.С., Лим Дж.Х., Шин Х.К., Су МК, Ку Б, Ли К.Х., Джу К., Робинсон Х., Ли Дж., Пак С.И., Ха NC, О Б.Х. (2009). «Структурные исследования бактериального конденсинового комплекса выявили АТФ-зависимое нарушение межсубъединичных взаимодействий» . Клетка . 136 (1): 85–96. дои : 10.1016/j.cell.2008.10.050 . ПМИД 19135891 . S2CID 4608756 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бюрманн Ф., Шин Х.К., Баскен Дж., Со Ю.М., Хименес-Оя В., Ким Ю.Г., О Б.Х., Грубер С. (2013). «Асимметричный мостик SMC-клейзин в прокариотическом конденсине». Нат. Структура. Мол. Биол . 20 (3): 371–379. дои : 10.1038/nsmb.2488 . ПМИД 23353789 . S2CID 21584205 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Камада К., Мията М., Хирано Т. (2013). «Молекулярные основы активации SMC-АТФазы: роль внутренних структурных изменений регуляторного субкомплекса ScpAB» . Структура . 21 (4): 581–594. дои : 10.1016/j.str.2013.02.016 . ПМИД 23541893 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Гриз Дж. Дж., Витте Г., Хопфнер К. П. (2010). «Структура и ДНК-связывающая активность шарнирного домена конденсина мыши подчеркивают общие и разнообразные особенности белков SMC» . Нуклеиновые кислоты Рез . 38 (10): 3454–3465. дои : 10.1093/nar/gkq038 . ПМЦ 2879519 . ПМИД 20139420 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Со Й, Бюрман Ф, Шин Х, Ода Т, Джин КС, Тоузленд КП, Ким С, Ли Х, Ким СДж, Конг М, Дюран-Дибольд М, Ким Й, Ким ХМ, Ли НК, Сато М, О Б, Грубер С (2015). «Молекулярные основы образования палочек ГМК и их растворения при связывании ДНК» . Мол. Клетка . 57 (2): 290–303. дои : 10.1016/j.molcel.2014.11.023 . ПМК 4306524 . ПМИД 25557547 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Кшонсак М., Меркель Ф., Бишт С., Мец Дж., Рыбин В., Хасслер М., Херинг CH (2017). «Структурная основа механизма ремня безопасности, который прикрепляет конденсин к хромосомам» . Клетка . 171 (3): 588–600.e24. дои : 10.1016/j.cell.2017.09.008 . ПМЦ 5651216 . ПМИД 28988770 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хара, Кодай; Киносита, Мигита, Томоко; Симидзу, Кенитиро, Кодо; Хасимото, Хироши (12 марта 2019 г.) . EMBO . Отчеты embr.201847183 20 5). doi : 10.15252/ . PMC 6501013. ( PMID 30858338 .
- ^ Ли Б.Г., Меркель Ф., Аллегретти М., Хасслер М., Кавуд С., Лекомт Л., О'Рейли Ф.Дж., Синн Л.Р., Гутьеррес-Эскрибано П., Кшонсак М., Браво С., Накане Т., Раппсильбер Дж., Арагон Л., Бек М., Лёве Дж, Херинг CH (2020). «Крио-ЭМ структуры голоконденсина обнаруживают механизм субъединичного триггера» . Nat Struct Мол Биол . 27 (8): 743–751. дои : 10.1038/s41594-020-0457-x . ПМЦ 7610691 . ПМИД 32661420 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ифтенс Дж.М., Катан А.Дж., Кшонсак М., Хасслер М., де Вильде Л., Диф Э.М., Херинг Ч.Х., Деккер С. (2016). «Димеры конденсина Smc2-Smc4 гибки и динамичны» . Представитель ячейки . 14 (8): 1813–1818. дои : 10.1016/j.celrep.2016.01.063 . ПМЦ 4785793 . ПМИД 26904946 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кимура К., Хирано Т. (1997). «АТФ-зависимая положительная суперспирализация ДНК с помощью 13S-конденсина: биохимическое значение конденсации хромосом» . Клетка . 90 (4): 625–634. дои : 10.1016/s0092-8674(00)80524-3 . ПМИД 9288743 . S2CID 15876604 .
- ^ Кимура К., Рыбенков В.В., Крисона Н.Дж., Хирано Т., Коццарелли Н.Р. (1999). «Конденсин 13S активно реконфигурирует ДНК, вызывая глобальное положительное скручивание: последствия для конденсации хромосом» . Клетка . 98 (2): 239–248. дои : 10.1016/s0092-8674(00)81018-1 . ПМИД 10428035 . S2CID 16671030 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хагстром К.А., Холмс В.Ф., Коцарелли Н.Р., Мейер Б.Дж. (2002). «Конденсин C. elegans способствует архитектуре митотических хромосом, организации центромер и сегрегации сестринских хроматид во время митоза и мейоза» . Генс Дев . 16 (6): 729–742. дои : 10.1101/gad.968302 . ПМК 155363 . ПМИД 11914278 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б с Сен-Пьер Ж, Дузьек М, Базиль Ф, Паскариу М, Бонней Э, Сове В, Рацима Х, Д'Амур Д (2009). «Полокиназа регулирует конденсацию митотических хромосом путем гиперактивации активности суперспирализации конденсиновой ДНК» . Мол Клетка . 120 (Часть 7): 1245–1255. doi : 10.1016/j.molcel.2009.04.013 . ПМИД 19481522 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Кимура К., Хирано М., Кобаяши Р., Хирано Т. (1998). «Фосфорилирование и активация 13S-конденсина с помощью Cdc2 in vitro». Наука . 282 (5388): 487–490. Бибкод : 1998Sci...282..487K . дои : 10.1126/science.282.5388.487 . ПМИД 9774278 .
- ^ Бакстер Дж., Сен Н., Мартинес В.Л., Де Карандини М.Э., Шварцман Дж.Б., Диффли Дж.Ф., Арагон Л. (2011). «Положительная суперспирализация митотической ДНК вызывает декатенацию топоизомеразы II у эукариот». Наука . 331 (6022): 1328–1332. Бибкод : 2011Sci...331.1328B . дои : 10.1126/science.1201538 . ПМИД 21393545 . S2CID 34081946 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Стрик Т.Р., Кавагути Т., Хирано Т. (2004). «Обнаружение в реальном времени уплотнения одиночных молекул ДНК конденсином I» . Курс. Биол . 14 (10): 874–880. дои : 10.1016/j.cub.2004.04.038 . ПМИД 15186743 . S2CID 10078994 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Теракава Т., Бишт С., Ифтенс Дж. М., Деккер С., Хэринг Ч., Грин ЕС (2017). «Комплекс конденсина представляет собой механохимический двигатель, который перемещается по ДНК» . Наука . 358 (6363): 672–676. Бибкод : 2017Sci...358..672T . дои : 10.1126/science.aan6516 . ПМК 5862036 . ПМИД 28882993 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ганджи М., Шалтиэль И.А., Бишт С., Ким Э., Каличава А., Херинг Ч.Х., Деккер С. (2018). «Визуализация экструзии петель ДНК конденсином в реальном времени» . Наука . 360 (6384): 102–105. Бибкод : 2018Sci...360..102G . дои : 10.1126/science.aar7831 . ПМК 6329450 . ПМИД 29472443 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ким Э., Керссемакерс Дж., Шалтиэль И.А., Херинг Ч.Х., Деккер С. (2020). «Конденсиновые комплексы, экструдирующие петли ДНК, могут пересекать друг друга» . Природа . 579 (7799): 438–442. Бибкод : 2020Natur.579..438K . дои : 10.1038/s41586-020-2067-5 . ПМИД 32132705 . S2CID 212407150 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Синтоми К., Такахаши Т.С., Хирано Т. (2015). «Восстановление митотических хроматид минимальным набором очищенных факторов». Nat Cell Biol . 17 (8): 1014–1023. дои : 10.1038/ncb3187 . ПМИД 26075356 . S2CID 8332012 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Синтоми К., Иноуэ Ф., Ватанабэ Х., Осуми К., Осуги М., Хирано Т. (2017). «Митотическая сборка хромосом, несмотря на истощение нуклеосом в Xenopus экстрактах яиц » . Наука . 356 (6344): 1284–1287. Бибкод : 2017Sci...356.1284S . дои : 10.1126/science.aam9702 . ПМИД 28522692 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Конг М., Каттс Э.Э., Пан Д., Берон Ф., Калияппан Т., Сюэ С., Моррис Э.П., Мусаккио А., Ваннини А., Грин ЕС (2020). «Человеческие конденсины I и II вызывают обширное АТФ-зависимое уплотнение ДНК, связанной с нуклеосомами» . Мол. Клетка . 79 (1): 99–114. doi : 10.1016/j.molcel.2020.04.026 . ПМЦ 7335352 . ПМИД 32445620 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Киносита К., Кобаяши Т.Дж., Хирано Т. (2015). «Балансирующие действия двух HEAT-субъединиц конденсина I поддерживают динамическую сборку осей хромосом» . Ячейка разработчиков . 33 (1): 94–106. дои : 10.1016/j.devcel.2015.01.034 . ПМИД 25850674 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Киносита К., Цубота Ю., Тане С., Айзава Ю., Саката Р., Такеучи К., Синтоми К., Нисияма Т., Хирано Т. (2022). «Механизм, независимый от экструзии петли, способствует формированию хромосом, опосредованному конденсином I» . J Клеточная Биол . 221 (3): e202109016. дои : 10.1083/jcb.202109016 . ПМЦ 8932526 . ПМИД 35045152 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Ёсида М.М., Киносита К., Айзава Ю., Тане С., Ямашита Д., Синтоми К., Хирано Т. (2022). «Молекулярное рассечение сборки хромосом, опосредованной конденсином II, с использованием анализов in vitro» . электронная жизнь . 11 : е78984. doi : 10.7554/eLife.78984 . ПМЦ 9433093 . ПМИД 35983835 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б с д Ёсида М.М., Киносита К., Синтоми К., Айзава Ю., Хирано Т. (2023). «Регуляция конденсина II посредством механизмов самоподавления и высвобождения» . Мол Биол Клетка . 35 (2): mbcE23100392. дои : 10.1091/mbc.E23-10-0392 . ПМЦ 10881152 . ПМИД 38088875 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Голобородько Антон; Имакаев Максим В.; Марко, Джон Ф; Мирный, Леонид (18 мая 2016 г.). «Уплотнение и сегрегация сестринских хроматид посредством активной экструзии петли» . электронная жизнь . 5 . дои : 10.7554/eLife.14864 . ПМЦ 4914367 . ПМИД 27192037 .
- ^ Ченг, Тэмми М.К.; Хигер, Себастьян; Шалей, Рафаэль АГ; Мэтьюз, Ник; Стюарт, Энгус; Райт, Джон; Лим, Кармей; Бейтс, Пол А; Ульманн, Франк (29 апреля 2015 г.). «Простая биофизическая модель имитирует конденсацию хромосом почкующихся дрожжей» . электронная жизнь . 4 : e05565. doi : 10.7554/eLife.05565 . ПМЦ 4413874 . ПМИД 25922992 .
- ^ Сакаи, Юджи; Мотидзуки, Ацуши; Киносита, Казухиса; Хирано, Тацуя; Татикава, Масаси; Морозов, Александр В. (18 июня 2018 г.). «Моделирование функций конденсина в формировании и сегрегации хромосом» . PLOS Вычислительная биология . 14 (6): e1006152. Бибкод : 2018PLSCB..14E6152S . дои : 10.1371/journal.pcbi.1006152 . ПМК 6005465 . ПМИД 29912867 .
- ^ Оно Т., Фанг Ю., Спектор Д.Л., Хирано Т. (2004). «Пространственная и временная регуляция конденсинов I и II в сборке митотических хромосом в клетках человека» . Мол. Биол. Клетка . 15 (7): 3296–308. дои : 10.1091/mbc.E04-03-0242 . ПМЦ 452584 . ПМИД 15146063 .
- ^ Хирота Т., Герлих Д., Кох Б., Элленберг Дж., Петерс Дж.М. (2004). «Различные функции конденсина I и II в сборке митотических хромосом» . Дж. Клеточная наука . 117 (Часть 26): 6435–45. дои : 10.1242/jcs.01604 . ПМИД 15572404 .
- ^ Jump up to: а б с Синтоми К., Хирано Т. (2011). «Относительное соотношение конденсина I и II определяет форму хромосом» . Генс Дев . 25 (14): 1464–1469. дои : 10.1101/gad.2060311 . ПМК 3143936 . ПМИД 21715560 .
- ^ Jump up to: а б Ли Дж., Огуши С., Сайто М., Хирано Т. (2011). «Конденсины I и II необходимы для построения двухвалентных хромосом в ооцитах мыши» . Мол. Биол. Клетка . 22 (18): 3465–3477. doi : 10.1091/mbc.E11-05-0423 . ПМК 3172270 . ПМИД 21795393 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б с д и ж Нишиде К., Хирано Т. (2014). «Перекрывающиеся и непересекающиеся функции конденсинов I и II в делениях нервных стволовых клеток» . ПЛОС Генет . 10 (12): e1004847. дои : 10.1371/journal.pgen.1004847 . ПМК 4256295 . ПМИД 25474630 .
- ^ Jump up to: а б с Хирано Т (2012). «Конденсины: универсальные организаторы хромосом с разнообразными функциями» . Генс Дев . 26 (4): 1659–1678. дои : 10.1101/gad.194746.112 . ПМЦ 3418584 . ПМИД 22855829 .
- ^ Грин Л.С., Калитсис П., Чанг Т.М., Ципетик М., Ким Дж.Х., Маршалл О., Тернбулл Л., Уитчерч С.Б., Вагнарелли П., Самедзима К., Эрншоу В.К., Чу К.Х., Хадсон Д.Ф. (2012). «Контрастные роли конденсина I и конденсина II в формировании митотических хромосом» . Дж. Клеточная наука . 125 (Часть 6): 1591–1604. дои : 10.1242/jcs.097790 . ПМЦ 3336382 . ПМИД 22344259 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сака Ю., Сутани Т., Ямашита Ю., Сайто С., Такеучи М., Накасеко Ю., Янагида М. (1994). «Делящиеся дрожжи Cut3 и Cut14, члены повсеместно распространенного семейства белков, необходимы для конденсации и разделения хромосом в митозе» . ЭМБО Дж . 13 (20): 4938–4952. дои : 10.1002/j.1460-2075.1994.tb06821.x . ПМЦ 395434 . ПМИД 7957061 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хадсон Д.Ф., Вагнарелли П., Гассманн Р., Эрншоу В.К. (2003). «Конденсин необходим для сборки негистоновых белков и структурной целостности митотических хромосом позвоночных». Дев. Клетка . 5 (2): 323–336. дои : 10.1016/s1534-5807(03)00199-0 . ПМИД 12919682 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б с Сакамото Т., Инуи Ю.Т., Урагучи С., Ёсидзуми Т., Мацунага С., Мастуи М., Умеда М., Фукуи К., Фудзивара Т. (2011). «Конденсин II облегчает повреждение ДНК и необходим для устойчивости к стрессу, вызванному перегрузкой бора, у арабидопсиса» . Растительная клетка . 23 (9): 3533–3546. дои : 10.1105/tpc.111.086314 . ПМК 3203421 . ПМИД 21917552 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Наумова Н, Имакаев М, Фуденберг Г, Жан Ю, Лажуа Б.Р., Мирный Л.А., Деккер Дж (2013). «Организация митотической хромосомы» . Наука . 342 (6161): 948–953. Бибкод : 2013Sci...342..948N . дои : 10.1126/science.1236083 . ПМК 4040465 . ПМИД 24200812 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Шалбеттер С.А., Голобородько А, Фуденберг Г, Белтон Дж.М., Майлз С., Ю.М., Деккер Дж., Мирный Л., Бакстер Дж. (2017). «Комплексы SMC дифференциально компактируют митотические хромосомы в соответствии с геномным контекстом» . Nat Cell Biol . 19 (9): 1071–1080. дои : 10.1038/ncb3594 . ПМК 5640152 . ПМИД 28825700 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Лазар-Стефанита Л., Сколари В.Ф., Мерси Г., Мюллер Х., Герен Т.М., Тьерри А., Моцциконаччи Дж., Кошуль Р. (2017). «Когезины и конденсины управляют четырехмерной динамикой хромосом дрожжей во время клеточного цикла» . ЭМБО Дж. 36 (18): 2684–2697. дои : 10.15252/embj.201797342 . ПМЦ 5599795 . ПМИД 28729434 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Какуи Ю., Рабиновиц А., Барри DJ, Ульманн Ф. (2017). «Конденсин-опосредованное ремоделирование митотического ландшафта хроматина у делящихся дрожжей» . Нат Жене . 49 (10): 1553–1557. дои : 10.1038/ng.3938 . ПМК 5621628 . ПМИД 28825727 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Танидзава Х., Ким К.Д., Ивасаки О., Нома КИ (2017). «Архитектурные изменения генома делящихся дрожжей во время клеточного цикла» . Nat Struct Мол Биол . 24 (11): 965–976. дои : 10.1038/nsmb.3482 . ПМК 5724045 . ПМИД 28991264 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Гибкус, Йохан Х.; Самедзима, Кумико; Голобородько Антон; Самедзима, Итару; Наумова, Наталья; Нюблер, Йоханнес; Канемаки, Масато Т.; Се, Линьфэн; Полсон, Джеймс Р.; Эрншоу, Уильям К.; Мирный, Леонид А.; Деккер, Иов (9 февраля 2018 г.). «Путь образования митотических хромосом» . Наука . 359 (6376): eaao6135. дои : 10.1126/science.aao6135 . ПМЦ 5924687 . ПМИД 29348367 .
- ^ Вальтер, Найк; Хоссейн, М. Юлиус; Полити, Антонио З.; Кох, Биргит; Кюбельбек, Мориц; Эдегард-Фогнер, Эйвинд; Лампе, Марко; Элленберг, январь (2 июля 2018 г.). «Количественная карта конденсинов человека дает новое представление об архитектуре митотических хромосом» . Журнал клеточной биологии . 217 (7): 2309–2328. дои : 10.1083/jcb.201801048 . ПМК 6028534 . ПМИД 29632028 .
- ^ Ю Х.Г., Кошланд Д.Э. (2003). «Мейотический конденсин необходим для правильного уплотнения хромосом, сборки SC и разрешения зависимых от рекомбинации хромосомных связей» . Дж. Клеточная Биол . 163 (5): 937–947. дои : 10.1083/jcb.200308027 . ПМК 2173617 . ПМИД 14662740 .
- ^ Хартл Т.А., Суини С.Дж., Неплер П.Дж., Боско Дж. (2008). «Конденсин II разрешает хромосомные ассоциации, обеспечивая сегрегацию анафазы I в мейозе самцов дрозофилы» . ПЛОС Генет . 4 (10): e1000228. дои : 10.1371/journal.pgen.1000228 . ПМК 2562520 . ПМИД 18927632 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Резник Т.Д., Деж К.Дж., Сян Ю., Хоули Р.С., Ан С., Орр-Уивер Т.Л. (2009). «Мутации в хромосомном пассажирском комплексе и конденсинном комплексе по-разному влияют на разборку синаптонемного комплекса и конфигурацию метафазы I в мейозе самок дрозофилы» . Генетика . 181 (3): 875–887. doi : 10.1534/genetics.108.097741 . ПМК 2651061 . ПМИД 19104074 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Чан Р.К., Северсон А.Ф., Мейер Б.Дж. (2004). «Конденсин реструктурирует хромосомы при подготовке к мейотическим делениям» . Дж. Клеточная Биол . 167 (4): 613–625. дои : 10.1083/jcb.200408061 . ПМК 2172564 . ПМИД 15557118 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хоулард М., Годвин Дж., Метсон Дж., Ли Дж., Хирано Т., Нэсмит К. (2015). «Конденсин придает хромосомам продольную жесткость» . Nat Cell Biol . 17 (6): 771–81. дои : 10.1038/ncb3167 . ПМК 5207317 . ПМИД 25961503 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Джозука К., Терасава М., Огава Х., Огава Т., Хориучи Т. (2006). «Конденсин, загруженный на участок барьера репликационной вилки в гене рРНК, повторяется во время S-фазы FOB1-зависимым образом, чтобы предотвратить сокращение длинного повторяющегося массива у Saccharomyces cerevisiae» . Мол Клеточная Биол . 26 (6): 2226–2236. дои : 10.1128/MCB.26.6.2226-2236.2006 . ПМЦ 1430289 . ПМИД 16507999 .
- ^ Хеуслер Р.А., Пратт-Хаятт М., Гуд П.Д., Гипсон Т.А., Энгельке Д.Р. (2008). «Кластеризация генов тРНК дрожжей опосредуется специфической ассоциацией конденсина с комплексами транскрипции генов тРНК» . Генс Дев . 22 (16): 2204–2214. дои : 10.1101/gad.1675908 . ПМК 2518813 . ПМИД 18708579 .
- ^ Аоно Н., Сутани Т., Томонага Т., Мочида С., Янагида М. (2002). «Cnd2 играет двойную роль в митотической конденсации и интерфазе». Природа . 417 (6885): 197–202. Бибкод : 2002Natur.417..197A . дои : 10.1038/417197a . ПМИД 12000964 . S2CID 4332524 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ивасаки О, Танака А, Танизава Х, Гревал С.И., Нома К (2010). «Центромерная локализация рассеянных генов Pol III у делящихся дрожжей» . Мол. Биол. Ячейка 21 (2): 254–265. дои : 10.1091/mbc.e09-09-0790 . ПМЦ 2808234 . ПМИД 19910488 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Крейн Э., Биан К., МакКорд Р.П., Ладжуа Б.Р., Уиллер Б.С., Ралстон Э.Дж., Узава С., Деккер Дж., Мейер Б.Дж. (2015). «Ремоделирование топологии Х-хромосомы, вызванное конденсином, во время дозовой компенсации» . Природа . 523 (7559): 210–244. Бибкод : 2015Natur.523..240C . дои : 10.1038/nature14450 . ПМЦ 4498965 . ПМИД 26030525 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Хартл Т.А., Смит Х.Ф., Боско Дж. (2008). «Выравниванию и трансвекции хромосом противодействует конденсин II». Наука . 322 (5906): 1384–1387. Бибкод : 2008Sci...322.1384H . дои : 10.1126/science.1164216 . ПМИД 19039137 . S2CID 5154197 .
- ^ Бауэр Ч.Р., Хартл Т.А., Боско Г. (2012). «Конденсин II способствует образованию хромосомных территорий, вызывая аксиальное уплотнение полиплоидных интерфазных хромосом» . ПЛОС Генет . 8 (8): e1002873. дои : 10.1371/journal.pgen.1002873 . ПМЦ 3431300 . ПМИД 22956908 .
- ^ Хасан А., Арагуас Родригес П., Хайдманн С.К., Уолмсли Э.Л., Оги Дж.Н., Саутхолл ТД (2020). «Субъединица конденсина I Cap-G необходима для правильной экспрессии генов во время созревания постмитотических нейронов» . электронная жизнь . 9 : е55159. дои : 10.7554/eLife.55159 . ПМК 7170655 . PMID 32255428 .
- ^ Оно Т., Ямашита Д., Хирано Т. (2013). «Конденсин II инициирует разрешение сестринских хроматид во время S-фазы» . Дж. Клеточная Биол . 200 (4): 429–441. дои : 10.1083/jcb.201208008 . ПМЦ 3575537 . ПМИД 23401001 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Пол М.Р., Марковиц Т.Е., Хохваген А., Эркан С. (2018). «Истощение конденсина вызывает разуплотнение генома, не изменяя уровень глобальной экспрессии генов у Saccharomyces cerevisiae» . Генетика . 210 (1): 331–344. дои : 10.1534/genetics.118.301217 . ПМК 6116964 . ПМИД 29970489 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хоке С, Робелле Х, Модоло Л, Сан ХМ, Берни С, Кайлен-Херинг С, Тоселли Е, Клаудер-Мюнстер С, Штайнмец Л, Херинг Ч., Маргерат С., Бернар П. (2018). «Конденсин контролирует уровни клеточной РНК посредством точного разделения хромосом вместо того, чтобы напрямую регулировать транскрипцию» . электронная жизнь . 7 : е38517. дои : 10.7554/eLife.38517 . ПМК 6173581 . ПМИД 30230473 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Сюй X, Наказава Н., Янагида М. Субъединицы конденсина HEAT, необходимые для восстановления ДНК, функции кинетохор / центромеры и поддержания плоидности у делящихся дрожжей. ПЛОС Один. 12 марта 2015 г.; 10(3):e0119347. doi: 10.1371/journal.pone.0119347. ПМИД 25764183; PMCID: PMC4357468
- ^ Деккер Б., Деккер Дж. (2022). «Регуляция машин сворачивания митотических хромосом» . Биохим Дж . 479 (20): 2153–2173. дои : 10.1042/BCJ20210140 . ПМК 9704520 . ПМИД 36268993 .
- ^ Jump up to: а б Базиль Ф, Сен-Пьер Ж, Д'Амур Д (2010). «Трехэтапная модель активации конденсина во время конденсации митотических хромосом» . Клеточный цикл . 9 (16): 3243–3255. дои : 10.4161/cc.9.16.12620 . ПМИД 20703077 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Робеллет X, Таттикота Й, Ван Ф, Ви ТЛ, Паскариу М, Шанкар С, Боннейл Э, Браун СМ, Д'Амур Д (2015). «Высокочувствительный фосфопереключатель, запускаемый Cdk1, управляет морфогенезом хромосом во время деления клеток» . Генс Дев . 29 (4): 426–439. дои : 10.1101/gad.253294.114 . ПМЦ 4335297 . ПМИД 25691469 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тадани Р., Каменц Дж., Хигер С., Муньос С., Ульманн Ф. (2018). «Регуляция клеточного цикла динамической хромосомной ассоциации конденсинового комплекса» . Представитель ячейки . 23 (8): 2308–2317. дои : 10.1016/j.celrep.2018.04.082 . ПМЦ 5986713 . ПМИД 29791843 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тане С., Синтоми К., Киносита К., Цубота Ю., Ёсида М.М., Нисияма Т., Хирано Т. (2022). «Специфическая для клеточного цикла загрузка конденсина I регулируется N-концевым хвостом его субъединицы клейзина» . электронная жизнь . 11 : е84694. doi : 10.7554/eLife.84694 . ПМЦ 9797191 . ПМИД 36511239 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Липп Дж.Дж., Хирота Т., Позер I, Питерс Дж.М. (2007). «Аврора B контролирует ассоциацию конденсина I, но не конденсина II, с митотическими хромосомами». J Cell Sci . 120 (Часть 7): 1245–1255. дои : 10.1242/jcs.03425 . ПМИД 17356064 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Наказава Н., Мехротра Р., Эбе М., Янагида М. (2011). «Конденсин, фосфорилированный Aurora-B-подобной киназой Ark1, постоянно необходим до телофазы в режиме, отличном от Top2». J Cell Sci . 124 (Часть 11): 1795–1807. дои : 10.1242/jcs.078733 . ПМИД 21540296 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Такемото А., Кимура К., Янагисава Дж., Ёкояма С., Ханаока Ф. (2006). «Негативная регуляция конденсина I посредством CK2-опосредованного фосфорилирования» . ЭМБО Дж . 25 (22): 5339–5348. дои : 10.1038/sj.emboj.7601394 . ПМЦ 1636611 . ПМИД 17066080 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Абэ С., Нагасака К., Хираяма Ю., Кодзука-Хата Х., Ояма М., Аояги Ю., Обусе С., Хирота Т. (2011). «Начальная фаза конденсации хромосом требует Cdk1-опосредованного фосфорилирования субъединицы CAP-D3 конденсина II» . Джинс Дев 25 (8): 863–874. дои : 10.1101/gad.2016411 . ПМК 3078710 . ПМИД 21498573 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бахребах М., Чжан Т., Манн-младший, Калитсис П., Хадсон Д.Ф. (2015). «Нарушение консервативного треонина CAP-D3 изменяет нагрузку конденсина на митотических хромосомах, что приводит к гиперконденсации хромосом» . J Биол Хим . 290 (10): 6156–6167. дои : 10.1074/jbc.M114.627109 . ПМЦ 4358255 . ПМИД 25605712 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ким Дж.Х., Шим Дж., Джи М.Дж., Юнг Й., Бонг СМ, Чан Й.Дж., Юн ЭК, Ли С.Дж., Ким К.Г., Ким Й.Х., Ли С., Ли Б.И., Ким КТ (2014). «Компонент конденсина NCAPG2 регулирует прикрепление микротрубочек к кинетохорам посредством привлечения Polo-подобной киназы 1 к кинетохорам» . Нат Коммун . 5 : 4588. дои : 10.1038/ncomms5588 . ПМИД 25109385 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кагами Ю, Нихира К, Вада С, Оно М, Хонда М, Ёсида К (2014). «Фосфорилирование конденсина II Mps1 контролирует конденсацию хромосом в начале митоза» . Дж. Клеточная Биол . 205 (6): 781–790. дои : 10.1083/jcb.201308172 . ПМК 4068140 . ПМИД 24934155 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Йеонг Ф.М., Хомбауэр Х., Вендт К.С., Хирота Т., Мудрак И., Мехтлер К., Лореггер Т., Марчлер-Бауэр А., Танака К., Петерс Дж.М., Огрис Э. (2003). «Идентификация субъединицы нового комплекса клейзин-бета/SMC как потенциального субстрата протеинфосфатазы 2А» . Курр Биол . 13 (23): 2058–2064. дои : 10.1016/j.cub.2003.10.032 . ПМИД 14653995 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бастер Д.В., Дэниел С.Г., штаб-квартира Нгуена, Виндлер С.Л., Скварек Л.С., Петерсон М., Робертс М., Месерв Дж.Х., Хартл Т., Клебба Дж.Э., Билдер Д., Боско Г., Роджерс Г.К. (2013). «Убиквитинлигаза SCFSlimb подавляет ядерную реорганизацию, опосредованную конденсином II, путем разрушения Cap-H2» . Дж. Клеточная Биол . 201 (1): 49–63. дои : 10.1083/jcb.201207183 . ПМЦ 3613687 . ПМИД 23530065 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ямашита Д., Синтоми К., Оно Т., Гаввовидис И., Шиндлер Д., Нейцель Х., Тримборн М., Хирано Т. (2011). «MCPH1 регулирует конденсацию и форму хромосом как составной модулятор конденсина II» . Дж. Клеточная Биол . 194 (6): 841–854. дои : 10.1083/jcb.201106141 . ПМК 3207293 . ПМИД 21911480 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тримборн М., Шиндлер Д., Нейцель Х., Хирано Т. (2006). «Неправильная регуляция конденсации хромосом при первичной микроцефалии MCPH1 опосредована конденсином II» . Клеточный цикл . 5 (3): 322–326. дои : 10.4161/cc.5.3.2412 . ПМИД 16434882 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мартин К.А., Мюррей Дж.Э., Кэрролл П., Лейтч А., Маккензи К.Дж., Халачев М., Фетит А.Е., Кейт С., Бикнелл Л.С., Флюто А., Готье П., Холл Э.А., Джосс С., Соарес Г., Силва Дж., Бобер М.Б., Дукер А. , Wise CA, Quigley AJ, Phadke SR, «Расшифровка исследования нарушений развития», Wood AJ, Vagnarelli P, Jackson AP (2016). «Мутации в генах, кодирующих белки конденсинового комплекса, вызывают микроцефалию из-за нарушения декатенации при митозе» . Генс Дев . 30 (19): 2158–2172. дои : 10.1101/gad.286351.116 . ПМК 5088565 . ПМИД 27737959 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Гослинг К.М., Макаров Л.Е., Теодоратос А., Ким Ю.Х., Уиттл Б., Руи Л., Ву Х., Хонг Н.А., Кеннеди Г.К., Фриц Дж.А., Йейтс А.Л., Гуднау К.С., Фахрер А.М. (2007). «Мутация в субъединице конденсина II хромосомы, бета-клезине, специфически нарушает развитие Т-клеток» . Учеб. Натл. акад. наук. США . 104 (30): 12445–12450. Бибкод : 2007PNAS..10412445G . дои : 10.1073/pnas.0704870104 . ЧВК 1941488 . ПМИД 17640884 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Вудворд Дж., Тейлор Г.К., Соарес Д.С., Бойл С., Си Д., Рид Д., Чатот К., Вукович М., Тарратс Н., Джеймисон Д., Кэмпбелл К.Дж., Блит К., Акоста Дж.С., Илстра Б., Арендс М.Дж., Кранц К.Р., Джексон А.П. , Бикмор В.А., Вуд А.Дж. (2016). «Мутация конденсина II вызывает Т-клеточную лимфому из-за тканеспецифической нестабильности генома» . Генс Дев . 30 (19): 2173–2186. дои : 10.1101/gad.284562.116 . ПМК 5088566 . ПМИД 27737961 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хёнкамп С, Дудченко О, Эльбатш АМО, Брахмачари С, Рааймакерс Х.А., ван Шайк Т., Седеньо Каччиаторе А., Контессото В.Г., ван Хесбин РГХП, ван ден Брук Б., Мхаскар А.Н., Теуниссен Х., Сент-Хилэр Б.Г., Вайс Д., Омер А.Д. , Фам М, Коларик З, Ян З, Рао ССП, Митра Н, Луи С, Яо В, Хан Р, Мороз ЛЛ, Кон А, Сент-Леже Дж, Мена А, Холкрофт К, Гамбетта MC, Лим Ф, Фарли Е, Стейн Н., Хаддад А., Чаусс Д., Мутлу А.С., Ван М.К., Янг Н.Д., Хильдебрандт Э., Ченг Х.Х., Найт С.Дж., Бёрнем ТЛУ, Ховел К.А., Бил А.Дж., Маттеи П.Дж., Корнберг Р.Д., Уоррен У.К., Кэри Дж., Гомес- Скармета Дж.Л., Хинман В., Линдблад-Тох К., Ди Пальма Ф., Маешима К., Мултани А.С., Патхак С., Нел-Темаат Л., Берингер Р.Р., Каур П., Медема Р.Х., ван Стинсел Б., де Вит Е, Онучич Дж.Н., Ди Пьеро М., Либерман Эйден Э., Роуленд Б.Д. (2021). «3D-геномика на древе жизни показывает, что конденсин II является определяющим фактором типа архитектуры» . Наука . 372 (6545): 984–989. дои : 10.1126/science.abe2218 . ПМК 8172041 . ПМИД 34045355 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )
Внешние ссылки
[ редактировать ]- конденсин в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)