Профаза
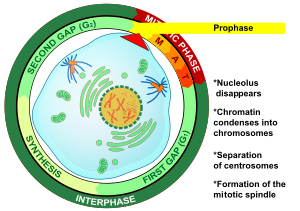

Профаза (от древнегреческого προ- ( про- ) «до» и φάσις (phásis) «появление») — первая стадия деления клеток как в митозе , так и в мейозе . Начиная с интерфазы , ДНК уже реплицируется, когда клетка входит в профазу. Основными явлениями в профазе являются конденсация хроматиновой сети и исчезновение ядрышка . [3]
Окрашивание и микроскопия
[ редактировать ]Микроскопию можно использовать для визуализации конденсированных хромосом по мере их прохождения через мейоз и митоз . [4]
различные пятна Для обработки клеток используются конденсацию хромосом по мере их прохождения через профазу. ДНК, позволяющие визуализировать [4]
Методика Гимзы G-бэндинга обычно используется для идентификации млекопитающих хромосом , но использование этой технологии на растительных клетках изначально было затруднено из-за высокой степени уплотнения хромосом в растительных клетках. [5] [4] G-бэндинг был полностью реализован для хромосом растений в 1990 году. [6] Как во время мейотической , так и в митотической профазе окрашивание по Гимзе можно применять к клеткам, чтобы вызвать появление G-полосок в хромосомах . [2] Окрашивание серебром, более современная технология, в сочетании с окрашиванием по Гимзе может использоваться для изображения синаптонемного комплекса на различных стадиях профазы мейоза . [7] Чтобы выполнить G-бэндинг , хромосомы должны быть зафиксированы, поэтому его невозможно выполнить на живых клетках. [8]
Флуоресцентные красители, такие как DAPI, можно использовать как в живых растительных , так и в животных клетках . Эти пятна не объединяют хромосомы , а вместо этого позволяют исследовать ДНК определенных областей и генов . Использование флуоресцентной микроскопии значительно улучшило пространственное разрешение . [9]
Митотическая профаза
[ редактировать ]Профаза — первая стадия митоза в животных клетках и вторая стадия митоза в растительных клетках . [10] В начале профазы в клетке имеются две идентичные копии каждой хромосомы вследствие репликации в интерфазе . Эти копии называются сестринскими хроматидами и присоединяются к ДНК элементу , называемому центромерой . [11] Основными событиями профазы являются: конденсация хромосом , движение центросом , образование митотического веретена и начало ядрышек . распада [3]
Конденсация хромосом
[ редактировать ]ДНК , реплицировавшаяся конденсируется в интерфазе, из нитей ДНК длиной до 0,7 мкм до 0,2-0,3 мкм. [3] В этом процессе используется конденсиновый комплекс. [11] Конденсированные хромосомы состоят из двух сестринских хроматид, соединенных в центромере . [12]
Движение центросом
[ редактировать ]Во время профазы в клетках животных центросомы раздвигаются достаточно далеко, чтобы их можно было различить с помощью светового микроскопа . [3] Активность микротрубочек в каждой центросоме увеличивается за счет рекрутирования γ-тубулина . Реплицированные центросомы из интерфазы раздвигаются к противоположным полюсам клетки под действием ассоциированных с центросомами моторных белков . [13] Взаимноотмеченные межполярные микротрубочки каждой центросомы взаимодействуют друг с другом, помогая перемещать центросомы к противоположным полюсам. [13] [3]
Формирование митотического веретена
[ редактировать ]Микротрубочки, участвующие в интерфазном каркасе, разрушаются по мере разделения реплицированных центросом . [3] Движение центросом к противоположным полюсам сопровождается в животных клетках организацией микротрубочек (астр). каждой центриоли отдельных радиальных массивов [13] Межполярные микротрубочки обеих центросом взаимодействуют, объединяя наборы микротрубочек и образуя базовую структуру митотического веретена . [13] Растительные клетки не имеют центросом, и хромосомы могут образовывать ядра микротрубочек в митотический аппарат . [13] В растительных клетках микротрубочки собираются на противоположных полюсах и начинают формировать веретенообразный аппарат в местах, называемых фокусами. [10] Митотическое веретено имеет большое значение в процессе митоза и в конечном итоге разделяет сестринские хроматиды в метафазе . [3]
Начало распада ядрышек
[ редактировать ]Ядрышки начинают разрушаться в профазе, что приводит к прекращению производства рибосом. [3] Это указывает на перенаправление клеточной энергии с общего клеточного метаболизма на клеточное деление . [3] Ядерная оболочка во время этого процесса остается нетронутой. [10]
Профаза мейоза
[ редактировать ]Мейоз включает два раунда сегрегации хромосом и, таким образом, дважды подвергается профазе, что приводит к профазе I и профазе II. [12] Профаза I — самая сложная фаза мейоза, поскольку гомологичные хромосомы должны спариваться и обмениваться генетической информацией . [3] : 98 Профаза II очень похожа на профазу митоза . [12]
Профаза I
[ редактировать ]Профаза I делится на пять фаз: лептотена, зиготена, пахитена, диплотена и диакинез. В дополнение к событиям, которые происходят в профазе митоза , внутри этих фаз происходит несколько важных событий, таких как спаривание гомологичных хромосом и реципрокный обмен генетическим материалом между этими гомологичными хромосомами . Профаза I происходит с разной скоростью в зависимости от вида и пола . Многие виды прекращают мейоз в диплотене профазы I до овуляции . [3] : 98 У людей могут пройти десятилетия, поскольку ооциты остаются задержанными в профазе I только для того, чтобы быстро завершить мейоз I до овуляции . [12]
Лептотена
[ редактировать ]На первой стадии профазы I, лептотене (от греческого «нежный»), хромосомы начинают конденсироваться. Каждая хромосома находится в диплоидном состоянии и состоит из двух сестринских хроматид ; однако хроматин сестринских хроматид еще недостаточно конденсирован, чтобы его можно было различить в микроскопии . [3] : 98 Гомологичные участки внутри гомологичных пар хромосом начинают ассоциироваться друг с другом. [2]
Зиготена
[ редактировать ]Во второй фазе профазы I, зиготене (от греческого «конъюгация»), все хромосомы, полученные по материнской и отцовской линии, нашли своего гомологичного партнера. [3] : 98 Затем гомологичные пары подвергаются синапсу - процессу, посредством которого синаптонемный комплекс (белковая структура) выравнивает соответствующие области генетической информации , полученных по материнской и отцовской линии на несестринских хроматидах гомологичных пар хромосом . [3] : 98 [12] Парные гомологичные хромосомы, связанные синаптонемным комплексом, называются бивалентами или тетрадами. [10] [3] : 98 Половые (X и Y) хромосомы не образуют синапсов полностью, поскольку гомологичны только небольшие участки хромосом. [3] : 98
Ядрышко перемещается из центрального положения в ядро на периферическое . [14]
Пахитена
[ редактировать ]Третья фаза профазы I, пахитена (от греческого «толстый»), начинается после завершения синапса. [3] : 98 Хроматин конденсировался настолько, что теперь хромосомы можно различить в микроскопии . [10] На синаптонемном комплексе бивалентов образуются структуры , называемые рекомбинационными узелками . Эти рекомбинационные узелки облегчают генетический обмен между несестринскими хроматидами синаптонемного комплекса в процессе, известном как кроссинговер или генетическая рекомбинация. [3] : 98 На каждом биваленте могут происходить множественные события рекомбинации. У человека на каждой хромосоме происходит в среднем 2-3 события. [13] : 681
Диплотена
[ редактировать ]В четвертой фазе профазы I, диплотене (от греческого «двойной»), кроссинговер . завершается [3] : 99 [10] Гомологичные хромосомы сохраняют полный набор генетической информации; однако гомологичные хромосомы теперь имеют смешанное материнское и отцовское происхождение. [3] : 99 Видимые соединения, называемые хиазмами, удерживают гомологичные хромосомы вместе в местах, где произошла рекомбинация при растворении синаптонемного комплекса . [12] [3] : 99 Именно на этой стадии у многих видов происходит остановка мейоза . [3] : 99
Диакинез
[ редактировать ]В пятой и последней фазе профазы I, диакинезе (от греческого «двойное движение»), произошла полная конденсация хроматина, и все четыре сестринские хроматиды можно увидеть в бивалентах при микроскопии . Остальная часть фазы напоминает ранние стадии митотической прометафазы , поскольку мейотическая профаза заканчивается веретенообразного аппарата началом формирования ядерной мембраны . и началом разрушения [10] [3] : 99
Профаза II
[ редактировать ]Профаза II мейоза очень похожа на профазу митоза . Наиболее заметное отличие состоит в том, что профаза II протекает с гаплоидным числом хромосом , а не с диплоидным числом в митотической профазе. [12] [10] Как в животных , так и в растительных клетках хромосомы могут деконденсироваться во время телофазы I, что требует их повторной конденсации в профазе II. [3] : 100 [10] Если хромосомам не требуется повторная конденсация, профаза II часто протекает очень быстро, как это видно на модельном организме Arabidopsis . [10]
Профаза я арестовываю
[ редактировать ]Самки млекопитающих и птиц рождаются со всеми ооцитами, необходимыми для будущих овуляций, и эти ооциты задерживаются на стадии профазы I мейоза . [15] У человека, например, ооциты образуются между тремя и четырьмя месяцами беременности внутри плода и, следовательно, присутствуют при рождении. Во время этой профазы I арестованной стадии ( диктиата ), которая может длиться десятилетиями, четыре копии генома в ооцитах присутствуют . Адаптивное значение остановки профазы I до сих пор до конца не изучено. Однако было высказано предположение, что арест ооцитов на стадии четырех копий генома может обеспечить информационную избыточность, необходимую для восстановления повреждений ДНК зародышевой линии . [15] Используемый процесс репарации, по-видимому, представляет собой гомологичную рекомбинационную репарацию. [15] [16] Ооциты с остановкой профазы обладают высокой способностью к эффективному восстановлению повреждений ДНК . [16] Способность к репарации ДНК, по-видимому, является ключевым механизмом контроля качества женской зародышевой линии и важнейшим фактором, определяющим фертильность . [16]
Различия в профазе растительных и животных клеток
[ редактировать ]
Наиболее заметное различие между профазой в растительных и животных клетках возникает из-за отсутствия центриолей в растительных клетках . Вместо этого организация веретенообразного аппарата связана с очагами на противоположных полюсах клетки или опосредована хромосомами. Другим заметным отличием является препрофаза , дополнительный этап митоза растений , который приводит к образованию препрофазной полосы , структуры, состоящей из микротрубочек . В профазе I митоза растений эта полоса исчезает. [10]
Контрольно-пропускные пункты сотовой связи
[ редактировать ]Профаза I в мейозе — наиболее сложная итерация профазы, которая встречается как в растительных, так и в животных клетках . [3] Чтобы обеспечить спаривание гомологичных хромосом и рекомбинацию генетического материала правильное , существуют клеточные контрольные точки . Сеть мейотических контрольных точек представляет собой систему реагирования на повреждение ДНК , которая контролирует двухцепочечных разрывов восстановление , структуру хроматина , а также движение и спаривание хромосом . [17] Система состоит из нескольких путей (включая контрольную точку мейотической рекомбинации ), которые не позволяют клетке войти в метафазу I с ошибками из-за рекомбинации. [18]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Нуссбаум Р.Л., Макиннес Р.Р., Хантингтон Ф. (2016). Томпсон и Томпсон Генетика в медицине . Филадельфия: Эльзевир . стр. 12–20. ISBN 9781437706963 .
- ^ Jump up to: а б с Шермелле Л., Карлтон П.М., Хаасе С., Шао Л., Виното Л., Кнер П. и др. (июнь 2008 г.). «Субдифракционная многоцветная визуализация ядерной периферии с помощью трехмерной микроскопии со структурированным освещением» . Наука . 320 (5881): 1332–36. Бибкод : 2008Sci...320.1332S . дои : 10.1126/science.1156947 . ПМЦ 2916659 . ПМИД 18535242 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и Хартвелл Л.Х., Худ Л., Голдберг М.Л., Рейнольдс А.Е., Сильвер Л.М., Верес Р.К. (2008). Генетика От генов к геномам . Нью-Йорк: МакГроу-Хилл. стр. 90–103 . ISBN 978-0-07-284846-5 .
- ^ Jump up to: а б с Сингх Р.Дж. (2017). Цитогенетика растений (Третье изд.). Бока-Ратон, Флорида: CBC Press, Taylor & Francisco Group. п. 19. ISBN 9781439884188 .
- ^ Ван ХК, Као К.Н. (1988). «G-бэндинг в хромосомах растений». Геном . 30 : 48–51. дои : 10.1139/g88-009 . S2CID 83823255 – через ResearchGate.
- ^ Какеда К., Ямагата Х., Фукуи К., Оно М., Фукуи К., Вэй З.З., Чжу Э.С. (август 1990 г.). «Полосы высокого разрешения в хромосомах кукурузы, полученные методами G-бэндирования». Теоретическая и прикладная генетика . 80 (2): 265–72. дои : 10.1007/BF00224397 . ПМИД 24220906 . S2CID 6600449 .
- ^ Патхак С., Сюй Т.С. (январь 1979 г.). «Окрашенные серебром структуры в профазе мейоза млекопитающих». Хромосома . 70 (2): 195–203. дои : 10.1007/bf00288406 . ПМИД 85512 . S2CID 27763957 .
- ^ Самнер А.Т. (май 1982 г.). «Природа и механизмы объединения хромосом». Генетика рака и цитогенетика . 6 (1): 59–87. дои : 10.1016/0165-4608(82)90022-x . ПМИД 7049353 .
- ^ де Йонг Х (декабрь 2003 г.). «Визуализация доменов и последовательностей ДНК с помощью микроскопии: пятидесятилетняя история молекулярной цитогенетики». Геном . 46 (6): 943–6. дои : 10.1139/g03-107 . ПМИД 14663510 .
- ^ Jump up to: а б с д и ж г час я дж к Тайз Л., Зейгер Э., Моллер И.М., Мерфи А. (2015). Физиология и развитие растений . Сандерленд, Массачусетс: Sinauer Associates. стр. 35–39. ISBN 978-1-60535-255-8 .
- ^ Jump up to: а б Цзэн XL, Цзяо МД, Ван XG, Сун ZX, Рао С (2001). «Электронно-микроскопические исследования окрашенного серебром ядрышкового цикла Physarum Polycephalum» (PDF) . Акта Ботаника Синика . 43 (7): 680–5. Архивировано из оригинала (PDF) 1 октября 2018 г. Проверено 24 февраля 2015 г.
- ^ Jump up to: а б с д и ж г Нуссбаум Р.Л., Макиннес Р.Р., Уиллард Х.Ф. (2016). Томпсон и Томпсон Генетика в медицине . Филадельфия: Эльзевир. стр. 12–20. ISBN 978-1-4377-0696-3 .
- ^ Jump up to: а б с д и ж Альбертс Б., Брэй Д., Хопкин К., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2004). Основная клеточная биология . Нью-Йорк, штат Нью-Йорк: Garland Science. стр. 639–658 . ISBN 978-0-8153-3481-1 .
- ^ Циклер Д., Клекнер Н. (1998). «Лептотен-зиготенный переход мейоза». Ежегодный обзор генетики . 32 : 619–97. дои : 10.1146/annurev.genet.32.1.619 . ПМИД 9928494 .
- ^ Jump up to: а б с Мира А. (сентябрь 1998 г.). «Почему останавливается мейоз?». Журнал теоретической биологии . 194 (2): 275–87. Бибкод : 1998JThBi.194..275M . дои : 10.1006/jtbi.1998.0761 . ПМИД 9778439 .
- ^ Jump up to: а б с Стрингер Дж. М., Уиншип А., Зерафа Н., Уэйкфилд М., Хатт К. (май 2020 г.). «Ооциты могут эффективно восстанавливать двухцепочечные разрывы ДНК, восстанавливая генетическую целостность и защищая здоровье потомства» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (21): 11513–11522. Бибкод : 2020PNAS..11711513S . дои : 10.1073/pnas.2001124117 . ПМК 7260990 . ПМИД 32381741 .
- ^ Хохваген А, Амон А (март 2006 г.). «Проверка ваших разрывов: механизмы наблюдения за мейотической рекомбинацией» . Современная биология . 16 (6): Р217-28. дои : 10.1016/j.cub.2006.03.009 . ПМИД 16546077 .
- ^ МакКуин Эй Джей, Хохваген А (июль 2011 г.). «Механизмы контрольно-пропускных пунктов: кукловоды профазы мейоза». Тенденции в клеточной биологии . 21 (7): 393–400. дои : 10.1016/j.tcb.2011.03.004 . ПМИД 21531561 .
Внешние ссылки
[ редактировать ] СМИ, связанные с профазой, на Викискладе?
СМИ, связанные с профазой, на Викискладе?