Центромера

Центромера хроматид связывает вместе пару сестринских во время деления клетки. Этот суженный участок хромосомы соединяет сестринские хроматиды, образуя короткое плечо (p) и длинное плечо (q) на хроматидах. Во время митоза прикрепляются волокна веретена к центромере через кинетохор .
Физическая роль центромеры заключается в том, чтобы действовать как место сборки кинетохор – очень сложной мультибелковой структуры, которая отвечает за фактические события сегрегации хромосом – т.е. связывание микротрубочек и передачу сигналов механизму клеточного цикла, когда все хромосомы приняли правильное положение. прикрепляются к веретену , так что деление клеток может безопасно завершиться и клетки вступают в анафазу .
Вообще говоря, существует два типа центромер. «Точечные центромеры» связываются со специфическими белками , которые с высокой эффективностью распознают определенные ДНК последовательности . Любой участок ДНК с последовательностью ДНК точечной центромеры обычно образует центромеру, если она присутствует у соответствующего вида. Лучше всего охарактеризованы точечные центромеры почкующихся дрожжей Saccharomyces cerevisiae . «Региональные центромеры» — это термин, придуманный для описания большинства центромер, которые обычно образуются на участках предпочтительной последовательности ДНК, но могут образовываться и на других последовательностях ДНК. Сигнал к образованию региональной центромеры, по-видимому, является эпигенетическим . Большинство организмов, от делящихся дрожжей Schizosaccharomyces pombe до человека, имеют регионарные центромеры.
Что касается структуры митотической хромосомы, центромеры представляют собой суженную область хромосомы (часто называемую первичной перетяжкой), где две идентичные сестринские хроматиды наиболее тесно контактируют. Когда клетки вступают в митоз, сестринские хроматиды (две копии каждой хромосомной молекулы ДНК, образующиеся в результате репликации ДНК в форме хроматина) соединяются по своей длине под действием комплекса когезина . В настоящее время считается, что этот комплекс в основном высвобождается из плеч хромосом во время профазы, так что к тому времени, когда хромосомы выстраиваются в средней плоскости митотического веретена (также известного как метафазная пластинка), последнем месте, где они соединяются. друг с другом находится в хроматине внутри и вокруг центромеры.
Должность [ править ]
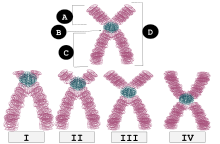
| я | Телоцентрический | Центромера расположена очень близко к верху, плечи почти не видны, если вообще видны. |
| II | Акроцентрический | q-плечи по-прежнему намного длиннее, чем p-плечи, но p-плечи длиннее, чем у телоцентрических. |
| III | Субметацентрический | Плечи p и q очень близки по длине, но не равны. |
| IV | Метацентрический | Плечи p и q имеют одинаковую длину. |
Б : Центромера
C : Длинная рука (q рука)
D : Сестринские хроматиды.
У человека положения центромеры определяют хромосомный кариотип , в котором каждая хромосома имеет два плеча: p (более короткое из двух) и q (более длинное). Сообщается, что короткая рука «p» названа в честь французского слова «petit», что означает «маленький». [1] Положение центромеры относительно любой конкретной линейной хромосомы используется для классификации хромосом на метацентрические, субметацентрические, акроцентрические, телоцентрические или голоцентрические. [2] [3]
| Классификация хромосом по относительной длине плеч. [3] | ||||||
| Положение центромеры | Соотношение длин рук | Знак | Описание | |||
| Медиальный смысл в строгом смысле | 1.0 – 1.6 | М | Метацентрический | |||
| Медиальная область | 1.7 | м | Метацентрический | |||
| субмедиальный | 3.0 | см | Субметацентрический | |||
| субтерминальный | 3.1 – 6.9 | ул. | Субтелоцентрический | |||
| Терминальный регион | 7.0 | т | Акроцентрический | |||
| Терминал в строгом смысле слова | ∞ | Т | Телоцентрический | |||
| Примечания | – | Метацентрический : М + м | Ателоцентрический : M + m + sm + st + t | |||
Метацентрический [ править ]
Метацентрический означает, что центромера расположена посередине между концами хромосом, в результате чего плечи примерно одинаковой длины. Когда центромеры метацентричны, хромосомы кажутся «х-образными».
Субметацентрический [ править ]
Субметацентричность означает, что центромера расположена ниже середины, при этом одно плечо хромосомы короче другого, что часто приводит к L-образной форме.
Акроцентрический [ править ]
Центромера акроцентрической хромосомы расположена так, что одно из плеч хромосомы значительно короче другого. Слово «акро-» в слове «акроцентрический» относится к греческому слову «пик». Геном человека имеет шесть акроцентрических хромосом, включая пять аутосомных хромосом ( 13 , 14 , 15 , 21 , 22 ) и Y-хромосому .
Короткие акроцентрические p-плечи содержат мало генетического материала и могут быть транслоцированы без значительного вреда, как при сбалансированной робертсоновской транслокации . В дополнение к некоторым генам, кодирующим белки, акроцентрические p-плечи человека также содержат области ядрышкового организатора (NOR), из которых рибосомальная РНК транскрибируется . Однако часть акроцентрических p-плечей в клеточных линиях и тканях нормальных доноров-человеков не содержит обнаруживаемых NOR. [4] Геном домашней лошади включает одну метацентрическую хромосому, гомологичную двум акроцентрическим хромосомам конспецифичной, но неодомашненной лошади Пржевальского . Это может отражать либо фиксацию сбалансированной робертсоновской транслокации у домашних лошадей, либо, наоборот, фиксацию деления одной метацентрической хромосомы на две акроцентрические хромосомы у лошадей Пржевальского. Аналогичная ситуация существует между геномами человека и человекообразных обезьян, с редукцией двух акроцентрических хромосом у человекообразных обезьян до одной метацентрической хромосомы у человека (см. анеуплоидия и хромосома человека 2 ).
Многие заболевания, возникающие в результате несбалансированных транслокаций, чаще поражают акроцентрические хромосомы, чем другие неакроцентрические хромосомы. Акроцентрические хромосомы обычно располагаются внутри и вокруг ядрышка . В результате эти хромосомы имеют тенденцию быть менее плотно упакованными, чем хромосомы на ядерной периферии. Соответственно, хромосомные области, которые менее плотно упакованы, также более склонны к хромосомным транслокациям при раке.
Телоцентрический [ править ]
Телоцентрические хромосомы имеют центромеру на одном конце хромосомы и поэтому на цитологическом (микроскопическом) уровне имеют только одно плечо. Они отсутствуют у людей, но могут образовываться в результате клеточных хромосомных ошибок. Телоцентрические хромосомы встречаются в природе у многих видов, например, у домовой мыши , у которой все хромосомы, кроме Y, являются телоцентрическими.
Субтелоцентрический [ править ]
Центромеры субтелоцентрических хромосом расположены между серединой и концом хромосом, но располагаются ближе к концу хромосом.
Типы центромер [ править ]
Ацентрический [ править ]
Ацентрическая хромосома – это фрагмент хромосомы, лишенный центромеры. Поскольку центромеры являются точкой прикрепления волокон веретена при делении клеток, ацентрические фрагменты неравномерно распределяются по дочерним клеткам во время клеточного деления. В результате в дочерней клетке будет отсутствовать ацентрический фрагмент, и могут возникнуть вредные последствия.
События, разрушающие хромосомы, также могут генерировать ацентрические хромосомы или ацентрические фрагменты.
Дицентрик [ править ]
Дицентрическая хромосома — это аномальная хромосома с двумя центромерами, которая может быть нестабильной в результате деления клеток. Он может образовываться путем транслокации или слияния двух сегментов хромосом, каждый из которых имеет центромеру. Некоторые перестройки приводят к образованию как дицентрических хромосом, так и ацентрических фрагментов, которые не могут прикрепляться к веретену при митозе. [5] Образование дицентрических хромосом связывают с генетическими процессами, такими как робертсоновская транслокация. [6] и парацентрическая инверсия. [7] Дицентрические хромосомы могут иметь различную судьбу, включая митотическую стабильность. [8] В некоторых случаях их стабильность достигается за счет инактивации одной из двух центромер, что приводит к созданию функционально моноцентрической хромосомы, способной к нормальной передаче дочерним клеткам во время клеточного деления. [9]
Например, человеческая хромосома 2 , которая, как полагают, является результатом робертсоновской транслокации в какой-то момент эволюции между человекообразными обезьянами и человеком , имеет вторую, рудиментарную центромеру около середины ее длинного плеча. [10]
Моноцентрический [ править ]
Моноцентрическая хромосома — это хромосома, которая имеет только одну центромеру в хромосоме и образует узкую перетяжку.
Моноцентрические центромеры являются наиболее распространенной структурой высокоповторяющейся ДНК у растений и животных. [11]
Голоцентрический [ править ]
В отличие от моноцентрических хромосом, голоцентрические хромосомы не имеют отчетливой первичной перетяжки, если смотреть на них в митозе. Вместо этого волокна веретена прикрепляются почти по всей (греч. голо-) длине хромосомы. В голоцентрических хромосомах центромерные белки, такие как CENPA (CenH3), распределены по всей хромосоме. [12] Нематода Caenorhabditis elegans — хорошо известный пример организма с голоцентрическими хромосомами. [13] но этот тип центромеры можно найти у различных видов, растений и животных, среди эукариот. Голоцентромеры на самом деле состоят из множества распределенных центромерных единиц, которые образуют линейную структуру вдоль хромосом во время митоза. [14] Альтернативные или нетрадиционные стратегии используются при мейозе для достижения гомологичного спаривания и разделения хромосом, необходимого для производства жизнеспособных гамет или гаметофитов для полового размножения.
У разных видов существуют разные типы голоцентромер, а именно с центромерными повторяющимися последовательностями ДНК или без них, а также с CenH3 или без него . Голоцентричность развивалась по крайней мере 13 раз независимо у различных зеленых водорослей, простейших, беспозвоночных и различных семейств растений. [15] В отличие от моноцентрических видов, у которых ацентрические фрагменты обычно теряются во время деления клеток, при разрыве голоцентрических хромосом образуются фрагменты с нормальными местами прикрепления волокон веретена. [16] Из-за этого организмы с голоцентрическими хромосомами могут быстрее развивать вариации кариотипа, способные восстанавливать фрагментированные хромосомы путем последующего добавления колпачков теломер в местах разрыва. [17]
Полицентрический [ править ]
Этот раздел пуст. Вы можете помочь, добавив к нему . ( сентябрь 2023 г. ) |
Хромосомы человека [ править ]

| хромосома | Центромера позиция ( Мбит / с ) | Категория | хромосома Размер (Мбит/с) | Центромера размер (Мбит/с) |
|---|---|---|---|---|
| 1 | 125.0 | метацентрический | 247.2 | 7.4 |
| 2 | 93.3 | субметацентрический | 242.8 | 6.3 |
| 3 | 91.0 | метацентрический | 199.4 | 6.0 |
| 4 | 50.4 | субметацентрический | 191.3 | — |
| 5 | 48.4 | субметацентрический | 180.8 | — |
| 6 | 61.0 | субметацентрический | 170.9 | — |
| 7 | 59.9 | субметацентрический | 158.8 | — |
| 8 | 45.6 | субметацентрический | 146.3 | — |
| 9 | 49.0 | субметацентрический | 140.4 | — |
| 10 | 40.2 | субметацентрический | 135.4 | — |
| 11 | 53.7 | субметацентрический | 134.5 | — |
| 12 | 35.8 | субметацентрический | 132.3 | — |
| 13 | 17.9 | акроцентрический | 114.1 | — |
| 14 | 17.6 | акроцентрический | 106.3 | — |
| 15 | 19.0 | акроцентрический | 100.3 | — |
| 16 | 36.6 | метацентрический | 88.8 | — |
| 17 | 24.0 | субметацентрический | 78.7 | — |
| 18 | 17.2 | субметацентрический | 76.1 | — |
| 19 | 26.5 | метацентрический | 63.8 | — |
| 20 | 27.5 | метацентрический | 62.4 | — |
| 21 | 13.2 | акроцентрический | 46.9 | — |
| 22 | 14.7 | акроцентрический | 49.5 | — |
| Х | 60.6 | субметацентрический | 154.9 | — |
| И | 12.5 | акроцентрический | 57.7 | — |
На основании микрографических характеристик размера, положения центромеры и иногда наличия хромосомного спутника хромосомы человека классифицируются на следующие группы: [18]
| Группа | Хромосомы | Функции |
|---|---|---|
| Группа А | Хромосома 1-3 | Большие, метацентрические и субметацентрические |
| Группа Б | Хромосома 4-5 | Большой, субметацентрический |
| Группа С | Хромосома 6-12, Х | Среднего размера, субметацентрический |
| Группа Д | Хромосома 13-15 | Среднего размера, акроцентрическая, со сателлитом. |
| Группа Е | Хромосома 16-18 | Малые, метацентрические и субметацентрические |
| Группа F | Хромосома 19-20 | Очень маленький, метацентрический |
| Группа G | Хромосома 21-22, Y | Очень маленький, акроцентрический, со сателлитом. |
Последовательность [ править ]
Существует два типа центромер. [19] В региональных центромерах последовательности ДНК вносят вклад в функцию, но не определяют ее. Региональные центромеры содержат большое количество ДНК и часто упакованы в гетерохроматин . У большинства эукариот последовательность ДНК центромеры состоит из больших массивов повторяющейся ДНК (например, сателлитной ДНК ), где последовательность отдельных повторяющихся элементов схожа, но не идентична. У людей первичная центромерная повторяющаяся единица называется α-сателлитом (или альфоидом), хотя в этой области обнаружен ряд других типов последовательностей. [20] Предполагается, что сателлиты центромеры развиваются в результате процесса, называемого послойным расширением. Они быстро эволюционируют между видами, и анализы на диких мышах показывают, что количество сателлитных копий и гетерогенность связаны с происхождением популяции и подвидами. [21] Кроме того, инбридинг может повлиять на сателлитные последовательности. [21]
Точечные центромеры меньше и компактнее. Последовательности ДНК необходимы и достаточны для определения идентичности и функции центромер у организмов с точечными центромерами. У почкующихся дрожжей центромерная область относительно мала (около 125 п.о. ДНК) и содержит две высококонсервативные последовательности ДНК, которые служат сайтами связывания для важных белков кинетохор . [20]
Наследование [ править ]
Поскольку последовательность центромерной ДНК не является ключевым фактором, определяющим идентичность центромер у многоклеточных животных , считается, что эпигенетическая наследственность играет важную роль в спецификации центромеры. [22] Дочерние хромосомы будут собирать центромеры в том же месте, что и родительская хромосома, независимо от последовательности. Было высказано предположение, что вариант гистона H3 CENP-A (центромерный белок A) является эпигенетической меткой центромеры. [23] Возникает вопрос, должен ли существовать еще какой-то оригинальный способ спецификации центромеры, даже если он впоследствии размножается эпигенетически. Если центромера наследуется эпигенетически от одного поколения к другому, проблема возвращается к происхождению первых многоклеточных животных.
С другой стороны, благодаря сравнению центромер Х-хромосом в этих регионах были обнаружены эпигенетические и структурные вариации. Кроме того, недавняя сборка генома человека выявила возможный механизм эволюции прицентромерных и центромерных структур посредством многоуровневой модели расширения последовательностей αSat. Эта модель предполагает, что различные повторы последовательности αSat периодически возникают и расширяются внутри активного вектора, вытесняя старые последовательности и становясь местом сборки кинетохор. αSat может происходить от одного и того же вектора или от разных векторов. Поскольку этот процесс повторяется с течением времени, слои, окружающие активную центромеру, сжимаются и разрушаются. Этот процесс поднимает вопросы о связи этого динамического эволюционного процесса с положением центромеры. [24]
Структура [ править ]
Центромерная ДНК обычно находится в состоянии гетерохроматина , что важно для рекрутирования комплекса когезина , который обеспечивает слипание сестринских хроматид после репликации ДНК, а также координирует разделение сестринских хроматид во время анафазы. В этом хроматине нормальный гистон H3 заменен на центромер-специфичный вариант CENP-A у человека. [25] Считается, что присутствие CENP-A важно для сборки кинетохора на центромере. Было показано, что CENP-C локализуется почти исключительно в этих областях хроматина, связанного с CENP-A. Обнаружено, что в клетках человека гистоны наиболее обогащены H4K20me3 и H3K9me3. [26] которые являются известными гетерохроматическими модификациями. У дрозофилы островки ретроэлементов являются основными компонентами центромер. [27]
У дрожжей Schizosaccharomyces pombe (и, вероятно, у других эукариот) образование центромерного гетерохроматина связано с РНКи . [28] У нематод, таких как Caenorhabditis elegans , некоторых растений и насекомых отрядов Lepidoptera и Hemiptera, хромосомы «голоцентричны», что указывает на отсутствие первичного места прикрепления микротрубочек или первичного сужения, а по всей длине собирается «диффузная» кинетохора. длину хромосомы.
Центромерные аберрации [ править ]
В редких случаях неоцентромеры могут образовываться на новых участках хромосомы в результате изменения положения центромеры. Это явление наиболее хорошо известно из клинических исследований на людях, и в настоящее время известно более 90 неоцентромеров человека, идентифицированных на 20 различных хромосомах. [29] [30] Образование неоцентромеры должно сопровождаться инактивацией предыдущей центромеры, поскольку хромосомы с двумя функциональными центромерами ( дицентрическая хромосома ) приведут к разрыву хромосомы во время митоза. В некоторых необычных случаях неоцентромеры человека наблюдались спонтанно формируются на фрагментированных хромосомах. Некоторые из этих новых позиций изначально были эухроматическими и вообще не имели альфа-сателлитной ДНК. Неоцентромеры лишены повторяющейся структуры, наблюдаемой в нормальных центромерах, что указывает на то, что образование центромер в основном контролируется эпигенетически . [31] [32] Со временем неоцентромера может накапливать повторяющиеся элементы и превращаться в так называемую эволюционную новую центромеру. Есть несколько хорошо известных примеров в хромосомах приматов, где положение центромеры отличается от человеческой центромеры той же хромосомы и считается эволюционно новыми центромерами. [31] Было высказано предположение, что репозиция центромер и образование эволюционных новых центромер являются механизмом видообразования . [33]
Центромерные белки также являются аутоантигенной мишенью для некоторых антинуклеарных антител , таких как антицентромерные антитела .
Дисфункция и болезнь [ править ]
Известно, что неправильная регуляция центромер способствует неправильной сегрегации хромосом, что тесно связано с раком и выкидышем. Примечательно, что сверхэкспрессия многих центромерных генов связана со злокачественными фенотипами рака. Сверхэкспрессия этих центромерных генов может увеличить нестабильность генома при раке. Повышенная геномная нестабильность, с одной стороны, связана со злокачественными фенотипами; с другой стороны, это делает опухолевые клетки более уязвимыми к специфическим адъювантным методам лечения, таким как определенные виды химиотерапии и лучевой терапии. [34] Нестабильность повторяющейся центромерной ДНК недавно была показана при раке и старении. [35]
ДНК центромерной Репарация
Когда разрывы ДНК происходят в центромерах в фазе G1 клеточного цикла , клетки способны рекрутировать гомологичный рекомбинационный механизм репарации к поврежденному участку, даже в отсутствие сестринской хроматиды . [36] Похоже, что гомологичная рекомбинационная репарация может происходить при разрывах центромер на протяжении всего клеточного цикла, чтобы предотвратить активацию неточных путей мутагенной репарации ДНК и сохранить целостность центромер. [36]
Этимология и произношение [ править ]
Слово центромера ( / ˈ s ɛ n t r ə ˌ m ɪər / [37] [38] ) использует сочетание форм центро- и -мере , образуя «центральную часть», описывающую расположение центромеры в центре хромосомы.
См. также [ править ]
Ссылки [ править ]
- ^ «p + q = решено: это правдивая история о том, как хромосома получила свое название» . 03 мая 2011 г.
- ^ «Какие типы хромосом существуют?» , Уроки генетики Николая , YouTube, 12 октября 2013 г., заархивировано из оригинала 11 декабря 2021 г. , получено 28 мая 2017 г.
- ^ Jump up to: Перейти обратно: а б Леван А., Фредга К., Сандберг А.А. (декабрь 1964 г.). «Номенклатура центромерного положения хромосом». Эредитас . 52 (2): 201–220. дои : 10.1111/j.1601-5223.1964.tb01953.x .
- ^ ван Слуис М., ван Вуурен С., Манган Х., Макстей Б. (май 2020 г.). «NOR на p-плечах акроцентрической хромосомы человека активны по умолчанию и могут связываться с ядрышками независимо от рДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (19): 10368–10377. Бибкод : 2020PNAS..11710368V . дои : 10.1073/pnas.2001812117 . ПМЦ 7229746 . ПМИД 32332163 .
- ^ Нуссбаум Р., Макиннес Р., Уиллард Х., Хамош А. (2007). Томпсон и Томпсон Генетика в медицине . Филадельфия (Пенсильвания): Сондерс. п. 72. ИСБН 978-1-4160-3080-5 .
- ^ Генетика Томпсона и Томпсона в медицине (7-е изд.). п. 62.
- ^ Хартвелл Л., Худ Л., Голдберг М., Рейнольдс А., Ли С. (2011). Генетика от генов к геномам (4-е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 9780073525266 .
- ^ Линч С.А., Эшкрофт К.А., Зволински С., Кларк С., Берн Дж. (март 1995 г.). «Особенности, подобные синдрому Кабуки, у монозиготных мальчиков-близнецов с псевдодицентрической хромосомой 13» . Журнал медицинской генетики . 32 (3): 227–230. дои : 10.1136/jmg.32.3.227 . ПМК 1050324 . ПМИД 7783176 .
- ^ Стимпсон, Кейтлин М.; Матени, Джастин Э.; Салливан, Бет А. (2012). «Дицентрические хромосомы: уникальные модели для изучения функции и инактивации центромер» . Хромосомные исследования . 20 (5): 595–605. дои : 10.1007/s10577-012-9302-3 . ПМЦ 3557915 .
- ^ Аварелло; и др. (1992). «Доказательства наличия наследственного альфоидного домена на длинном плече хромосомы 2 человека». Генетика человека . 89 (2): 247–9. дои : 10.1007/BF00217134 . ПМИД 1587535 . S2CID 1441285 .
- ^ Барра В., Фачинетти Д. (октябрь 2018 г.). «Темная сторона центромер: типы, причины и последствия структурных аномалий, затрагивающих центромерную ДНК» . Природные коммуникации . 9 (1): 4340. Бибкод : 2018NatCo...9.4340B . дои : 10.1038/s41467-018-06545-y . ПМК 6194107 . ПМИД 30337534 .
- ^ Нойманн П., Навратилова А., Шредер-Рейтер Е., Коближкова А., Штайнбауэрова В., Чохолова Е. и др. (2012). «Расширяя правила: моноцентрические хромосомы с несколькими центромерными доменами» . PLoS Генетика . 8 (6): e1002777. дои : 10.1371/journal.pgen.1002777 . ПМЦ 3380829 . ПМИД 22737088 .
- ^ Дернбург А.Ф. (июнь 2001 г.). «Здесь, там и везде: функция кинетохор на голоцентрических хромосомах» . Журнал клеточной биологии . 153 (6): Ф33–Ф38. дои : 10.1083/jcb.153.6.F33 . ПМК 2192025 . ПМИД 11402076 .
- ^ Маркес А., Рибейро Т., Нейман П., Макас Дж., Новак П., Шуберт В. и др. (ноябрь 2015 г.). «Голоцентромеры у Rhynchospora связаны с массивами специфичных для центромер повторов по всему геному, вкрапленными среди эухроматина» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (44): 13633–13638. Бибкод : 2015PNAS..11213633M . дои : 10.1073/pnas.1512255112 . ПМЦ 4640781 . ПМИД 26489653 .
- ^ Мелтерс Д.П., Палиулис Л.В., Корф И.Ф., Чан С.В. (июль 2012 г.). «Голоцентрические хромосомы: конвергентная эволюция, мейотическая адаптация и геномный анализ» . Хромосомные исследования . 20 (5): 579–593. дои : 10.1007/s10577-012-9292-1 . ПМИД 22766638 . S2CID 3351527 .
- ^ Хьюз-Шрейдер С ; Рис Х (август 1941 г.). «Диффузное прикрепление веретена кокцид, подтвержденное митотическим поведением индуцированных фрагментов хромосом» . Журнал экспериментальной зоологии . 87 (3): 429–456. дои : 10.1002/jez.1400870306 . ISSN 0022-104X .
- ^ Янковска М., Фукс Дж., Клоке Е., Фойтова М., Поланска П., Файкус Дж. и др. (декабрь 2015 г.). «Голокинетические центромеры и эффективное заживление теломер обеспечивают быструю эволюцию кариотипа». Хромосома . 124 (4): 519–528. дои : 10.1007/s00412-015-0524-y . ПМИД 26062516 . S2CID 2530401 .
- ^ Эрвинсия Р., Рианди и Нурджани М. (2017). «Актуальность анализа хромосом человека в сравнении с концепцией мутаций в курсе генетики. Серия конференций IOP» . Материаловедение и инженерия . дои : 10.1088/1757-899x/180/1/012285 . S2CID 90739754 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Плута А.Ф., Маккей А.М., Айнштейн А.М., Гольдберг И.Г., Эрншоу В.К. (декабрь 1995 г.). «Центромера: центр хромосомной активности». Наука . 270 (5242): 1591–1594. Бибкод : 1995Sci...270.1591P . дои : 10.1126/science.270.5242.1591 . ПМИД 7502067 . S2CID 44632550 .
- ^ Jump up to: Перейти обратно: а б Мехта Г.Д., член парламента Агарвала, Гош С.К. (август 2010 г.). «Центромерная идентичность: проблема, с которой придется столкнуться». Молекулярная генетика и геномика . 284 (2): 75–94. дои : 10.1007/s00438-010-0553-4 . ПМИД 20585957 . S2CID 24881938 .
- ^ Jump up to: Перейти обратно: а б Арора У.П., Шарлебуа К., Лаваль Р.А., Дюмон Б.Л. (апрель 2021 г.). «Популяция и подвидовое разнообразие сателлитов центромеры мыши» . БМК Геномика . 22 (1): 279. дои : 10.1186/s12864-021-07591-5 . ПМЦ 8052823 . ПМИД 33865332 .
- ^ Далал Й. (февраль 2009 г.). «Эпигенетическая спецификация центромер». Биохимия и клеточная биология . 87 (1): 273–282. дои : 10.1139/O08-135 . ПМИД 19234541 .
- ^ Бернад Р., Санчес П., Лосада А. (ноябрь 2009 г.). «Эпигенетическая спецификация центромер с помощью CENP-A». Экспериментальные исследования клеток . 315 (19): 3233–3241. doi : 10.1016/j.yexcr.2009.07.023 . ПМИД 19660450 .
- ^ Альтемосе, Николас; Логсдон, Гленнис А.; Бзикадзе Андрей Владимирович; Сидхвани, Прагья; Лэнгли, Саша А.; Калдас, Джина В.; Хойт, Саванна Дж.; Уральский, Лев; Рябов Федор Дмитриевич; Шью, Колин Дж.; Саурия, Майкл Э.Г.; Борчерс, Мэтью; Гершман, Ариэль; Михеенко Алла; Шепелев, Валерий А. (апрель 2022 г.). «Полные геномные и эпигенетические карты центромер человека» . Наука . 376 (6588): eabl4178. дои : 10.1126/science.abl4178 . ISSN 0036-8075 . ПМЦ 9233505 . ПМИД 35357911 .
- ^ Чуэ А.С., Вонг Л.Х., Вонг Н., Чу К.Х. (январь 2005 г.). «Переменное и иерархическое распределение размеров кластеров CENP-A, обогащенных L1-ретроэлементами, внутри функциональной неоцентромеры человека» . Молекулярная генетика человека . 14 (1): 85–93. дои : 10.1093/hmg/ddi008 . ПМИД 15537667 .
- ^ Розенфельд Дж. А., Ван З., Шонес Д.Э., Чжао К., ДеСалле Р., Чжан М.К. (март 2009 г.). «Определение обогащенных модификаций гистонов в негенных частях генома человека» . БМК Геномика . 10 :143. дои : 10.1186/1471-2164-10-143 . ПМЦ 2667539 . ПМИД 19335899 .
- ^ Чанг Ч., Чаван А., Палладино Дж., Вэй Х., Мартинс Н.М., Сантинелло Б. и др. (май 2019 г.). «Островки ретроэлементов являются основными компонентами центромер дрозофилы» . ПЛОС Биология . 17 (5): e3000241. дои : 10.1371/journal.pbio.3000241 . ПМК 6516634 . ПМИД 31086362 .
- ^ Вольпе Т.А., Киднер С., Холл И.М., Тенг Г., Гревал С.И., Мартиенссен Р.А. (сентябрь 2002 г.). «Регуляция гетерохроматического молчания и метилирования гистона H3 лизина-9 с помощью РНКи» . Наука . 297 (5588): 1833–1837. Бибкод : 2002Sci...297.1833V . дои : 10.1126/science.1074973 . ПМИД 12193640 . S2CID 2613813 .
- ^ Маршалл О.Дж., Чуэ А.С., Вонг Л.Х., Чу К.Х. (февраль 2008 г.). «Неоцентромеры: новое понимание структуры центромер, развития заболеваний и эволюции кариотипа» . Американский журнал генетики человека . 82 (2): 261–282. дои : 10.1016/j.ajhg.2007.11.009 . ПМК 2427194 . ПМИД 18252209 .
- ^ Уорбертон PE (2004). «Хромосомная динамика формирования неоцентромеры человека». Хромосомные исследования . 12 (6): 617–626. дои : 10.1023/B:CHRO.0000036585.44138.4b . ПМИД 15289667 . S2CID 29472338 .
- ^ Jump up to: Перейти обратно: а б Рокки М., Арчидиаконо Н., Шемпп В., Капоцци О., Станьон Р. (январь 2012 г.). «Репозиция центромеры у млекопитающих» . Наследственность . 108 (1): 59–67. дои : 10.1038/hdy.2011.101 . ПМК 3238114 . ПМИД 22045381 .
- ^ Птолемей Д., Капоцци О., Станьон Р.Р., архидиакон Н., Д'Аддаббо П., Катаккио Ч.Р. и др. (февраль 2017 г.). «Эпигенетическое происхождение эволюционных новых центромер» . Научные отчеты 7 (1): 41980. Бибкод : 2017NatSR...741980T . дои : 10.1038/srep41980 . ПМК 5290474 . ПМИД 28155877 .
- ^ Браун ДжейДи, О'Нил Р.Дж. (сентябрь 2010 г.). «Хромосомы, конфликт и эпигенетика: новый взгляд на хромосомное видообразование». Ежегодный обзор геномики и генетики человека . 11 (1): 291–316. doi : 10.1146/annurev-genom-082509-141554 . ПМИД 20438362 .
- ^ Чжан В., Мао Дж.Х., Чжу В., Джайн А.К., Лю К., Браун Дж.Б., Карпен Г.Х. (август 2016 г.). «Неправильная экспрессия генов центромеры и кинетохор предсказывает выживаемость онкологических больных и реакцию на лучевую терапию и химиотерапию» . Природные коммуникации . 7 : 12619. Бибкод : 2016NatCo...712619Z . дои : 10.1038/ncomms12619 . ПМК 5013662 . ПМИД 27577169 .
- ^ Джунта С., Фунабики Х. (февраль 2017 г.). «Целостность повторов центромерной ДНК человека защищена CENP-A, CENP-C и CENP-T» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (8): 1928–1933. Бибкод : 2017ПНАС..114.1928Г . дои : 10.1073/pnas.1615133114 . ПМЦ 5338446 . ПМИД 28167779 .
- ^ Jump up to: Перейти обратно: а б Йилмаз Д., Фёрст А., Миберн К., Лезая А., Вэнь Ю., Альтмейер М., Рейна-Сан-Мартин Б., Сутоглу Э. (декабрь 2021 г.). «Активация гомологичной рекомбинации в G1 сохраняет целостность центромеры». Природа . 600 (7890): 748–753. Бибкод : 2021Natur.600..748Y . дои : 10.1038/s41586-021-04200-z . ПМИД 34853474 . S2CID 244800481 .
- ^ «Центромера» . Словарь Merriam-Webster.com .
- ^ «Центромера» . Dictionary.com Полный (онлайн). nd
Дальнейшее чтение [ править ]
- Мехта Г.Д., член парламента Агарвала, Гош С.К. (август 2010 г.). «Центромерная идентичность: проблема, с которой придется столкнуться». Молекулярная генетика и геномика . 284 (2): 75–94. дои : 10.1007/s00438-010-0553-4 . ПМИД 20585957 . S2CID 24881938 .
- Лодиш Х., Берк А., Кайзер К.А., Кайзер С., Кригер М., Скотт М.П., Бретчер А., Плух Х., Мацудайра (2008). Молекулярно-клеточная биология (6-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-7601-7 .
- Нагаки К., Ченг З., Оуян С., Талберт П.Б., Ким М., Джонс К.М. и др. (февраль 2004 г.). «Секвенирование центромеры риса обнаруживает активные гены» . Природная генетика . 36 (2): 138–145. дои : 10.1038/ng1289 . ПМИД 14716315 .
Внешние ссылки [ править ]
- «Центромера риса, предположительно тихий генетический домен, сюрпризы» . ScienceDaily (пресс-релиз). 13 января 2004 г.