Палеополиплоидия

[1] Палеополиплоидия — результат дупликации генома , произошедшей как минимум несколько миллионов лет назад (MYA). Такое событие может либо удвоить геном одного вида ( аутополиплоидия ), либо объединить геномы двух видов ( аллополиплоидия ). Из-за функциональной избыточности гены быстро замолкают или теряются из дублированных геномов. Большинство палеополиплоидов в процессе эволюции утратили свой полиплоидный статус в результате процесса, называемого диплоидизацией , и в настоящее время считаются диплоидами , например, пекарские дрожжи , [2] арабидопсис Талиана , [3] и, возможно, люди . [4] [5] [6] [7]
Палеополиплоидия широко изучается в линиях растений. Было обнаружено, что почти все цветковые растения в какой-то момент своей эволюционной истории претерпели хотя бы один раунд дупликации генома. Дупликации древнего генома также обнаружены у ранних предков позвоночных (включая человеческую линию) недалеко от происхождения костистых рыб , а еще одна - у стволовой линии костистых рыб. [8] Имеющиеся данные свидетельствуют о том, что пекарские дрожжи ( Saccharomyces cerevisiae ), имеющие компактный геном, в ходе своей эволюционной истории претерпели полиплоидизацию.
Термин «мезополиплоид» иногда используется для обозначения видов, которые претерпели события размножения всего генома (дупликация всего генома, триплификация всего генома и т. д.) в более поздней истории, например, за последние 17 миллионов лет. [9]
Эукариоты
[ редактировать ]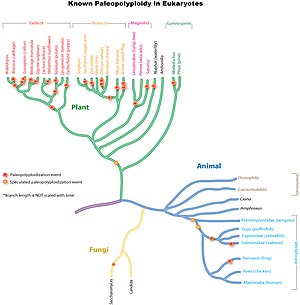
Дупликации древних геномов широко распространены во всех эукариотических линиях, особенно у растений. Исследования показывают, что общий предок Poaceae , семейства трав, в которое входят важные виды сельскохозяйственных культур, такие как кукуруза, рис, пшеница и сахарный тростник, имел общую дупликацию генома около 70 миллионов лет назад . [10] В более древних линиях однодольных произошел один или, вероятно, несколько циклов дополнительных дупликаций всего генома, которые, однако, не были присущи предковым эвдикотам . [11] Дальнейшие независимые, более поздние дупликации всего генома произошли в линиях, ведущих к кукурузе, сахарному тростнику и пшенице, но не к рису, сорго или просо. [ нужна ссылка ]
Предполагается, что событие полиплоидии, произошедшее 160 миллионов лет назад, привело к созданию наследственной линии, которая привела к появлению всех современных цветковых растений. [12] Это событие палеополиплоидии было изучено путем секвенирования генома древнего цветкового растения Amborella trichopoda . [13]
Сердцевинные эвдикоты также имели общую трипликацию всего генома (палеогексаплоидию), которая, по оценкам, произошла после расхождения однодольных и эвдикотов , но до расхождения розидов и астерид . [14] [15] [16] У многих видов эвдикотов произошли дополнительные дупликации или трипликации всего генома. Например, модельное растение Arabidopsis thaliana , первое растение, у которого секвенирован весь геном, претерпело как минимум два дополнительных раунда дупликации всего генома, поскольку дупликация была общей для основных эвдикотов. [3] произошло до расхождения линий Arabidopsis и Brassica , примерно миллионов лет 20–45 Самое последнее событие назад . Другие примеры включают секвенированные эвдикотовые геномы яблок, сои, томатов, хлопка и т. д. [ нужна ссылка ]
По сравнению с растениями палеополиплоидия в животном мире встречается гораздо реже. Идентифицирован преимущественно у земноводных и костистых рыб. Хотя некоторые исследования предполагают, что одна или несколько общих дупликаций генома присущи всем позвоночным животным (включая человека), доказательства не так убедительны, как в других случаях, поскольку дупликации, если они и существуют, произошли очень давно (около 400-500 млн лет назад). до менее 200 млн лет у растений), и этот вопрос до сих пор дискутируется. Идея о том, что позвоночные имеют общую дупликацию всего генома, известна как гипотеза 2R . Многих исследователей интересует причина, по которой в линиях животных, особенно млекопитающих, дупликаций целого генома намного меньше, чем в линиях растений. [ нужна ссылка ]
Хорошо подтвержденная палеополиплоидия была обнаружена у хлебопекарных дрожжей ( Saccharomyces cerevisiae ), несмотря на их небольшой и компактный геном (~ 13 МБП), после расхождения с Kluyveromyces Lactis и K. marxianus . [17] Благодаря оптимизации генома дрожжи потеряли 90% дублированного генома за время эволюции и теперь признаны диплоидным организмом. [ нужна ссылка ]
Метод обнаружения
[ редактировать ]Дуплицированные гены можно идентифицировать по гомологии последовательностей на уровне ДНК или белка. Палеополиплоидию можно определить как массовое дупликацию генов за один раз с помощью молекулярных часов . Чтобы отличить дупликацию всего генома от совокупности (более распространенных) событий дупликации одного гена , часто применяются следующие правила:

- Дуплицированные гены расположены в больших дуплицированных блоках. Дупликация одного гена является случайным процессом и имеет тенденцию к тому, что дублированные гены разбросаны по всему геному.
- Дублированные блоки не перекрываются, поскольку они были созданы одновременно. Сегментарная дупликация внутри генома может соответствовать первому правилу; но множественные независимые сегментные дупликации могут перекрывать друг друга.
Теоретически два дублированных гена должны иметь одинаковый «возраст»; то есть расхождение последовательности должно быть равным между двумя генами, дуплицированными палеополиплоидией ( гомеологами ). Синонимичная скорость замещения, Ks , часто используется в качестве молекулярных часов для определения времени дупликации гена. Таким образом, палеополиплоидия определяется как «пик» на графике зависимости числа дубликатов от Ks (показано справа).
Однако использование графиков Ks для идентификации и документирования древних полиплоидных событий может быть проблематичным, поскольку этот метод не позволяет выявить дупликации генома, за которыми последовало массовое удаление генов и уточнение генома. Другие подходы к смешанным моделям, сочетающие графики Ks с другими методами, разрабатываются для лучшего понимания палеополиплоидии. [18]
События дупликации, которые произошли давным-давно в истории различных эволюционных линий, могут быть трудно обнаружить из-за последующей диплоидизации (например, полиплоид с течением времени начинает вести себя цитогенетически как диплоид), поскольку мутации и трансляции генов постепенно создают по одной копии каждого из них. хромосома в отличие от своего аналога. Обычно это приводит к низкой достоверности идентификации очень древней палеополиплоидии.
Эволюционное значение
[ редактировать ]События палеополиплоидизации приводят к массивным клеточным изменениям, включая удвоение генетического материала, изменения в экспрессии генов и увеличение размера клеток. Потеря генов во время диплоидизации не является полностью случайной, а тщательно отобранной. Гены из больших генных семейств дублируются. С другой стороны, отдельные гены не дублируются. [ нужны разъяснения ] В целом палеополиплоидия может оказывать как краткосрочное, так и долгосрочное эволюционное воздействие на приспособленность организма в естественной среде. [ нужна ссылка ]
Усиленная фенотипическая эволюция
[ редактировать ]Полное дублирование генома может увеличить скорость и эффективность приобретения организмами новых биологических признаков. Однако одна проверка этой гипотезы, в ходе которой сравнивались темпы эволюции инноваций у ранних костистых рыб (с дублированными геномами) и ранних голостовых рыб (без дублированных геномов), обнаружила небольшую разницу между ними. [8]
Разнообразие геномов
[ редактировать ]Удвоение генома обеспечило организм избыточными аллелями, которые могут свободно развиваться при небольшом давлении отбора. Дуплицированные гены могут подвергаться неофункционализации или субфункционализации , что может помочь организму адаптироваться к новой среде или пережить различные стрессовые условия. [ нужна ссылка ]
Гибридная сила
[ редактировать ]Полиплоиды часто имеют более крупные клетки и даже более крупные органы. Многие важные сельскохозяйственные культуры, включая пшеницу, кукурузу и хлопок , являются палеополиплоидами, которые были отобраны для одомашнивания древними народами. [ нужна ссылка ]
Видообразование
[ редактировать ]Было высказано предположение, что многие события полиплоидизации привели к созданию новых видов за счет приобретения адаптивных признаков или из-за половой несовместимости со своими диплоидными собратьями. Примером может служить недавнее видообразование аллополиплоида Spartina — S. anglica ; полиплоидное растение настолько успешно, что оно занесено в список инвазивных видов . во многих регионах [19]
Аллополиплоидия и аутополиплоидия
[ редактировать ]Выделяют два основных отдела полиплоидии : аллополиплоидия и аутополиплоидия. Аллополиплоиды возникают в результате гибридизации двух родственных видов, а автополиплоиды возникают в результате дупликации генома вида в результате гибридизации двух родственных родителей. [20] или соматическое удвоение в репродуктивной ткани родителя. Считается, что аллополиплоидные виды гораздо более распространены в природе. [20] возможно, потому, что аллополиплоиды наследуют разные геномы, что приводит к повышенной гетерозиготности и, следовательно, к более высокой приспособленности. Эти различные геномы приводят к повышенной вероятности крупных геномных реорганизаций. [20] [21] что может быть как вредным, так и полезным. Однако аутополиплоидия обычно считается нейтральным процессом. [21] хотя была выдвинута гипотеза, что автополиплоидия может служить полезным механизмом для стимулирования видообразования и, следовательно, способствовать способности организма быстро колонизировать новые среды обитания, не подвергаясь трудоемкому и дорогостоящему периоду геномной реорганизации, который испытывают аллополиплоидные виды. Одним из распространенных источников аутополиплоидии растений являются «идеальные цветы», способные к самоопылению или «самопылению». Это, наряду с ошибками в мейозе , которые приводят к анеуплоидии , может создать среду, в которой аутополиплоидия весьма вероятна. Этот факт можно использовать в лабораторных условиях, используя колхицин для ингибирования сегрегации хромосом во время мейоза и создавая синтетические автополиплоидные растения. [ нужна ссылка ]
После событий полиплоидии существует несколько возможных судеб дуплицированных генов ; обе копии могут сохраниться как функциональные гены, изменение функции гена может произойти в одной или обеих копиях, замалчивание генов может маскировать одну или обе копии или может произойти полная потеря гена. [20] [22] События полиплоидии приведут к более высокому уровню гетерозиготности и со временем могут привести к увеличению общего количества функциональных генов в геноме. По прошествии времени после события дупликации генома многие гены изменят функцию в результате либо изменения функции дублирующихся генов как для алло-, так и для аутополиплоидных видов, либо в результате изменений в экспрессии генов, вызванных геномными перестройками, вызванными дупликацией генома у аллополиплоидов. . Когда обе копии гена сохраняются и, таким образом, количество копий удваивается, существует вероятность того, что произойдет пропорциональное увеличение экспрессии этого гена, что приведет к мРНК образованию вдвое большего количества транскриптов . Существует также вероятность того, что транскрипция дуплицированного гена будет подавляться, что приведет к менее чем двукратному увеличению транскрипции этого гена, или что событие дупликации приведет к более чем двукратному увеличению транскрипции. [23] У одного вида, Glycine dolichocarpa (близкого родственника соевых бобов , Glycine max ), было замечено, что после дупликации генома примерно 500 000 лет назад произошло увеличение транскрипции в 1,4 раза, что указывает на пропорциональное снижение в транскрипции относительно количества копий гена после события дупликации. [23]
Позвоночные животные как палеополиплоиды
[ редактировать ]Гипотеза палеополиплоидии позвоночных возникла еще в 1970-х годах и была предложена биологом Сусуму Оно . Он рассуждал, что геном позвоночных не мог бы достичь такой сложности без крупномасштабных дупликаций целого генома. гипотеза «двух раундов дупликации генома» ( гипотеза 2R ), которая приобрела популярность, особенно среди биологов развития. Появилась [ нужна ссылка ]
Некоторые исследователи поставили под сомнение гипотезу 2R, поскольку она предсказывает, что геномы позвоночных должны иметь соотношение генов 4:1 по сравнению с геномами беспозвоночных, и это не подтверждается результатами 48 проектов геномов позвоночных, доступных в середине 2011 года. Например, согласно подсчетам, проведенным в июне 2011 года в центрах анализа генома UCSC и Ensembl, геном человека состоит из примерно 21 000 генов, кодирующих белки. [ нужна ссылка ] в то время как средний размер генома беспозвоночных составляет около 15 000 генов. Последовательность генома амфиоксуса подтвердила гипотезу о двух раундах дупликации всего генома, за которой последовала потеря дубликатов копий большинства генов. [24] Дополнительные аргументы против 2R были основаны на отсутствии топологии дерева (AB) (CD) среди четырех членов семейства генов у позвоночных. Однако если бы две дупликации генома произошли близко друг к другу, мы бы не ожидали обнаружить такую топологию. [25] Недавнее исследование создало генетическую карту морской миноги , которая предоставила убедительную поддержку гипотезе о том, что в базальной линии позвоночных произошла одна дупликация всего генома, которой предшествовали и следовали несколько эволюционно независимых сегментных дупликаций, которые произошли в ходе эволюции хордовых. [26]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Гарсмер, Оливье; Шнабл, Джеймс С.; Алмейда, Ана; Журда, Кирилл; Д'Онт, Анжелика; Фрилинг, Майкл (1 февраля 2014 г.). «Два эволюционно различных класса палеополиплоидии» . Оксфордский академический . Проверено 28 июня 2024 г.
- ^ Келлис М., Биррен Б.В., Ландер Э.С. (апрель 2004 г.). «Доказательство и эволюционный анализ дублирования древнего генома дрожжей Saccharomyces cerevisiae». Природа . 428 (6983): 617–24. Бибкод : 2004Natur.428..617K . дои : 10.1038/nature02424 . ПМИД 15004568 . S2CID 4422074 .
- ^ Jump up to: а б Бауэрс Дж. Э., Чепмен Б. А., Ронг Дж., Патерсон А. Х. (март 2003 г.). «Разгадка эволюции генома покрытосеменных путем филогенетического анализа событий хромосомной дупликации». Природа . 422 (6930): 433–8. Бибкод : 2003Natur.422..433B . дои : 10.1038/nature01521 . ПМИД 12660784 . S2CID 4423658 .
- ^ Смит Дж.Дж., Кураку С., Холт С., Саука-Шпенглер Т., Цзян Н., Кэмпбелл М.С. и др. (апрель 2013 г.). «Секвенирование генома морской миноги (Petromyzon marinus) дает представление об эволюции позвоночных» . Природная генетика . 45 (4): 415–21, 421e1-2. дои : 10.1038/ng.2568 . ПМЦ 3709584 . ПМИД 23435085 .
- ^ Вулф К.Х. (май 2001 г.). «Вчерашние полиплоиды и тайна диплоидизации». Обзоры природы. Генетика . 2 (5): 333–41. дои : 10.1038/35072009 . ПМИД 11331899 . S2CID 20796914 .
- ^ Блан Дж., Вулф К.Х. (июль 2004 г.). «Широко распространенная палеополиплоидия у модельных видов растений, выведенная на основе возрастного распределения дублирующих генов» . Растительная клетка . 16 (7): 1667–78. дои : 10.1105/tpc.021345 . ПМК 514152 . ПМИД 15208399 .
- ^ Блан Дж., Вулф К.Х. (июль 2004 г.). «Функциональная дивергенция дуплицированных генов, образовавшихся в результате полиплоидии в ходе эволюции Arabidopsis» . Растительная клетка . 16 (7): 1679–91. дои : 10.1105/tpc.021410 . ПМК 514153 . ПМИД 15208398 .
- ^ Jump up to: а б Кларк Дж.Т., Ллойд Г.Т., Фридман М. (октябрь 2016 г.). «Мало доказательств усиленной фенотипической эволюции ранних костистых рыб по сравнению с их живой сестринской группой ископаемых» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (41): 11531–11536. Бибкод : 2016PNAS..11311531C . дои : 10.1073/pnas.1607237113 . ПМК 5068283 . ПМИД 27671652 .
- ^ Ван X , Ван Х, Ван Дж, Сунь Р, Ву Дж, Лю С и др. (август 2011 г.). «Геном мезополиплоидного вида сельскохозяйственных культур Brassica rapa» . Природная генетика . 43 (10): 1035–9. дои : 10.1038/ng.919 . ПМИД 21873998 . S2CID 205358099 .
- ^ Патерсон А.Х., Бауэрс Дж.Э., Чепмен Б.А. (июнь 2004 г.). «Древняя полиплоидизация, предшествовавшая дивергенции злаков, и ее последствия для сравнительной геномики» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (26): 9903–8. Бибкод : 2004PNAS..101.9903P . дои : 10.1073/pnas.0307901101 . ПМК 470771 . ПМИД 15161969 .
- ^ Тан Х, Бауэрс Дж. Э., Ван X, Патерсон А. Х. (январь 2010 г.). «Сравнение генома покрытосеменных растений выявляет раннюю полиплоидию в линии однодольных» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (1): 472–7. Бибкод : 2010PNAS..107..472T . дои : 10.1073/pnas.0908007107 . ПМК 2806719 . ПМИД 19966307 .
- ^ Каллауэй Э (декабрь 2013 г.). «Геном кустарника раскрывает секреты силы цветов» . Природа . дои : 10.1038/nature.2013.14426 . S2CID 88293665 .
- ^ Адамс К. (декабрь 2013 г.). «Геномика. Геномные ключи к разгадке предков цветковых растений». Наука . 342 (6165): 1456–7. Бибкод : 2013Sci...342.1456A . дои : 10.1126/science.1248709 . ПМИД 24357306 . S2CID 206553839 .
- ^ Тан Х, Ван X, Бауэрс Дж. Э., Минг Р., Алам М., Патерсон А. Х. (декабрь 2008 г.). «Раскрытие древней гексаплоидии с помощью множественных карт генов покрытосеменных» . Геномные исследования . 18 (12): 1944–54. дои : 10.1101/гр.080978.108 . ПМЦ 2593578 . ПМИД 18832442 .
- ^ Жайон О, Ори Дж. М., Ноэль Б., Поликрити А., Клепет С., Касагранде А. и др. (сентябрь 2007 г.). «Последовательность генома виноградной лозы предполагает предковую гексаплоидизацию основных типов покрытосеменных» . Природа . 449 (7161): 463–7. Бибкод : 2007Natur.449..463J . дои : 10.1038/nature06148 . hdl : 11577/2430527 . ПМИД 17721507 .
- ^ Тан Х., Бауэрс Дж.Э., Ван Х, Минг Р., Алам М., Патерсон А.Х. (апрель 2008 г.). «Синтения и коллинеарность в геномах растений». Наука . 320 (5875): 486–8. Бибкод : 2008Sci...320..486T . дои : 10.1126/science.1153917 . ПМИД 18436778 . S2CID 206510918 .
- ^ Вонг С., Батлер Г., Вулф К.Х. (июль 2002 г.). «Эволюция порядка генов и палеополиплоидия у гемиаскомицетных дрожжей» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (14): 9272–7. Бибкод : 2002PNAS...99.9272W . дои : 10.1073/pnas.142101099 . ПМЦ 123130 . ПМИД 12093907 .
- ^ Тайли Г.П., Баркер М.С., Берли Дж.Г. (ноябрь 2018 г.). «Оценка эффективности Ks-графиков для обнаружения древних дупликаций целого генома» . Геномная биология и эволюция . 10 (11): 2882–2898. дои : 10.1093/gbe/evy200 . ПМК 6225891 . ПМИД 30239709 .
- ^ te Beest M, Le Roux JJ, Richardson DM, Brysting AK, Suda J, Kubesová M, Pysek P (январь 2012 г.). «Чем больше, тем лучше? Роль полиплоидии в облегчении инвазий растений» . Анналы ботаники . 109 (1): 19–45. дои : 10.1093/aob/mcr277 . ПМЦ 3241594 . ПМИД 22040744 .
- ^ Jump up to: а б с д Солтис PS , Солтис DE (июнь 2000 г.). «Роль генетических и геномных признаков в успехе полиплоидов» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (13): 7051–7. Бибкод : 2000PNAS...97.7051S . дои : 10.1073/pnas.97.13.7051 . ПМК 34383 . ПМИД 10860970 .
- ^ Jump up to: а б Паризод С., Холдереггер Р., Брохманн С. (апрель 2010 г.). «Эволюционные последствия аутополиплоидии». Новый фитолог . 186 (1): 5–17. дои : 10.1111/j.1469-8137.2009.03142.x . ПМИД 20070540 .
- ^ Вендел Дж. Ф. (2000). «Эволюция генома полиплоидов». Молекулярная эволюция растений . Том. 42. С. 225–249. дои : 10.1007/978-94-011-4221-2_12 . ISBN 978-94-010-5833-9 . ПМИД 10688139 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Jump up to: а б Коут Дж. Э., Дойл Дж. Дж. (2010). «Количественная оценка размера всего транскриптома - необходимое условие для понимания эволюции транскриптома у разных видов: пример аллополиплоида растения» . Геномная биология и эволюция . 2 : 534–46. дои : 10.1093/gbe/evq038 . ПМЦ 2997557 . ПМИД 20671102 .
- ^ Патнэм Н.Х., Баттс Т., Ферье Д.Э., Ферлонг Р.Ф., Хеллстен Ю., Кавашима Т. и др. (июнь 2008 г.). «Геном амфиоксуса и эволюция кариотипа хордовых» . Природа . 453 (7198): 1064–71. Бибкод : 2008Natur.453.1064P . дои : 10.1038/nature06967 . ПМИД 18563158 .
- ^ Ферлонг РФ, Голландия PW (апрель 2002 г.). «Были ли позвоночные октоплоидными?» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 357 (1420): 531–44. дои : 10.1098/rstb.2001.1035 . ПМК 1692965 . ПМИД 12028790 .
- ^ Смит Джей Джей, Кейнат MC (август 2015 г.). «Мейотическая карта морской миноги улучшает разрешение дупликаций генома древних позвоночных» . Геномные исследования . 25 (8): 1081–90. дои : 10.1101/гр.184135.114 . ПМК 4509993 . ПМИД 26048246 .
Дальнейшее чтение
[ редактировать ]- Адамс К.Л., Вендел Дж.Ф. (апрель 2005 г.). «Полиплоидия и эволюция генома растений». Современное мнение в области биологии растений . 8 (2): 135–41. дои : 10.1016/j.pbi.2005.01.001 . ПМИД 15752992 .
- Куи Л., Уолл П.К., Либенс-Мак Дж.Х., Линдсей Б.Г., Солтис Д.Е., Дойл Дж.Дж. и др. (июнь 2006 г.). «Широко распространенные дупликации генома на протяжении всей истории цветковых растений» . Геномные исследования . 16 (6): 738–49. дои : 10.1101/гр.4825606 . ПМЦ 1479859 . ПМИД 16702410 .
- Комай Л. (ноябрь 2005 г.). «Преимущества и недостатки полиплоида». Обзоры природы. Генетика . 6 (11): 836–46. дои : 10.1038/nrg1711 . ПМИД 16304599 . S2CID 3329282 .
- Экардт Н.А. (июль 2004 г.). «Два генома лучше, чем один: широко распространенная палеополиплоидия у растений и эволюционные эффекты» . Растительная клетка . 16 (7): 1647–1649. дои : 10.1105/tpc.160710 . ПМК 514149 . ПМИД 15272471 .
- Отто С.П., Уиттон Дж (2000). «Заболеваемость и эволюция полиплоидов». Ежегодный обзор генетики . 34 (1): 401–437. CiteSeerX 10.1.1.323.1059 . дои : 10.1146/annurev.genet.34.1.401 . ПМИД 11092833 .
- Макаловский В. (май 2001 г.). «Мы полиплоиды? Краткая история одной гипотезы» . Геномные исследования . 11 (5): 667–70. дои : 10.1101/гр.188801 . ПМИД 11337465 .
- Келлис М., Биррен Б.В., Ландер Э.С. (апрель 2004 г.). «Доказательство и эволюционный анализ дублирования древнего генома дрожжей Saccharomyces cerevisiae». Природа . 428 (6983): 617–24. Бибкод : 2004Natur.428..617K . дои : 10.1038/nature02424 . ПМИД 15004568 . S2CID 4422074 .