Экологическое видообразование

Экологическое видообразование — это форма видообразования , возникающая в результате репродуктивной изоляции , возникающая из-за экологического фактора, который уменьшает или устраняет поток генов между двумя популяциями вида. Экологические факторы могут включать изменения в условиях окружающей среды, в которых находится вид, например, изменения в поведении, связанные с хищничеством , избеганием хищников, привлечением опылителей и добычей пищи ; а также изменения в выборе партнера из-за полового отбора или систем коммуникации. Экологическая репродуктивная изоляция в условиях дивергентного естественного отбора приводит к образованию новых видов. Это было задокументировано во многих случаях в природе и было основным направлением исследований видообразования в течение последних нескольких десятилетий. [1] : 179
Экологическое видообразование определялось по-разному, чтобы отличить его от неэкологических форм видообразования. [2] Биолог-эволюционист Дольф Шлютер определяет это как «эволюцию репродуктивной изоляции между популяциями или подгруппами одной популяции путем адаптации к различным средам или экологическим нишам». [3] в то время как другие полагают, что движущей силой является естественный отбор. [4] [5] [6] Ключевое различие между экологическим видообразованием и другими видами видообразования заключается в том, что оно запускается дивергентным естественным отбором среди различных сред обитания, в отличие от других видов процессов видообразования, таких как случайный генетический дрейф , фиксация несовместимых мутаций в популяциях, испытывающих аналогичное селективное давление , или различные формы полового отбора, не включающие отбор по экологически значимым признакам. Экологическое видообразование может происходить либо в аллопатрии , симпатрии или парапатрии — единственное требование состоит в том, чтобы видообразование происходило в результате адаптации к различным экологическим или микроэкологическим условиям. [6]
Экологическое видообразование может происходить презиготно (барьеры размножения, возникающие до образования зиготы ) или постзиготно (барьеры размножения, возникающие после образования зиготы). Примеры презиготической изоляции включают изоляцию среды обитания , изоляцию с помощью систем опылителей- опыления и временную изоляцию. Примеры постзиготической изоляции включают генетическую несовместимость гибридов , гибридов с низкой приспособленностью и половой отбор против гибридов.
Существуют некоторые дебаты по поводу определения того, является ли событие видообразования экологическим или неэкологическим. «Повсеместный эффект отбора предполагает, что адаптивная эволюция и видообразование неразделимы, что ставит под сомнение то, что видообразование когда-либо неэкологично». [2] Однако существуют многочисленные примеры близкородственных, экологически сходных видов (например, наземные улитки Albinaria на островах Средиземноморья , [7] Батрахосепс Саламандры из Калифорнии , [8] и некоторые сверчки [9] и стрекозы [10] ), что соответствует возможности неэкологического видообразования. [8] [11]
отбора дивергентного Экологические причины
Дивергентный отбор является ключом к возникновению экологического видообразования. Выявлены три экологические причины дивергентного отбора: различия в условиях окружающей среды, экологические взаимодействия и половой отбор. Причины приведены в следующем списке. [12] [13] [4]

Эксперимент 1: событие видообразования, которое, по прогнозам, произошло из-за экологически обоснованного фактора дивергенции, приведшего к появлению двух новых видов (1a). В результате эксперимента получаются жизнеспособные и плодовитые гибридные потомки, которые помещаются в изолированные условия, соответствующие их родительской среде (1b). Эксперимент предсказывает, что «репродуктивная изоляция должна затем развиваться во взаимосвязи с окружающей средой, создавая [увеличиваясь] между популяциями в разных средах и отсутствуя между лабораторными и естественными популяциями из схожих сред». [4]
Эксперимент 2: происходит перипатрическое видообразование между материковым видом и изолированной эндемической популяцией (2a). Лабораторная обстановка повторяет условия окружающей среды материка, которые, как считается, способствовали видообразованию, и в нее помещена материковая популяция. Эксперимент предсказывает, что трансплантат покажет признаки изоляции, соответствующие островным эндемикам (2b). [4]
- Различия в условиях среды как предпосылка видообразования, несомненно, являются наиболее изученными. [4] Хищничество , наличие ресурсов (обилие пищи), климатические условия и структура среды обитания — вот лишь некоторые из примеров, которые могут различаться и вызывать дивергентный отбор. [14] Несмотря на то, что это один из наиболее изученных факторов экологического видообразования, многие аспекты еще менее понятны, например, насколько распространен этот процесс в природе. [4] а также происхождение барьеров постзиготической изоляции (в отличие от гораздо более легко обнаруживаемых презиготических барьеров). [1] : 181 Лабораторные эксперименты, включающие различия в одной среде, ограничены и часто не позволяют отследить черты, проявляющиеся в изоляции. Исследования в природе были сосредоточены на различных факторах окружающей среды, таких как дивергентный отбор, вызванный хищничеством; однако мало что было изучено в отношении патогенов и паразитов . [4]
- Экологические взаимодействия могут привести к различному отбору между симпатическими популяциями. [4] Примерами таких взаимодействий могут быть внутривидовая (между одними и теми же видами) и межвидовая (между разными видами) конкуренция. [15] или такие отношения, как экологическое содействие . [16] [17] Межвидовая конкуренция, в частности, поддерживается экспериментами; [14] однако неизвестно, может ли это привести к репродуктивной изоляции, несмотря на расходящийся отбор. [4] Подкрепление (усиление изоляции путем отбора, благоприятствующего спариванию членов их собственных популяций из-за снижения приспособленности гибридов) считается формой экологического видообразования или участвует в нем. [4] [18] Однако существуют споры о том, как определить конечные причины, поскольку подкрепление может завершить процесс видообразования независимо от того, как он возник. [19] Кроме того, смещение персонажа может иметь тот же эффект. [4]
- Половой отбор может играть роль в экологическом видообразовании, поскольку признание партнеров играет центральную роль в репродуктивной изоляции. [20] — то есть, если вид не может распознать своих потенциальных партнеров, поток генов приостанавливается. Несмотря на свою роль, только два типа полового отбора могут быть вовлечены в экологическое видообразование: пространственная изменчивость вторичных половых признаков (половые признаки, возникающие именно в период половой зрелости) [21] или системы связи и спаривания . [22] Это ограничение основано на том факте, что они создают различные среды, в которых может действовать отбор. [4] Например, изоляция будет возрастать между двумя популяциями, где существует несоответствие между сигналами (например, оперением самца) и предпочтениями (например, сексуальными предпочтениями самки). [22] Эта закономерность была обнаружена у колюшек. [23]
| Тип репродуктивной изоляции | Презиготический или постзиготический | Экологическая причина выбора | |||
|---|---|---|---|---|---|
| Различные среды | Экологические взаимодействия | Половой отбор | Армирование | ||
| среда обитания | Предварительно | ✓ | ✓ | ✓ | |
| Половой/Опылитель | Предварительно | ✓ | ✓ | ✓ | ✓ |
| Временной | Предварительно | ✓ | ✓ | ✓ | |
| Отбор против мигрантов | Предварительно | ✓ | ✓ | ✓ | |
| После спаривания | Предварительно | ✓ | ✓ | ✓ | ✓ |
| Отбор против гибридов | Почта | ✓ | ✓ | ✓ | ✓ |
| Экологически независимый | Почта | ✓ | ✓ | ✓ | ✓ |
| Экологически зависимый | Почта | ✓ | ✓ | ||
Виды репродуктивной изоляции [ править ]
среды Изоляция обитания

Популяции вида могут стать пространственно изолированными из-за предпочтения отдельных местообитаний. [4] Разделение снижает вероятность спаривания между двумя популяциями, подавляя поток генов и способствуя презиготической изоляции, что приводит к полному видообразованию. [4] Изоляция среды обитания не эквивалентна географическому барьеру, подобному барьеру аллопатрического видообразования . [1] : 182 Вместо этого он основан на генетических различиях, когда один вид не может использовать другую среду обитания из-за преимуществ приспособленности, недостатков приспособленности или конкуренции за ресурсы. [1] : 182
Джерри Койн и Х. Аллен Орр постулируют две различные формы изоляции среды обитания: микропространственную изоляцию среды обитания (когда спаривание между двумя видами сокращается из-за предпочтений или адаптации к экологически различающимся территориям, несмотря на то, что они занимают одну и ту же обобщенную территорию) и макропространственную изоляцию среды обитания (определяемую полностью аллопатрические среды обитания, которые ингибируют поток генов.) [1] : 182–3 Идентификация обеих форм изоляции местообитаний в природе затруднена из-за географических эффектов. Для измерения микропространственной изоляции необходимо учитывать несколько факторов: [1] : 184
- пространственное разделение представителей разных видов больше, чем у представителей одного и того же вида
- во время одновременных периодов размножения пространственное разделение уменьшает поток генов
- уменьшение потока генов является прямым результатом уменьшения спаривания
- генетические различия соответствуют пространственному разделению
Аллопатрическое распределение создает несколько проблем для обнаружения истинной изоляции среды обитания, поскольку разные среды обитания двух аллопатрически изолированных видов не подразумевают экологически обусловленное видообразование. Альтернативные объяснения могут объяснить закономерности: [1] : 185
- Видовые различия могут быть вызваны географической изоляцией
- виды могут занимать или не занимать разные среды обитания, если они существовали в симпатии
- в случае схожих местообитаний в аллопатрии виды могут быть адаптированы к неизвестным экологическим факторам.
- если бы виды существовали в симпатрии, конкуренция могла бы привести к сегрегации среды обитания, которая была бы необнаружима в аллопатрии.
Эти проблемы (как с микро-, так и с макропространственной изоляцией) можно решить с помощью полевых или лабораторных экспериментов, таких как трансплантация особей в противоположные места обитания. [1] : 185 (хотя это может оказаться затруднительным, если особи не полностью непригодны для навязанной среды обитания). [1] : 186 Изоляцию среды обитания можно измерить для пары видов ( и ) в период размножения путем:



Здесь, - это доля наблюдаемых встреч между спариваниями с участием партнеров разных видов. это доля от общего количества особей вида . это доля от общего количества особей вида . Ожидаемая доля спариваний между разными видами, если спаривание носит случайный характер, обозначается как . Статистика указывает на отсутствие спаривающихся встреч разных видов, где указывает на случайное спаривание разных видов.






География [ править ]
Экологическое видообразование, вызванное изоляцией среды обитания, может происходить в любом географическом смысле: аллопатрически, парапатрически или симпатрически. [4] Видообразование, возникающее в результате изоляции среды обитания в аллопатрии (и парапатрии), является простым в том смысле, что уменьшенный поток генов между двумя популяциями приобретает адаптации, соответствующие экологическим условиям их среды обитания. Адаптации подкрепляются отбором и во многих случаях, например, у животных, подкрепляются поведенческими предпочтениями (например, у птиц, которые предпочитают определенные вокализации). [1] : 189 в зависимости от хозяина. Классическим примером изоляции среды обитания, происходящей при аллопатрии, является ковидообразование [1] : 189 например, у карманных сусликов и их хозяина, жующего вшей [24] или в отношениях инжирной осы и инжирного дерева и отношениях юкки и юкки- мотыльки - примеры экологического видообразования, вызванного изоляцией опылителей. [1] : 189 В случае симпатии сценарий более сложен, поскольку поток генов может быть недостаточно сокращен, чтобы обеспечить видообразование. Считается, что этот процесс может стимулироваться отбором в пользу дивергенции ниш . Кроме того, если симпатрия возникает в результате вторичного контакта двух ранее разделенных популяций, процесс подкрепления, отбор против непригодных гибридов между двумя популяциями, может привести к их полному видообразованию. Конкуренция за ресурсы также может сыграть свою роль. [1] : 191

Изоляция среды обитания является серьезным препятствием для потока генов, о чем свидетельствует общее наблюдение, что растения и животные часто пространственно разделены в зависимости от их адаптации. [1] : 183 Чтобы понять природу видообразования, вызванного изоляцией среды обитания, были проведены многочисленные полевые исследования, эксперименты по трансплантации и удалению, а также лабораторные исследования. [1] : 186–188 Horkelia fusca , например, растет на калифорнийских склонах и лугах на высоте более 4500 футов, где ее ближайшие родственники H.californica и H.cuneata растут ниже 3200 футов в прибрежных средах обитания. Когда виды пересаживаются в альтернативные места обитания, их жизнеспособность снижается, что указывает на то, что поток генов между популяциями маловероятен. [25] Аналогичные закономерности были обнаружены у Artemisia tridentata tridentata и A. tridentata subsp. vaseyana в штате Юта, где существуют гибридные зоны между высотными популяциями, а эксперименты по трансплантации снижают приспособленность подвида. [26]
Видообразование за счет изоляции среды обитания также изучалось у змеевидных минирующих мух. [27] жуки-божьи коровки ( Epilachna ), [28] Золотарник галловый летает , [29] Раголетис помонелла , [30] [31] листоеды , [32] и гороховая тля . [33]
Сексуальная изоляция [ править ]
Экологическое видообразование из-за половой изоляции является результатом различных условий окружающей среды, которые со временем изменяют системы коммуникации или модели выбора партнера. [4] Примеров в природе предостаточно. [4] Прибрежный вид улиток Littorina saxatilis оказался в центре внимания исследований. [4] поскольку два экотипа, обитающие на разных уровнях побережья, демонстрируют репродуктивную изоляцию в результате выбора партнера с учетом различий в размерах тела экотипа. [34] Как морские, так и пресноводные рыбы-колюшки продемонстрировали убедительные доказательства того, что они образовались таким образом. [35] [36] [37] [38] Доказательства также обнаружены у Neochlamisus bebbianae . листоедов [32] Timema cristinae , Насекомые-трости [39] [40] и у видов бабочек Heliconius melpomene и H. cydno, которые, как полагают, недавно разошлись из-за усиления ассортативного спаривания там, где популяции видов встречаются в симпатии. [41]
Изоляция опылителей [ править ]
Покрытосеменные (цветковые растения) требуют той или иной формы опыления , многие из которых требуют, чтобы другое животное переносило пыльцу с одного цветка на другой. [42] Биотические методы опыления требуют наличия опылителей, таких как насекомые (например, пчелы, бабочки, мотыльки, осы, жуки и другие беспозвоночные). [42] птицы, летучие мыши, [43] и другие виды позвоночных. Из-за этой эволюционной связи между опылителями и растениями, производящими пыльцу, растения и животные становятся взаимозависимыми друг от друга: опылитель получает пищу в виде нектара, а цветок приобретает способность размножать свои гены.
В случае, если животное использует другой источник опыления, растения могут оказаться репродуктивно изолированными. [1] : 193 Изоляция опылителей — это особая форма половой изоляции. [4] Ботаник Верн Грант различал два типа изоляции опылителей: механическую изоляцию и этологическую изоляцию. [1] : 193 [44] : 75
изоляция Механическая опылителей

Механическая изоляция возникает в результате анатомических различий цветка или опылителя, препятствующих опылению. [44] Например, у пчелы Eulaema cingulata пыльца Catasetum discolor и C. saccatum прикрепляется к разным частям тела (соответственно вентрально и дорсально). [1] : 194 [45] Другой пример — растения «голова слона» и «голова маленького слона» . Известно, что они не гибридизуются, несмотря на то, что растут в одном и том же регионе и опыляются одними и теми же видами пчел. Пыльца прикрепляется к различным частям пчелы, что делает цветы изолированными. [44] Механическая изоляция также включает в себя опылителей, которые не могут опылять из-за физических недостатков. [1] : 194 нектарной шпоры Например, длина может варьироваться по размеру в зависимости от вида цветка, что приводит к опылению от разных видов чешуекрылых из-за того, что длина предотвращает контакт тела с пыльцой цветка.
опылителей Этологическая изоляция
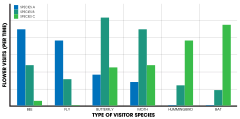
Этологическая изоляция основана на поведенческих особенностях опылителей, которые отдают предпочтение различным морфологическим характеристикам цветка либо генетически, либо в результате приобретенного поведения. Этими характеристиками могут быть общая форма и структура, цвет, тип нектара или запах цветка. [1] : 194 В некоторых случаях мутуализмы между опылителем и его хозяином развиваются , образуя почти конгруэнтные параллельные филогении . [1] : 196 То есть зависимые отношения приводят к образованию почти идентичных эволюционных деревьев, что указывает на то, что события видообразования и скорость видообразования идентичны. Примеры встречаются у инжирных ос и их хозяев , причем каждый вид инжирных ос опыляет определенный вид инжира. [46] Юкка . и юкка моль демонстрируют ту же самую картину [47]


Поразительный случай: два близкородственных цветковых растения ( Erythranthe lewisii и E. cardinalis ) развились благодаря изоляции опылителей в полной симпатии (видообразование происходит без какой-либо физической, географической изоляции). [4] E. lewisii значительно изменился по сравнению со своим родственным видом: у него появились розовые цветки, широкие лепестки, более короткие тычинки (часть растения, производящая пыльцу) и меньший объем нектара. Он полностью опыляется пчелами, практически не допуская скрещивания в природе. E. cardinalis опыляется колибри и имеет красные цветки трубчатой формы, более крупные тычинки и много нектара. Считается, что объем нектара, а также генетический компонент (замена аллелей, контролирующая изменение цвета) поддерживают изоляцию. [48] [49] Похожая картина была обнаружена у Aquilegia pubescens и A. formosa . В этой паре видов A. pubescens опыляется ястребами, а A. formosa опыляется колибри. [1] : 197 В отличие от Erythranthe , эти виды обитают в разных средах обитания, но демонстрируют гибридные формы, в которых их среды обитания перекрываются; [50] хотя они остаются отдельными видами, что позволяет предположить, что гибридные цветы могут быть менее привлекательными для своих хозяев-опылителей. [1] : 197
География [ править ]
Известно, что происходят четыре географических сценария, включающих изоляцию опылителей:
- Наиболее распространенная схема изоляции опылителей в географическом контексте предполагает, что дивергенция цветочных признаков происходит в результате географической изоляции (аллопатрически). Таким образом, популяция может столкнуться с разными опылителями, что в конечном итоге приведет к отбору благоприятных признаков для привлечения опылителей и достижения репродуктивного успеха. [1] : 198
- Другой сценарий включает начальную аллопатрическую стадию, на которой вторичный контакт происходит на различном уровне репродуктивной изоляции: высокая изоляция фактически является аллопатрическим видообразованием, тогда как низкая изоляция фактически является симпатрическим. [4] Эта «двухступенчатая» модель обозначена у трехиглых колюшек. [51] а также яблоневая личинка и ее яблочные хозяева. [52]
- Опылитель может изменить предпочтения благодаря своей собственной эволюции, приводящей к отбору в пользу признаков, которые соответствуют изменившимся предпочтениям опылителя. [1] : 198
- Существует вероятность того, что, когда две популяции оказываются географически изолированными, растение или опылитель может вымереть в одной из популяций, что приведет к отбору в пользу разных признаков. [53]
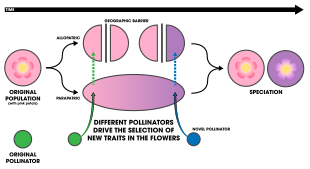
Джерри Койн и Х. Аллен Орр утверждают, что любой сценарий изоляции опылителей в аллопатрии требует, чтобы начальные стадии были обнаружены в разных популяциях. Это наблюдалось в разной степени у нескольких пар вид-опылитель. размер цветков Raphanus sativus (в данном случае дикой редьки в 32 популяциях Калифорнии) различается в зависимости от более крупных пчел-опылителей. Было обнаружено, что [54] Polemonium viscosum Было обнаружено, что цветы увеличиваются в размерах вдоль альпийского уклона в Скалистых горах Колорадо , поскольку мухи опыляют линию леса, а шмели опыляют более высокие возвышенности. [55] Аналогичная закономерность, включающая время активности бражников ( Hyles lineata ), зарегистрирована у трех подвидов Aquilegia coerulea , водосборника Скалистых гор, встречающегося на западе Соединенных Штатов. [56]
Наиболее ярким примером, по мнению Койна и Орра, являются подвиды африканских орхидей Satyrium Hallackii Hallackii и Satyrium Hallackii ocellatum . [1] : 199–200 Последний опыляется мотыльками и имеет длинные нектарные шпоры, соответствующие хоботку мотылька. В отличие от внутренней, луговой среды обитания подвида Hallackii , ocellatum обитает в прибрежных популяциях и имеет короткие отростки, которые коррелируют с его основным опылителем-пчелой-плотником. Мотыльки не могут найти подходящие места для гнезд в прибрежных местах обитания, а пчелы - в глубине суши. Эта закономерность разделяет популяции опылителей, но не разделяет популяцию орхидей, что приводит к отбору в пользу цветочных различий, которые лучше соответствуют местным опылителям. [57] Похожая закономерность была обнаружена при исследовании комплекса Disa draconis в Южной Африке. [58]
Временная изоляция ( аллохронное видообразование )
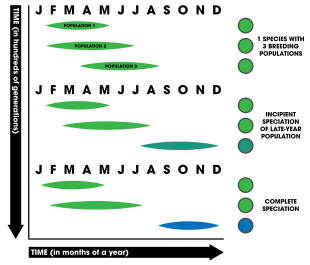
Временная изоляция основана на уменьшении потока генов между двумя популяциями из-за разного времени размножения ( фенология ). Его также называют аллохронной изоляцией, аллохронным видообразованием или аллохронией. У растений селекция по времени может включать в себя восприимчивость рыльца к принятию спермы, периоды выброса пыльцы (например, у хвойных деревьев, где шишки разносят пыльцу ветром) или общие сроки цветения. Напротив, у животных часто бывают периоды или сезоны спаривания (а у многих водных животных есть время нереста ). [1] : 202 Миграционные модели также участвуют в аллохронном видообразовании. [59] [60] [61] : 92–96
Чтобы считать, что аллохронное видообразование действительно произошло, модель предъявляет три основных требования: [62]
- Филогенетический анализ показывает, что зарождающиеся виды являются сестринскими таксонами.
- Время размножения генетически обусловлено (передается по наследству потомству).
- Источником дивергенции является явная аллохрония, а не результат подкрепления или других механизмов.
Считается, что аллохрония развивается тем легче, чем выше наследственность сроков размножения, то есть чем больше связь между генами и временем размножения, тем более вероятно, что произойдет видообразование. [63] Временная изоляция уникальна тем, что она может быть как явно симпатрической, так и негенетической; [1] : 203 однако для изоляции должны быть задействованы генетические факторы, чтобы привести к полной репродуктивной изоляции и последующему видообразованию. Известно, что видообразование посредством аллохронии происходит в трех временных рамках: ежегодно (например, периодическое появление цикад в течение десятилетий или многодесятилетнее цветение бамбука ), сезонное (организмы, которые размножаются в течение времени года, например, зимой или летом) и ежедневно (например, ежедневно время нереста кораллов). [62] В приведенном ниже списке таблиц суммирован ряд исследований, которые считаются яркими или убедительными примерами аллохронного видообразования, происходящего в природе. [62]
| Разновидность | Описание |
|---|---|
| виды Acropora . | Японские кораллы оказались репродуктивно изолированными по времени их нереста. [64] |
| Montastraea annularis , M. faveolata и M. Franksi. | Три родственных вида кораллов , которые видоизменились в зависимости от времени нереста. [65] |
| Онкоринхус нерка | Ежегодные ходы размножения нерки, происходящие в течение двух периодов года (позднего и раннего), привели к генетической изоляции зарождающихся популяций. Известно, что разведение лосося является генетическим, но специфические гены этого вида неизвестны. [66] [67] [68] |
| Тауметопея питиокампа | Кодоминирование генов связано со временем вылета личиночных стадий этого вида бабочек. Зимние и летние популяции личинок находятся в процессе видообразования. [69] [70] [71] |
| Inurois punctigera | Размножение предотвращается в районах, где температура середины зимы не подходит для этих видов бабочек. Это привело к появлению поздних и ранних популяций. [72] |
| Pemphigus populi-transversus и P. obesinymphae | Галлообразующие тли образуют галлы на разных листьях одного и того же вида деревьев-хозяев. P. populi-transversus образует галлы на листьях ранней весной, а P. obesinymphae образует их на листьях летом. Это привело к полной репродуктивной изоляции. [73] |
| Asphondylia spp. | Три вида мошек поражают стебли Larrea tridentata , A. auripila летом, A. reinosa зимой и A. foliosa весной. [74] |
| Акропора самоенсис | Популяции симпатрических видов кораллов нерестятся отдельно осенью и весной, причем нерест является наследственным, вероятно, с участием гена PaxC. [75] |
| Целлана виды. | Обитая на разной глубине в пределах нескольких сантиметров, блюда стали репродуктивно изолированными, вероятно, из-за сочетания парапатрического видообразования и сигналов нереста (например, нерест в зависимости от уровня воды. [76] |
| Гидробаты виды. | В группе буревестников есть репродуктивно изолированные (на Азорских островах ) и зарождающиеся виды (на других архипелагах), вызванные прохладным и теплым сезоном размножения. [77] [78] [79] |
| Howea belmoreana и H. forsteriana | Генетически контролируемое время цветения привело (в сочетании с различным уровнем pH почвы ) к репродуктивной изоляции двух видов пальм на острове Лорд-Хау . [80] |
| Эрисифа-убийца | Обладает доказательствами изоляции из-за временных различий между видами-хозяевами Vitis vinifera . [81] |
| Онкоринхус горбуша | Четные и нечетные двухлетние жизненные циклы в сочетании с сезонным размножением горбуши привели к генетической дифференциации между двумя популяциями. [82] [83] [84] |
| Magicicada виды. | Группы пар видов цикад с 13- и 17-летним жизненным циклом (всего семь видов) появляются для размножения, разделенные большими временными интервалами между сезонами размножения. [85] [86] [87] Только каждые 221 год 13- и 17-летние циклы совпадают, когда обе пары возникают одновременно. [62] |
| Немного антитрогуса | Две когорты жуков демонстрируют генетическую дифференциацию жизненных циклов, разделенных двухлетними интервалами. [88] |
| Оэнейс был полубогом | Двухлетний жизненный цикл селекционных групп видов бабочек привел к генетической дифференциации. [89] |
| Бамбусовые | Бамбук подвергается семелпарному размножению , при котором они живут годами, прежде чем начнут массовое цветение сразу же . Это может произойти в разные годы и в разных местах. Считается, что аллохронные пятна привели к диверсификации глобальных видов бамбука. [90] [91] [92] |
презиготические формы экологической Другие изоляции
Предполагается, что отбор против мигрантов, или нежизнеспособность иммигрантов, является формой экологической изоляции. Этот тип видообразования предполагает низкую выживаемость мигрантов между популяциями из-за отсутствия у них адаптации к чужеродным местообитаниям. [4] Существует мало понимания взаимосвязи между пост-спариванием, презиготической изоляцией и экологией. [4] Изоляция после спаривания происходит между процессом совокупления (или опыления) и оплодотворением, также известной как гаметическая изоляция. [1] : 232 Некоторые исследования, посвященные изоляции гамет у дрозофилы , плодовых мушек [93] земляные сверчки, [94] и Хелиантус растения [95] предполагают, что экология может сыграть свою роль; однако это неопределенно. [4]
Постзиготические формы экологической изоляции
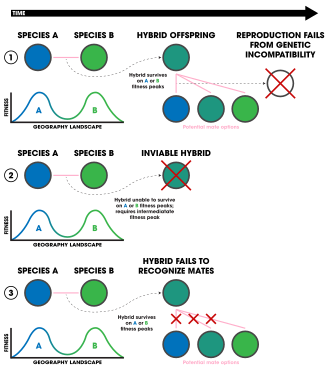
1. Экологически независимая постзиготическая изоляция.
2. Экологически зависимая постзиготическая изоляция.
3. Отбор против гибридов.
Экологически независимая постзиготическая изоляция возникает в результате генетической несовместимости между двумя гибридными особями одного вида. [96] Считается, что в некоторых случаях гибриды имеют более низкую приспособленность, особенно в зависимости от среды, в которой они живут. [96] Например, в экстремальных условиях с ограниченными экологическими нишами, которые можно использовать, необходима высокая приспособленность, тогда как, если в среде много ниш, особи с более низкой приспособленностью могут выжить дольше. Некоторые исследования показывают, что эта несовместимость является причиной экологического видообразования, поскольку они могут быстро развиваться в результате дивергентного отбора. [4]
Экологически зависимая постзиготическая изоляция возникает в результате снижения приспособленности гибрида из-за его положения в экологической нише. [4] — то есть родительские виды занимают несколько разные ниши, но их гибридному потомству в конечном итоге требуется ниша, представляющая собой смесь двух из них, которая обычно не существует (с точки зрения фитнес-ландшафта). Это было обнаружено в популяциях колюшек ( Gasterosteus aculeatus ). [97] [98] жуки-кувшинки ( Galerucella nymphaeae ), [99] гороховая тля, [100] и тефритидные мухи ( Eurosta Solidaginis ). [101]
Отбор против гибридов иногда (возможно, к нему можно отнести и неэкологическое видообразование) можно рассматривать как форму экологической изоляции, если он обусловлен экологическим механизмом. [4] Например, гибридное потомство может рассматриваться как «менее привлекательное» для партнеров из-за промежуточных сексуальных проявлений или различий в сексуальном общении. Конечным результатом является то, что гены каждой родительской популяции не могут смешиваться, поскольку они передаются гибридом, который вряд ли сможет воспроизводиться. Этот образец полового отбора против гибридного потомства был обнаружен у Heliconius . бабочек [4] Два вида H. cydno и H. melpomene симпатрически распространены в Южной Америке и нечасто гибридизуются. [102] Когда они гибридизуются, этот вид демонстрирует сильное ассортативное спаривание из-за развитой мимикрии цветовой схемы, промежуточным звеном которой является гибридное потомство. [102] Подобные закономерности были обнаружены у златоглазок. [103] модели миграции популяций птиц Sylvia atricapilla , [104] пауки-волки ( Schizocosa ocreata и S. rovneri ) и их брачное поведение, [105] симпатрические донные и пресноводные колюшки ( комплекс Gasterosteus aculeatus ), [106] и панамские бабочки Anartia fatima и A. amathea . [107] Цветы, в которых опылители различают гибриды, также продемонстрировали эту закономерность у обезьяньих цветов ( Erythranthe lewisii и Erythranthe cardinalis ). [108] и у двух видов группы ирисов Луизианы: Iris fulva и I. hexagona . [109]
См. также [ править ]
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и Джерри А. Койн ; Х. Аллен Орр (2004), Видообразование , Sinauer Associates, стр. 1–545, ISBN 0-87893-091-4
- ↑ Перейти обратно: Перейти обратно: а б Джеймс М. Собел, Грейс Ф. Чен, Лорна Р. Ватт и Дуглас В. Шемске (2009), «Биология видообразования», Evolution , 64 (2): 295–315, doi : 10.1111/j.1558- 5646.2009.00877.x , PMID 19891628 , S2CID 10168162
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Дольф Шлютер (2009), «Свидетельства экологического видообразования и его альтернативы», Science , 323 (5915): 737–741, Бибкод : 2009Sci...323..737S , doi : 10.1126/science.1160006 , PMID 19197053 , S2CID 307207
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и Ховард Д. Рандл и Патрик Носил (2005), «Экологическое видообразование», Ecology Letters , 8 (3): 336–352, Бибкод : 2005EcolL...8..336R , doi : 10.1111/j.1461-0248.2004.00715 .x
- ^ Патрик Носил, Люк Дж. Хармон и Оле Зеехаузен (2009), «Экологические объяснения (неполного) видообразования», Тенденции в экологии и эволюции , 24 (3): 145–156, doi : 10.1016/j.tree.2008.10. 011 , ПМИД 19185951
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ↑ Перейти обратно: Перейти обратно: а б Патрик Носил (2012), Экологическое видообразование , Оксфорд: Oxford University Press, стр. 280, ISBN 978-0199587117
- ^ Гиттенбергер, Э. (1 августа 1991 г.). «А как насчет неадаптивной радиации?» . Биологический журнал Линнеевского общества . 43 (4): 263–272. дои : 10.1111/j.1095-8312.1991.tb00598.x . ISSN 0024-4066 .
- ↑ Перейти обратно: Перейти обратно: а б Рунделл, Ребекка Дж.; Прайс, Тревор Д. (1 июля 2009 г.). «Адаптивная радиация, неадаптивная радиация, экологическое видообразование и неэкологическое видообразование» . Тенденции в экологии и эволюции . 24 (7): 394–399. дои : 10.1016/j.tree.2009.02.007 . ISSN 0169-5347 . ПМИД 19409647 .
- ^ Сюй, Минцзы; Шоу, Керри Л. (05 февраля 2020 г.). «Пространственное смешивание между призывающими самцами двух близкородственных симпатрических сверчков предполагает полезные гетероспецифические взаимодействия в неадаптивном излучении» . Журнал наследственности . 111 (1): 84–91. doi : 10.1093/jhered/esz062 . ISSN 0022-1503 . ПМИД 31782960 .
- ^ Велленройтер, Марен ; Санчес-Гильен, Роза Ана (2016). «Неадаптивная радиация у стрекоз» . Эволюционные приложения . 9 (1): 103–118. Бибкод : 2016EvApp...9..103W . дои : 10.1111/eva.12269 . ISSN 1752-4571 . ПМЦ 4780385 . ПМИД 27087842 .
- ^ Чекански-Мойр, Джесси Э.; Рунделл, Ребекка Дж. (01 мая 2019 г.). «Экология неэкологического видообразования и неадаптивных излучений» . Тенденции в экологии и эволюции . 34 (5): 400–415. дои : 10.1016/j.tree.2019.01.012 . ISSN 0169-5347 . ПМИД 30824193 . S2CID 73494468 .
- ^ Киркпатрик, Марк и Равинье, Виржини (2002), «Видообразование путем естественного и полового отбора: модели и эксперименты», The American Naturalist , 159 : S22–S35, doi : 10.1086/338370 , PMID 18707367 , S2CID 16516804
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Дольф Шлютер (2001), «Экология и происхождение видов», Trends in Ecology & Evolution , 16 (17): 327–380, doi : 10.1016/S0169-5347(01)02198-X , PMID 11403870 , S2CID 9845298
- ↑ Перейти обратно: Перейти обратно: а б Дольф Шлютер (2000), Экология адаптивной радиации , Oxford University Press, ISBN 0198505221
- ^ Питер А. Абрамс (2000), «Смена характера видов добычи, разделяющих хищников», American Naturalist , 154 (4): 45–61, doi : 10.1086/303415 , PMID 29592581 , S2CID 4387648
- ^ Трой Дэй и Кайл А. Янг (2004), «Конкурентная и стимулирующая эволюционная диверсификация», BioScience , 54 (2): 101–109, doi : 10.1641/0006-3568(2004)054[0101:CAFED]2.0.CO; 2 , S2CID 41757319
- ^ Майкл Дебели и Ульф Дикманн (2000), «Эволюционное ветвление и симпатрическое видообразование, вызванное различными типами экологических взаимодействий», American Naturalist , 156 (4): 77–101, doi : 10.1086/303417 , PMID 29592583 , S2CID 4409112
- ^ Мария Р. Серведио; Мохамед А. Ф. Нур (2003), «Роль подкрепления в видообразовании: теория и данные», Ежегодный обзор экологии, эволюции и систематики , 34 : 339–364, doi : 10.1146/annurev.ecolsys.34.011802.132412
- ^ Марк Киркпатрик (2001), «Подкрепление во время экологического видообразования», Proceedings of the Royal Society B , 268 (1473): 1259–1263, doi : 10.1098/rspb.2000.1427 , PMC 1088735 , PMID 11410152
- ^ Тами М. Панхуиса, Роджер Батлин, Марлен Зук и Том Трегенза (2001), «Половой отбор и видообразование», Тенденции в экологии и эволюции , 16 (7): 364–371, doi : 10.1016/S0169-5347(01) 02160-7 , ПМИД 11403869
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Рассел Ланде (1982), «Быстрое возникновение сексуальной изоляции и расхождения характеров в группе», Evolution , 36 (2): 213–223, doi : 10.1111/j.1558-5646.1982.tb05034.x , PMID 28563171 , S2CID 20428163
- ↑ Перейти обратно: Перейти обратно: а б Джанетт Венрик Бофман (2002), «Как сенсорное влечение может способствовать видообразованию», Trends in Ecology & Evolution , 17 (12): 571–577, doi : 10.1016/S0169-5347(02)02595-8
- ^ Джанетт Венрик Бофман (2001), «Дивергентный половой отбор усиливает репродуктивную изоляцию колюшек», Nature , 411 (6840): 944–948, Bibcode : 2001Natur.411..944B , doi : 10.1038/35082064 , PMID 11418857 , S2CID 56697 95
- ^ Родерик Д.М. Пейдж (2005). «Ковидообразование». ЭЛС . Чичестер: John Wiley & Sons Ltd. doi : 10.1038/npg.els.0004124 . ISBN 0470016175 .
- ^ Хан Ван, Э. Дюрант Макартур, Стюарт К. Сандерсон, Джон Х. Грэм и Д. Карл Фриман (1997), «Узкая гибридная зона между двумя подвидами большой полыни ( Artemisia Tridentata : Asteraceae). IV. Эксперименты по взаимной трансплантации» , Evolution , 51 (1): 95–102, doi : 10.1111/j.1558-5646.1997.tb02391.x , PMID 28568779 , S2CID 19274910
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Йенс Клаузен, Дэвид Д. Кек и Уильям М. Хизи (1940), Экспериментальные исследования природы видов. I. Влияние разнообразной окружающей среды на растения западной части Северной Америки , Вашингтон, округ Колумбия: Вашингтонский институт Карнеги, ISBN 9780608062204
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сальваторе Дж. Тавормина (1982), «Симпатрическая генетическая дивергенция минирующего насекомого Liriomyza brassicae (Dipter: Agromyzidae)», Evolution , 36 (3): 523–534, doi : 10.1111/j.1558-5646.1982.tb05073. х , PMID 28568038 , S2CID 29041437
- ^ Харуо Катакура, Миюки Сиои и Юми Кира (1989), «Репродуктивная изоляция по специфичности хозяина в паре жуков-фитофагов-божьих коровок», Evolution , 43 (5): 1045–1053, doi : 10.1111/j.1558-5646.1989.tb02549 .x , PMID 28564150 , S2CID 22996209
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тимоти П. Крейг, Джоан К. Итами, Уоррен Г. Абрахамсон, Джон Д. Хорнер (1993), «Поведенческие доказательства формирования расы-хозяина у Eurosta Solidaginis », Evolution , 47 (6): 1696–1710, doi : 10.1111 /j.1558-5646.1993.tb01262.x , PMID 28567992 , S2CID 205778515
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Джеффри Л. Федер, Сьюзан Б. Опп, Брайан Влазло, Кэтрин Рейнольдс, Уэсли Го и Стив Списак (1994), «Верность хозяина является эффективным барьером перед спариванием между симпатрическими расами яблочной личинки», PNAS , 91 (17) : 7990–7994, Бибкод : 1994PNAS...91.7990F , doi : 10.1073/pnas.91.17.7990 , PMC 44530 , PMID 11607491
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Чарльз Линн младший, Джеффри Л. Федер, Сатоши Нодзима, Хэтти Р. Дамброски, Стюарт Х. Берлохер и Венделл Рулофс (2003), «Дискриминация по запаху фруктов и формирование симпатической расы хозяев у Rhagoletis », PNAS , 100 (20): 11490–11493, Bibcode : 2003PNAS..10011490L , doi : 10.1073/pnas.1635049100 , PMC 208785 , PMID 14504399
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ↑ Перейти обратно: Перейти обратно: а б Дэниел Дж. Фанк (1998), «Выделение роли естественного отбора в видообразовании: адаптация хозяина и половая изоляция у Neochlamisus bebbianae листоедов », Evolution , 52 (6): 1744–1759, doi : 10.1111/j.1558-5646.1998 .tb02254.x , PMID 28565322 , S2CID 22704901
- ^ Сара Виа (1999), «Репродуктивная изоляция между симпатрическими расами гороховой тли. I. Ограничение потока генов и выбор среды обитания», Evolution , 53 (5): 1446–1457, doi : 10.1111/j.1558-5646.1999.tb05409.x , PMID 28565574 , S2CID 28392433
- ^ Р. Крус, М. Карбальо, П. Конде-Падин и Э. Ролан-Альварес (2004), «Тестирование альтернативных моделей половой изоляции в природных популяциях Littorina saxatilis : косвенная поддержка побочного экологического видообразования?» , Журнал эволюционной биологии , 17 (2): 288–293, doi : 10.1111/j.1420-9101.2003.00689.x , PMID 15009262 , S2CID 23589841
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Джеффри С. Маккиннон, Сейичи Мори, Бенджамин К. Блэкман, Лиор Дэвид, Дэвид М. Кингсли, Лея Джеймисон, Дженнифер Чоу и Дольф Шлютер (2004), «Доказательства роли экологии в видообразовании», Nature , 429 (6989): 294–298 , Bibcode : 2004Natur.429..294M , doi : 10.1038/nature02556 , PMID 15152252 , S2CID 2744267
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ JW Boughman (2001), «Дивергентный половой отбор усиливает репродуктивную изоляцию у колюшек», Nature , 411 (6840): 944–948, Bibcode : 2001Natur.411..944B , doi : 10.1038/35082064 , PMID 11418857 , S2CID 5669795
- ^ Говард. Д. Рандл, Л. Нагель, Дж. Венрик Бофман и Д. Шлютер (2000), «Естественный отбор и параллельное видообразование у симпатрических колюшек», Science , 287 (5451): 306–308, Bibcode : 2000Sci ... 287 ..306R , doi : 10.1126/science.287.5451.306 , PMID 10634785 , S2CID 7696251
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Лаура Нагель и Дольф Шлютер (1998), «Размер тела, естественный отбор и видообразование колюшек», Evolution , 52 (1): 209–218, doi : 10.1111/j.1558-5646.1998.tb05154.x , PMID 28568156 , S2CID 37489257
- ^ Патрик Носил, Бернард Дж. Креспи и Кристина П. Сандовал (2002), «Адаптация растения-хозяина стимулирует параллельную эволюцию репродуктивной изоляции», Nature , 417 (6887): 440–443, Bibcode : 2002Natur.417..440N , doi : 10.1038/417440a , PMID 12024213 , S2CID 4421774
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ П. Носил, Б. Дж. Креспи и К. П. Сандовал (2003), «Репродуктивная изоляция, обусловленная комбинированным эффектом экологической адаптации и подкрепления», Proceedings of the Royal Society B , 270 (1527): 1911–1918, doi : 10.1098/rspb. 2003.2457 , PMC 1691465 , PMID 14561304
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Крис Д. Джиггинс, Рассел Э. Нэйсбит, Ребекка Л. Коу и Джеймс Маллет (2001), «Репродуктивная изоляция, вызванная мимикрией цветового рисунка» (PDF) , Nature , 411 (6835): 302–305, Бибкод : 2001Natur.411 ..302J , doi : 10.1038/35077075 , PMID 11357131 , S2CID 2346396
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ↑ Перейти обратно: Перейти обратно: а б Аброл ДП (2012). «Взаимодействие непчелиных опылителей и растений». Биология опыления . Том. Глава 9. С. 265–310. дои : 10.1007/978-94-007-1942-2_9 . ISBN 978-94-007-1941-5 .
- ^ Стюарт, Алисса Б.; Дудаш, Мишель Р. (01 января 2018 г.). «Стратегии добывания нектара летучих мышей Старого Света широкого и специального назначения в ответ на меняющиеся во времени цветочные ресурсы». Биотропика . 50 (1): 98–105. Бибкод : 2018Биотр..50...98С . дои : 10.1111/btp.12492 . S2CID 90515964 .
- ↑ Перейти обратно: Перейти обратно: а б с Верн Грант (1971), Видообразование растений , Нью-Йорк: Издательство Колумбийского университета, стр. 432, ISBN 978-0231083263
- ^ Роберт Л. Дресслер (1968), «Опыление пчелами-эуглоссинами» , « Эволюция » , 22 (1): 202–210, doi : 10.2307/2406664 , JSTOR 2406664 , PMID 28564982
- ^ Назия Сулеман, Стив Саит и Стивен Г. Комптон (2015), «Женский инжир как ловушка: их влияние на динамику экспериментального сообщества опылителей и паразитоидов фигового дерева» (PDF) , Acta Oecologica , 62 : 1–9, Bibcode : 2015AcO....62....1S , doi : 10.1016/j.actao.2014.11.001
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Пеллмир, Олле; Томпсон, Джон Н.; Браун, Джонатан М.; Харрисон, Ричард Г. (1996). «Эволюция опыления и мутуализма в линии юкки». Американский натуралист . 148 (5): 827–847. дои : 10.1086/285958 . JSTOR 2463408 . S2CID 84816447 .
- ^ HD Брэдшоу-младший и Дуглас В. Шемске (2003), «Замена аллелей в локусе цвета цветка приводит к смене опылителей у обезьяньих цветов», Nature , 426 (6963): 176–178, Bibcode : 2003Natur.426..176B , doi : 10.1038/nature02106 , PMID 14614505 , S2CID 4350778
- ^ Джастин Рэмси, HD Брэдшоу и Дуглас В. Шемске (2003), «Компоненты репродуктивной изоляции между Monkeyflowers Mimulus lewisii и M. cardinalis (Phrymaceae)», Evolution , 57 (7): 1520–1534, doi : 10.1554/01 -352 , PMID 12940357 , S2CID 198156112
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Скотт А. Ходжес и Майкл Л. Арнольд (1994), «Цветочная и экологическая изоляция между Aquilegia formosa и Aquilegia pubescens» (PDF) , Proceedings of the National Academy of Sciences , 91 (7): 2493–2496, Bibcode : 1994PNAS. ..91.2493H , doi : 10.1073/pnas.91.7.2493 , PMC 43395 , PMID 8146145
- ^ AYK Альберт и Д. Шлютер (2004), «Смещение репродуктивного характера предпочтений самца колюшки: подкрепление или прямой отбор?», Evolution , 58 (5): 1099–1107, doi : 10.1111/j.0014-3820.2004.tb00443.x , PMID 15212390 , S2CID 13882516
- ^ Джеффри Л. Федер, Стюарт Х. Берлохер, Джозеф Б. Ротел, Хэтти Дамброски, Джеймс Дж. Смит, Уильям Л. Перри, Весна Гаврилович, Кеннет Э. Филчак, Хуан Рулл и Мартин Алуха (2003), «Аллопатрическое генетическое происхождение симпатрических сдвигов растения-хозяина» и формирование расы у Rhagoletis», Proceedings of the National Academy of Sciences of the United States of America , 100 (18): 10314–10319, Bibcode : 2003PNAS..10010314F , doi : 10.1073/pnas.1730757100 , PMC 193558 , PMID 1292 8500
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Роберт Уильям Круден (1972), «Биология опыления Nemophila menziesii (Hydrophyllaceae) с комментариями к эволюции олиголектических пчел», Evolution , 26 (3): 373–389, doi : 10.1111/j.1558-5646.1972.tb01943.x , PMID 28563062 , S2CID 39629939
- ^ Мазер, Сьюзан Дж. и Мид, Дэниел Э. (2000). «Глава 7: Географические различия в размерах цветков дикой редьки: потенциальная роль опылителей в дифференциации популяций». В Муссо, Тимоти А.; Синерво, Барри и Эндлер, Джон (ред.). Адаптивная генетическая изменчивость в дикой природе . Издательство Оксфордского университета. стр. 157–186. ISBN 978-0195121834 .
- ^ Кэндис Гален (1989), «Измерение опосредованного опылителем отбора по морфометрическим признакам цветка: шмели и пилот альпийского неба, Polemonium viscosum», Evolution , 43 (4): 882–890, doi : 10.2307/2409315 , JSTOR 2409315 , PMID 28564200
- ^ Рассел Б. Миллер (1981), «Бражники и географические закономерности цветочной изменчивости Aquilegia caerulea», Evolution , 35 (4): 763–774, doi : 10.1111/j.1558-5646.1981.tb04936.x , PMID 28563131 , S2CID 38127528
- ^ С.Д. Джонсон (1997), «Экотипы опыления Satyrium Hallackii (Orchidaceae) в Южной Африке», Ботанический журнал Линнеевского общества , 123 (3): 225–235, doi : 10.1111/j.1095-8339.1997.tb01415.x
- ^ С.Д. Джонсон и К.Е. Штайнер (1997), «Опыление длинноязычными мухами и эволюция длины цветочных шпор в комплексе Disa draconis (Orchidaceae)», Evolution , 51 (1): 45–53, doi : 10.1111/j.1558- 5646.1997.tb02387.x , PMID 28568792 , S2CID 43420068
- ^ Шила П. Турбек, Элизабет С.К. Скордато и Ребекка Дж. Сафран (2018), «Роль сезонной миграции в демографической дивергенции и репродуктивной изоляции», Тенденции в экологии и эволюции , 33 (3): 164–175, doi : 10.1016 /j.tree.2017.11.008 , PMID 29289354
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Клаудия Гермес, Раэнн Меттлер, Диего Сантьяго-Аларкон, Гернот Сегельбахер и Х. Мартин Шефер (2015), «Пространственная изоляция и временные изменения в физической форме и состоянии способствуют расхождению в миграционном разделении», PLOS ONE , 10 (12): e0144264 , Bibcode : 2015PLoSO..1044264H , doi : 10.1371/journal.pone.0144264 , PMC 4681481 , PMID 26656955
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тревор Прайс (2008), Видообразование у птиц , издательство Roberts and Company, стр. 1–64, ISBN 978-0-9747077-8-5
- ↑ Перейти обратно: Перейти обратно: а б с д Ребекка С. Тейлор и Вики Л. Фризен (2017), «Роль аллохронии в видообразовании», Molecular Ecology , 26 (13): 3330–3342, Бибкод : 2017MolEc..26.3330T , doi : 10.1111/mec.14126 , ПМИД 28370658 , С2КИД 46852358
- ^ Эндрю П. Хендри и Трой Дэй (2005), «Структура населения, обусловленная репродуктивным временем: изоляция временем и адаптация временем», Molecular Ecology , 14 (4): 901–916, Бибкод : 2005MolEc..14..901H , doi : 10.1111/j.1365-294X.2005.02480.x , PMID 15773924 , S2CID 8226535
- ^ Х. Фуками, М. Омори, К. Симойке, Т. Хаясибара и М. Хатта (2003), «Экологические и генетические аспекты репродуктивной изоляции в зависимости от времени нереста кораллов Acropora», Marine Biology , 142 (4): 679. –684, Bibcode : 2003MarBi.142..679F , doi : 10.1007/s00227-002-1001-8 , S2CID 81981786
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Н. Ноултон, Дж. Л. Мате, Х. М. Гусман, Р. Роуэн и Дж. Хара (1997), «Прямые доказательства репродуктивной изоляции среди трех видов комплекса Montastraea annularis в Центральной Америке (Панама и Гондурас)», Морская биология , 127 (4): 705–711, Bibcode : 1997MarBi.127..705K , doi : 10.1007/s002270050061 , S2CID 37997956
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Эндрю П. Хендри, Оле К. Берг и Томас П. Куинн (1999), «Зависимость от условий и адаптация ко времени: дата размножения, история жизни и распределение энергии в популяции лосося», Oikos , 85 (3 ): 499–514, Bibcode : 1999Oikos..85..499H , doi : 10.2307/3546699 , JSTOR 3546699
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Эндрю П. Хендри, Иоланда Э. Морби, Оле К. Берг и Джон К. Венбург (2004), «Адаптивная изменчивость старения: продолжительность репродуктивной жизни в популяции дикого лосося», Proceedings of the Royal Society B: Biological Sciences , 271 (1536): 259–266, doi : 10.1098/rspb.2003.2600 , PMC 1691593 , PMID 15058436
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ EK Fillatre, P. Etherton и DD Heath (2003), «Бимодальное распределение пробега в северной популяции нерки (Oncorhynchus nerka): история жизни и генетический анализ во временном масштабе», Molecular Ecology , 12 (7): 1793 –1805, Bibcode : 2003MolEc..12.1793F , doi : 10.1046/j.1365-294x.2003.01869.x , PMID 12803632 , S2CID 25772120
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ К.Пиментел, Т.Кальвао, М.Сантос, К.Феррейра, М.Невес и Ж.-О.Нильссон (2006), «Создание и расширение Thaumetopoeapityocampa (Den. & Schiff.) (Lep. Notodontidae ) популяция со смещенным жизненным циклом в продуктивном сосновом лесу, Центрально-Прибрежная Португалия», Forest Ecology and Management , 233 (1): 108–115, doi : 10.1016/j.foreco.2006.06.005
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хелена М. Сантос, Мария-Роза Пайва, Сусана Роша, Кэрол Кердельуэ и Мануэла Бранко (2013), «Фенотипическое расхождение репродуктивных признаков популяции бабочек, испытывающих фенологический сдвиг», Ecology and Evolution , 3 (15): 5098–5108. , Bibcode : 2013EcoEv...3.5098S , doi : 10.1002/ece3.865 , PMC 3892371 , PMID 24455139
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мануэла Бранко, Мария-Роза Пайва, Хелена Мария Сантос, Кристиан Бурбан и Кэрол Кердельуэ (2017), «Экспериментальные доказательства наследственного репродуктивного времени в двух аллохронных популяциях сосновой процессионной моли», Insect Science , 24 (2): 325–335 , Bibcode : 2017InsSc..24..325B , doi : 10.1111/1744-7917.12287 , PMID 26530538 , S2CID 9091980
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сатоши Ямамото и Тейджи Сота (2012), «Параллельная аллохронная дивергенция у зимней бабочки из-за нарушения репродуктивного периода из-за суровых зим», Molecular Ecology , 21 (1): 174–183, Bibcode : 2012MolEc..21..174Y , doi : 10.1111/j.1365-294X.2011.05371.x , PMID 22098106 , S2CID 23572464
- ^ Патрик Эббот и Джеймс Х. Уитготт (2004), «Филогенетические и молекулярные доказательства аллохронного видообразования у тлей, образующих галлы (пузырчатка)», Evolution , 58 (3): 539–553, doi : 10.1111/j.0014-3820.2004.tb01677 .x , PMID 15119438 , S2CID 25277034
- ^ Джеффри Б. Джой и Бернард Дж. Креспи (2007), «Адаптивная радиация насекомых, вызывающих галл, в пределах одного вида растений-хозяев», Evolution , 61 (4): 784–795, doi : 10.1111/j.1558-5646.2007.00069 .x , PMID 17439611 , S2CID 16864372
- ^ Натали Л. Россер (2015), «Асинхронный нерест в симпатрических популяциях твердого коралла выявляет загадочные виды и древние генетические линии», Molecular Ecology , 24 (19): 5006–5019, Bibcode : 2015MolEc..24.5006R , doi : 10.1111/ mec.13372 , PMID 26339867 , S2CID 13151100
- ^ Кристофер Э. Берд, Бренден С. Холланд, Брайан В. Боуэн и Роберт Дж. Тунен (2011), «Диверсификация симпатрических блюдечек, нерестящихся широковещательно (Cellana spp.) На Гавайском архипелаге», Молекулярная экология , 20 (10): 2128 –2141, Bibcode : 2011MolEc..20.2128B , doi : 10.1111/j.1365-294X.2011.05081.x , PMID 21481050 , S2CID 23432529
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Л. Р. Монтейро (1998), «Видообразование посредством временной сегрегации популяций мадейрских кастро (Oceanodroma castro) на Азорских островах?», « Философские труды Лондонского королевского общества». Серия B, Биологические науки , 353 (1371): 945–953, doi : 10.1098/rstb.1998.0259 , PMC 1692297.
- ^ В.Л. Фризен, А.Л. Смит, Э. Гомес-Диас, М. Болтон, Р.В. Фернесс, Дж. Гонсалес-Солис и Л.Р. Монтейро (2007), «Симпатрическое видообразование путем аллохронии у морских птиц» (PDF) , PNAS , 104 (47) ): 18589–18594, Бибкод : 2007PNAS..10418589F , doi : 10.1073/pnas.0700446104 , PMC 2141821 , PMID 18006662
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Марк Болтон, Андреа Л. Смит, Елена Гомес-Диас, Вики Л. Фризен, Рената Медейрос, Жоэль Брид, Хосе Л. Роскалес и Роберт В. Фернесс (2008), «Буревестник Монтейро Oceanodroma monteiroi: новый вид из Азорские острова», Ibis , 150 (4): 717–727, doi : 10.1111/j.1474-919X.2008.00854.x
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Винсент Саволайнен, Мари-Шарлотта Анстетт, Кристиан Лексер, Ян Хаттон, Джеймс Дж. Кларксон, Мария В. Норуп, Мартин П. Пауэлл, Дэвид Спрингейт, Николас Саламин и Уильям Дж. Бейкер (2006), «Симпатрическое видообразование пальм на океаническом острове» , Nature , 441 (7090): 210–213, Бибкод : 2006Natur.441..210S , doi : 10.1038/nature04566 , PMID 16467788 , S2CID 867216
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Жослен Монтарри, Филипп Картоларо, Сильви Ришар-Сервера и Франсуа Дельмотт (2009), «Пространственно-временное распределение генетических групп Erysiphenecator и их взаимосвязь с уровнями заболеваний на виноградниках», Европейский журнал патологии растений , 123 : 61–70, Bibcode : 2009EJPP..123...61M , CiteSeerX 10.1.1.511.8215 , doi : 10.1007/s10658-008-9343-9 , S2CID 13114251
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Лев А. Животовский, А. Дж. Гарретт, А. Дж. МакГрегор, М. К. Глубоковский и Маркус В. Фельдман (1994), «Дифференциация генов у тихоокеанского лосося (Oncorhynchus Sp.): факты и модели применительно к горбуше (O. Gorbuscha)», Канадский журнал рыболовства и водных наук , 51 : 223–232, doi : 10.1139/f94-308.
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Д. Чуриков и А.Дж. Гарретт (2002), «Сравнительная филогеография двух выводков горбуши: анализ, основанный на генеалогии митохондриальной ДНК», Molecular Ecology , 11 (6): 1077–1101, Bibcode : 2002MolEc..11.1077C , doi : 10.1046/j.1365-294x.2002.01506.x , PMID 12030984 , S2CID 24965183
- ^ Мортен Т. Лимборг, Райан К. Уэплс, Джеймс Э. Сиб и Лиза В. Сиб (2014), «Временно изолированные линии горбуши обнаруживают уникальные признаки отбора в различных пулах постоянных генетических вариаций», Журнал наследственности , 105 (6): 835–845, doi : 10.1093/jhered/esu063 , PMID 25292170
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Д. К. Маршалл и Дж. Р. Кули (2000), «Смещение репродуктивных признаков и видообразование у периодических цикад с описанием новых видов, 13-летняя Magicicada neotredecem», Evolution , 54 (4): 1313–1325, doi : 10.1111/j.0014 -3820.2000.tb00564.x , hdl : 2027.42/73691 , PMID 11005298 , S2CID 28276015
- ^ К. Саймон, Дж. Тан, С. Далвади, Г. Стейли, Дж. Дениега и Т. Р. Уннаш (2000), «Генетические доказательства ассортативного спаривания между 13-летними цикадами и симпатрическими» 17-летними цикадами с 13-летней жизнью. циклы» обеспечивает поддержку аллохронного видообразования», Evolution , 54 (4): 1326–1336, doi : 10.1111/j.0014-3820.2000.tb00565.x , PMID 11005299 , S2CID 19105047
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тейджи Сота, Сатоши Ямамото, Джон Р. Кули, Кэти Б.Р. Хилл, Крис Саймон и Джин Йошимура (2013), «Независимое расхождение 13- и 17-летних жизненных циклов среди трех периодических линий цикад», PNAS , 110 (17) : 6919–6924, Бибкод : 2013PNAS..110.6919S , doi : 10.1073/pnas.1220060110 , PMC 3637745 , PMID 23509294
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Д. П. Логан, П. Г. Олсопп и М. П. Залуки (2003), «Зимовка, распределение почвы и фенология Canegrub Чайлдерса, Antitrogus parvulus (Coleoptera: Scarabaeidae) в сахарном тростнике Квинсленда», Бюллетень энтомологических исследований , 93 (4): 307–314, doi : 10.1079/ber2003245 , PMID 12908916
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ А.Э. Градиш, Н. Кейгобади и Г.В. Отис (2015), «Популяционная генетическая структура и генетическое разнообразие находящейся под угрозой исчезновения арктической бабочки Белой горы (Oeneis melissa semidea)», Conservation Genetics , 16 (5): 1253–1264, Bibcode : 2015ConG ...16.1253G , doi : 10.1007/s10592-015-0736-y , S2CID 13307002
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мадхав Гаджил и С. Нарендра Прасад (1984), «Экологические детерминанты эволюции жизненного цикла двух индийских видов бамбука», Biotropica , 16 (3): 161–172, Бибкод : 1984Biotr..16..161G , doi : 10.2307/ 2388050 , JSTOR 2388050
- ^ Дональд К. Франклин (2004), «Синхронность и асинхронность: наблюдения и гипотезы волны цветения у долгоживущего семелеродящего бамбука», Journal of Biogeography , 31 (5): 773–786, Bibcode : 2004JBiog..31.. 773F , doi : 10.1111/j.1365-2699.2003.01057.x , S2CID 55279438
- ^ Анелена Л. де Карвальо, Брюс В. Нельсон, Милтон К. Бьянкини, Даниэла Планьоль, Татьяна М. Куплич и Дуглас К. Дейли (2013), «Леса юго-западной Амазонки с преобладанием бамбука: обнаружение, пространственный масштаб, жизненный цикл Длина и волны цветения", PLOS ONE , 8 (1): e54852, Bibcode : 2013PLoSO...854852C , doi : 10.1371/journal.pone.0054852 , PMC 3554598 , PMID 23359438
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кэтрин С.К. Прайс, Кристин Х. Ким, Карина Дж. Гронлунд и Джерри А. Койн (2001), «Загадочная репродуктивная изоляция в комплексе видов Drosophila simulans», Evolution , 55 (1): 81–92, doi : 10.1111/ j.0014-3820.2001.tb01274.x , PMID 11263748 , S2CID 18100324
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Дэниел Дж. Ховард, Памела Г. Грегори, Джиминг Чу и Майкл Л. Кейн (1998), «Преимущество сперматозоидов одного вида является эффективным барьером для гибридизации между близкородственными видами», Evolution , 52 (2): 511–516, doi : 10.1111/j.1558-5646.1998.tb01650.x , PMID 28568320 , S2CID 8184734
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Лорен Х. Ризеберг, Андре М. Дерошерс и Сью Дж. Юн (1995), «Межвидовая конкуренция пыльцы как репродуктивный барьер между симпатрическими видами Helianthus (Asteraceae)», American Journal of Botany , 82 (4): 515–519, дои : 10.2307/2445699 , JSTOR 2445699
- ↑ Перейти обратно: Перейти обратно: а б Ховард Д. Рандл, Майкл К. Уитлок (2001), «Генетическая интерпретация экологически зависимой изоляции», Evolution , 55 (1): 198–201, doi : 10.1111/j.0014-3820.2001.tb01284.x , PMID 11263739 , S2CID 14710367
- ^ Говард Д Рандл (2002), «Тест на экологически зависимую изоляцию после спаривания между симпатрическими колюшками», Evolution , 56 (2): 322–329, doi : 10.1111/j.0014-3820.2002.tb01342.x , PMID 11926500 , S2CID 11550301
- ^ Тодд Хэтфилд и Дольф Шлютер (1999), «Экологическое видообразование колюшек: гибридная приспособленность, зависящая от окружающей среды», Evolution , 53 (3): 866–873, doi : 10.1111/j.1558-5646.1999.tb05380.x , PMID 28565618 , S2CID 10638478
- ^ Стефани М. Папперс, Жерар ван дер Вельде, Н. Йооп Оуборг и Ян М. ван Грюнендаль (2002), «Генетически обоснованные полиморфизмы в морфологии и истории жизни, связанные с предполагаемыми расами-хозяевами листоеда-кувшинки Galerucella nymphaeae», « Эволюция » , 56 (8): 1610–1621, doi : 10.1111/j.0014-3820.2002.tb01473.x , PMID 12353754 , S2CID 23891554
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сара Виа, А.С. Бук и С. Скиллман (2000), «Репродуктивная изоляция между дивергентными расами гороховой тли на двух хозяевах. II. Отбор против мигрантов и гибридов в родительской среде», Evolution , 54 (5): 1626–1637. , doi : 10.1111/j.0014-3820.2000.tb00707.x , PMID 11108590 , S2CID 26339284
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тимоти П. Крейг, Джон Д. Хорнер и Джоан К. Итами (1997), «Исследования гибридизации рас-хозяев Eurosta Solidaginis: последствия для симпатрического видообразования», Evolution , 51 (5): 1552–1560, doi : 10.1111/j. 1558-5646.1997.tb01478.x , PMID 28568625 , S2CID 6447741
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ↑ Перейти обратно: Перейти обратно: а б RE Naisbit, CD Jiggins и J. Mallet (2001), «Разрушительный половой отбор против гибридов способствует видообразованию между Heliconius cydno и Heliconius melpomene», Proceedings of the Royal Society B: Biological Sciences , 268 (1478): 1849–1854, doi : 10.1098/rspb.2001.1753 , PMC 1088818 , PMID 11522205
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Уэллс, Марта Мартинес и Генри, Чарльз (1998). «Песни, репродуктивная изоляция и видообразование загадочных видов насекомых: пример использования зеленых златоглазок». В Ховард, Дэниел Дж. и Берлочер, Стюарт Х. (ред.). Бесконечные формы: виды и видообразование . Издательство Оксфордского университета. стр. 217–233. ISBN 978-0195109016 .
- ^ Андреас Дж. Хельбиг (1991), «Мигрирующие на юго-восток и юго-запад популяции черной шапочки (Sylvia atricapilla) в Центральной Европе: ориентация птиц в зоне контакта», Journal of Evolutionary Biology , 4 (4): 657–670, doi : 10.1046/j.1420-9101.1991.4040657.x , S2CID 84847304
- ^ Гейл Э. Стрэттон и Джордж Уетц (1986), «Наследование ухаживающего поведения и его роль как репродуктивного изолирующего механизма у двух видов пауков-волков Schizocosa (Araneae; Lycosidae)», Evolution , 40 (1): 129– 141, doi : 10.1111/j.1558-5646.1986.tb05724.x , PMID 28564117 , S2CID 7755906
- ^ Стивен М. Вамози и Дольф Шлютер (1999), «Половой отбор против гибридов между симпатрическими видами колюшки: данные полевого эксперимента», Evolution , 53 (8): 874–879, doi : 10.1111/j.1558-5646.1999.tb05381. х , PMID 28565643 , S2CID 205781377
- ^ Н. Дэвис, А. Айелло, Дж. Маллет, А. Помианковски и Р. Е. Силберглайд (1997), «Видообразование двух неотропических бабочек: расширение правила Холдейна», Proceedings of the Royal Society B: Biological Sciences , 264 (1383): 845–851, Bibcode : 1997RSPSB.264..845D , doi : 10.1098/rspb.1997.0118 , PMC 1688429
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Дуглас В. Шемске и Х.Д. Брэдшоу младший (1999), «Предпочтение опылителей и эволюция цветочных признаков обезьяньих цветов (Mimulus)» (PDF) , Труды Национальной академии наук , 96 (21): 11910–11915, Bibcode : 1999PNAS...9611910S , doi : 10.1073/pnas.96.21.11910 , PMC 18386 , PMID 10518550
- ^ Саймон К. Эммс и Майкл Л. Арнольд (2000), «Различия между участками в характере посещения опылителей в гибридной зоне ирисов Луизианы», Oikos , 91 (3): 568–578, doi : 10.1034/j.1600 -0706.2000.910319.x