Трехиглая колюшка
| Трехиглая колюшка | |
|---|---|

| |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Актиноптеригии |
| Заказ: | Скорпениобразные |
| Семья: | Гастеростеиды |
| Род: | Гастеростеус |
| Разновидность: | Г. акулеатус
|
| Биномиальное имя | |
| Гастеростеус остроконечный | |
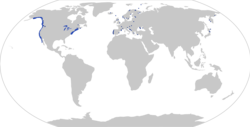 | |
| Распространение Gasterosteus aculeatus с наблюдениями (точки; по данным [ 2 ] ) и распространение (заштрихованная область; на основе https://nas.er.usgs.gov/queries/factsheet.aspx?SpeciesID=702 ) | |
| Синонимы [ 3 ] | |
| |

( Трехиглая колюшка Gasterosteus aculeatus ) — рыба, обитающая в большинстве внутренних и прибрежных вод к северу от 30° с.ш. Оно уже давно стало предметом научных исследований по многим причинам. Он демонстрирует большие морфологические вариации по всему ареалу, что идеально подходит для вопросов эволюции и популяционной генетики . Многие популяции являются анадромными (они живут в морской воде, но размножаются в пресной или солоноватой воде) и очень толерантны к изменениям солености, что представляет интерес для физиологов. Он демонстрирует сложное поведение при размножении (защита территории, строительство гнезда, уход за икрой и мальками) и может быть социальным (живет стаями вне сезона размножения ), что делает его популярным предметом исследований в этологии рыб и поведенческой экологии . Его адаптация к хищникам, взаимодействие хозяина и паразита, сенсорная физиология, репродуктивная физиология и эндокринология также хорошо изучены. Облегчает эти исследования тот факт, что трехиглую колюшку легко найти в природе и легко содержать в аквариумах. [ 4 ]
Описание
[ редактировать ]
Иногда этот вид может достигать длины 8 см (3,1 дюйма), но чаще встречается длина 3–4 см (1,2–1,6 дюйма) в зрелом возрасте. Тело сжато с боков. Основание хвоста тонкое. Хвостовой плавник имеет 12 лучей. В спинном плавнике 10–14 лучей; перед ним расположены три шипа, давшие рыбе ее название (хотя у некоторых особей их может быть только два или четыре). Третий шип (ближайший к спинному плавнику) значительно короче двух других. Задняя часть каждого шипа соединена с телом тонкой перепонкой. Анальный плавник имеет от восьми до 11 лучей, ему предшествует короткий шип. Брюшные плавники состоят всего из позвоночника и одного луча. Все шипы могут быть зафиксированы в вертикальном положении, что делает рыбу чрезвычайно трудной для проглатывания хищником. Грудные плавники крупные, с 10 лучами. Тело не имеет чешуи, но защищено костными пластинами на спине, боках и животе. Присутствует только одна вентральная пластина, но количество боковых пластин сильно варьируется в зависимости от ареала распространения и типов местообитаний (см. Ниже); обычно он выше у морских популяций (у некоторых пресноводных популяций вообще могут отсутствовать боковые пластины). [ 3 ]
Окраска спины варьируется, но имеет тенденцию к тускло-оливковому или серебристо-зеленому, иногда с коричневыми крапинками. Бока и живот серебристые. У самцов в период размножения глаза становятся голубыми, а нижняя часть головы, горло и передняя часть брюха становятся ярко-красными. Горло и живот у размножающихся самок могут слегка порозоветь. Однако в некоторых популяциях есть размножающиеся самцы, все черные. [ 5 ] или все белое. [ 6 ]

Среда обитания и распространение
[ редактировать ]Трехиглая колюшка встречается только в Северном полушарии, где обычно населяет прибрежные воды или пресноводные водоёмы. Он может жить как в пресной, солоноватой, так и в соленой воде. Предпочитает медленнотекущую воду с участками зарождающейся растительности. Его можно найти в канавах, прудах, озерах, заводях, тихих реках, защищенных бухтах, болотах и гаванях.
В Северной Америке он распространен вдоль восточного побережья от Чесапикского залива до южной половины Баффинова острова и западного берега Гудзонова залива, а также вдоль западного побережья от южной Калифорнии до западного берега Аляски и Алеутских островов. Его можно найти по всей Европе между 35 и 70° с.ш. В Азии распространение простирается от Японии и Корейского полуострова до Берингова пролива.
Его распространение можно было бы назвать циркумполярным, если бы не тот факт, что он отсутствует на северном побережье Сибири, северном побережье Аляски и арктических островах Канады.
Различия в морфологии и распространении
[ редактировать ]
три подвида признает В настоящее время МСОП :
- Г. а. aculeatus встречается на большей части ареала вида и является подвидом, который наиболее строго называют трехиглой колюшкой; его распространенное название в Великобритании — тиддлер , хотя иногда также используется «титлбат».
- Г. а. williamsoni , небронированная трехиглая колюшка , встречается только в Северной Америке; его признанный ареал - южная Калифорния , хотя были сделаны отдельные сообщения о том, что он встречается в Британской Колумбии и Мексике ;
- Г. а. santaeannae , колюшка Санта-Ана , также обитает только в Северной Америке.
Фактически эти подвиды представляют собой три примера из огромного диапазона морфологических вариаций, присутствующих у трехиглых колюшек. Гибриды между некоторыми из этих морф демонстрируют недостатки в добывании пищи , что является формой подкрепления в ходе видообразования . Это свидетельство видообразования путем подкрепления . [ 7 ]
В целом эти морфы делятся на две грубые категории: проходные и пресноводные формы:
Проходная форма проводит большую часть своей взрослой жизни, поедая планктон и рыбу в море, а затем возвращается в пресную воду для размножения. Взрослые рыбы обычно имеют длину от 6 до 10 см и имеют от 30 до 40 боковых броневых пластин по бокам. У них также есть длинные спинные и тазовые шипы. Анадромная форма морфологически схожа во всем Северном полушарии , так что анадромные рыбы из Балтики, Атлантики и Тихого океана очень похожи друг на друга.
Популяции трехиглой колюшки встречаются также в пресноводных озерах и ручьях. Эти популяции, вероятно, сформировались, когда анадромные рыбы начали проводить весь свой жизненный цикл в пресной воде и, таким образом, приспособились жить там круглый год. Пресноводные популяции чрезвычайно морфологически разнообразны до такой степени, что многие наблюдатели (и некоторые систематики) описали бы новый подвид трехиглой колюшки почти в каждом озере Северного полушария. Одним из постоянных различий между пресноводными популяциями и их анадромными предками является количество панциря, поскольку у большинства пресноводных рыб отсутствуют от двух до 12 боковых панцирных пластин, а также более короткие спинные и тазовые шипы. Однако между озерами существуют и большие морфологические различия. Одна из основных осей вариаций проходит между популяциями, обитающими в глубоких озерах с крутыми склонами, и популяциями, обитающими в небольших и мелких озерах. Рыбы в глубоких озерах обычно питаются планктоном в поверхностных водах и часто имеют большие глаза, короткое, тонкое тело и перевернутые челюсти. Некоторые исследователи называют это лимнетическая форма. Рыба из мелких озер питается в основном на дне озера и часто имеет длинное и тяжелое тело, относительно горизонтальные челюсти и маленькие глаза. Эти популяции называются бентосной формой.
Поскольку каждый водораздел, вероятно, был заселен анадромными колюшками отдельно, широко распространено мнение, что морфологически сходные популяции в разных водоразделах или на разных континентах развивались независимо. Уникальная популяция обитает в меромиктическом Розовом озере в парке Гатино , Квебек . Было замечено, что популяции быстро адаптируются к различным условиям, например, в озере Юнион , где колюшки теряли и восстанавливали броневые пластины в ответ на загрязнение в результате деятельности человека вокруг водораздела. [ 8 ] [ 9 ] [ 10 ] [ 11 ]
Одним из аспектов этой морфологической изменчивости является то, что в ряде озер присутствуют как пресноводный, так и донный тип, и они не скрещиваются друг с другом. Биологи-эволюционисты часто определяют виды как популяции, которые не скрещиваются друг с другом ( концепция биологических видов ), поэтому бентосные и лимнетики в каждом озере будут составлять отдельные виды. Эти пары видов являются прекрасным примером того, как адаптация к различным средам обитания (в данном случае кормление в поверхностных водах или на дне озера) может привести к появлению новых видов. Этот процесс получил название экологического видообразования . Этот тип пары видов встречается в Британской Колумбии . В самих озерах водятся только трехиглая колюшка и головорезная форель , и все они находятся на островах. К сожалению, пара в озере Хэдли на острове Ласкети была уничтожена в середине 1980-х годов из-за завоза хищного сома, а пара в озере Энос на острове Ванкувер начала скрещиваться и больше не является двумя отдельными видами. [ 12 ] Две оставшиеся пары обитают на острове Тексада , в озерах Пакстон и Прист-Лейк, и они занесены в список находящихся под угрозой исчезновения в Законе о канадских видах, находящихся под угрозой исчезновения. [ 13 ]
Другие пары видов, состоящие из хорошо бронированной морской формы и более мелкой, небронированной пресноводной формы, изучаются в прудах и озерах на юге центральной Аляски, которые когда-то были морской средой обитания, например, поднятыми во время землетрясения на Аляске 1964 года . Эволюционная динамика этих пар видов представляет собой модель процессов видообразования, которые произошли менее чем за 20 лет по крайней мере в одном озере. В 1982 году программа химического уничтожения, призванная освободить место для форели и лосося на озере Лоберг на Аляске, привела к гибели местных пресноводных популяций колюшек. Океанические колюшки, завезенные через близлежащий залив Кука, повторно заселили озеро. Всего за 12 лет, начиная с 1990 года, частота океанической формы неуклонно снижалась со 100% до 11%, в то время как разновидность с меньшим количеством плит увеличилась до 75% популяции, при этом еще небольшую долю составляли различные промежуточные формы. [ 14 ] Считается, что такая быстрая эволюция возможна благодаря генетическим вариациям, которые дают конкурентные преимущества для выживания в пресной воде, когда условия быстро меняются от соленой к пресной воде. Однако реальная молекулярная основа этой эволюции до сих пор остается неизвестной.
Хотя колюшки встречаются во многих местах вдоль побережья Северного полушария и, таким образом, рассматриваются МСОП как виды, вызывающие наименьшее беспокойство , уникальная эволюционная история, заключенная во многих пресноводных популяциях, указывает на то, что может потребоваться дополнительная правовая охрана. [ 1 ]
Диета
[ редактировать ]В разных формах и стадиях жизни трехиглая колюшка может питаться донным питанием (чаще всего личинками хирономид и бокоплавами). [ 15 ] или планктонная кормушка в озерах или океане; он также может поедать наземную добычу, упавшую на поверхность. [ 16 ] Он может поедать яйца и жарить. [ 17 ]
История жизни
[ редактировать ]
Многим популяциям требуется два года, чтобы созреть, и перед смертью они переживают только один сезон размножения, а некоторым может потребоваться до трех лет, чтобы достичь зрелости. Однако некоторые пресноводные популяции и популяции крайних широт могут достичь зрелости всего за один год.
Воспроизведение
[ редактировать ]

Половое созревание зависит от температуры окружающей среды и фотопериода. [ 18 ] Более длинные и теплые дни стимулируют более яркую окраску у самцов и развитие яиц у самок.
С конца апреля самцы и самки переходят из более глубоких вод на мелководье. Там каждый самец защищает территорию , на дне которой строит гнездо. Он начинает с того, что выкапывает небольшую яму. Затем он наполняет его растительным материалом (часто нитчатыми водорослями), песком и различным мусором, который склеивает спиггином — белковым веществом, выделяемым почками. Слово «спиггин» происходит от слова «spigg» , шведского названия трехиглой колюшки. Затем он создает туннель через более или менее сферическое гнездо, энергично проплывая через него. Строительство гнезда обычно занимает 5–6 часов. [ 19 ] хотя это также может быть растянуто на несколько дней. После этого самец ухаживает за беременной самкой, которая зигзагообразным танцем проходит мимо. (В некоторых популяциях самец ведет самку к гнезду, а не танцует зигзаг. [ 20 ] ) Он приближается к самке, проплывая очень короткие расстояния влево и вправо, а затем таким же образом плывет обратно к гнезду. Если самка следует за ней, самец часто просовывает голову внутрь гнезда и может проплыть через туннель. Затем самка также плывет через туннель, где откладывает 40–300 яиц. Самец следует за ними, чтобы оплодотворить икру. Затем самку прогоняет самец. На время развития яиц самец будет прогонять других самцов и небеременных самок. Однако он может ухаживать за другими беременными самками (в одно гнездо может быть отложено более одной партии яиц). [ нужна ссылка ]
Последовательность территориального ухаживания и брачного поведения была подробно описана Нико Тинбергеном в знаковом раннем исследовании в области этологии . Тинберген показал, что красный цвет на горле территориального самца действует как простой знаковый стимул , вызывая агрессию у других самцов и привлекая самок. [ 21 ] Красная окраска также может использоваться самками как способ оценки мужских качеств. Красная окраска обусловлена каротиноидами, содержащимися в рационе рыб. Поскольку каротиноиды не могут быть синтезированы de novo , степень окраски указывает на качество самцов (способность находить пищу), при этом самцы более высокого качества демонстрируют более интенсивную окраску. Кроме того, самцы, у которых меньше паразитов, имеют более яркий красный цвет. Многие исследования показали, что самки предпочитают самцов с более яркой красной окраской. [ 22 ] [ 23 ] [ 24 ] [ 25 ] Однако реакция на красный цвет не универсальна для всех видов. [ 26 ] [ 27 ] с чернозобыми популяциями, часто встречающимися в водах, загрязненных торфом.
Самец заботится о развивающихся икрах, обмахивая их веером. Он примыкает ко входу в туннель гнезда и плывет на месте. Движение его грудных плавников создает ток воды через гнездо, доставляя к яйцам свежую (хорошо насыщенную кислородом) воду. Он делает это не только днем, но и ночью. [ 28 ] Уровень раздувания имеет тенденцию повышаться до момента вылупления яиц, что занимает 7–8 дней при температуре 18–20 °C. Уровень раздувания также увеличивается, когда вода плохо насыщена кислородом. [ 29 ] Ближе к концу фазы развития яйца самец часто проделывает отверстия в крыше и возле края гнезда, предположительно для улучшения вентиляции гнезда во время обмахивания веером в то время, когда яйца более метаболически активны. После вылупления детенышей самец пытается удержать их вместе в течение нескольких дней, засасывая в рот всех странников и выплевывая их обратно в гнездо. После этого птенцы расходятся, и самец либо покидает гнездо, либо ремонтирует его, готовясь к следующему циклу размножения.
В Новой Шотландии форма трехиглой колюшки отходит от обычного образа родительской заботы. В отличие от других колюшек, гнездящихся на субстрате, самцы новошотландской колюшки строят гнезда в матах нитчатых водорослей. Удивительно, но почти сразу после оплодотворения самцы выгоняют икру из гнезда и возобновляют вымогательство у самок за яйцами. Следовательно, по-видимому, в этой группе населения произошла потеря родительской заботы. Поскольку у этих самцов снижена пигментация спины, в результате чего они стали перламутрово-белыми, их прозвали «белыми колюшками». В настоящее время неизвестно, являются ли они отдельным видом или просто разновидностью обыкновенной атлантической колюшки. [ 30 ] [ 31 ] [ 32 ]
Поскольку цикл размножения трехиглой колюшки зависит от света и температуры, размножением также можно манипулировать в лаборатории. Например, при правильных условиях можно стимулировать размножение колюшек дважды в календарный год, а не один раз. [ 33 ] Это может быть полезно для генетических и поведенческих исследований нескольких поколений.
Заражение цестодным паразитом Schistocephalus Solidus может вызвать снижение массы яиц или полное отсутствие яиц у самок трехиглой колюшки. [ 34 ] [ 35 ] [ 36 ]
Кооперативное поведение
[ редактировать ]Некоторые данные указывают на существование кооперативного поведения среди трехиглых колюшек, в основном совместного осмотра хищников . Осмотр хищников, по-видимому, позволяет получить информацию о риске, который представляет потенциальный хищник, и может сдержать нападение, причем ценой является увеличение вероятности нападения, если хищник окажется голодным.
Стратегия «Око за око»
[ редактировать ]Известно, что колюшки сотрудничают в стратегии «око за око» (TFT) при осмотре хищников. Идея TFT заключается в том, что человек сотрудничает на первом ходу, а затем делает то же, что и его противник на предыдущем ходу. Это позволяет сочетать совместные (начинается с сотрудничества), ответные (наказание за дезертирство) и прощающие (реагировать на сотрудничество других, даже если они ранее сбежали) поведенческие реакции. [ 37 ] Когда трехиглой колюшке, приближающейся к живому хищнику, предоставляли либо симулированного сотрудничающего компаньона, либо симулированного отступника, рыба вела себя в соответствии со стратегией «око за око», подтверждая гипотезу о том, что сотрудничество может развиваться среди эгоистов. [ 38 ]
Обычно колюшки действуют парами. У особей есть партнеры, с которыми они неоднократно совершают попарные инспекционные визиты хищников. Две реципрокные пары в каждом испытании встречаются значительно чаще, чем ожидалось, из-за случайности. Эти результаты дают дополнительные доказательства в пользу стратегии взаимного сотрудничества в борьбе с колюшками. [ 39 ]
Поведение колюшки часто называют архетипическим примером сотрудничества во время осмотра хищников. Рыбы из трех участков, различающихся по риску нападения хищников, осматривали модельного хищника парами и отвечали взаимностью как на совместные действия, так и на отступничество партнера, но не при каждой возможности. [ 40 ] Колюшки, обитающие в двух участках, где обитают рыбоядные рыбы, с большей вероятностью ответят взаимностью после совместного действия, чем после бегства. Лица из мест повышенного риска, как правило, были более склонны к сотрудничеству. [ 40 ] Лица, сопровождаемые компаньоном-моделью, примерно в трети случаев демонстрируют взаимные действия сотрудничества и отступления в ответ на движения модели. Оба примера поведения колюшки демонстрируют элементы стратегии сотрудничества, которая может напоминать «око за око». [ 40 ]
Партнерская зависимость
[ редактировать ]Стратегия взаимного сотрудничества, как было показано, проявляется и в случае с колюшками. Кроме того, размер рыбы-партнера колюшки также может быть фактором, определяющим, что будет делать колюшка, когда обе рыбы столкнутся с хищником. Две колюшки, одновременно представленные радужной форели , хищнику, гораздо большему по размеру, будут подвергаться разному риску нападения. Обычно более крупный из двух колюшек подвергается более высокому риску нападения. [ 41 ] Отдельные колюшки с большей вероятностью подойдут ближе к форели (или какому-либо другому хищнику), когда более крупный потенциальный партнер приближается к форели, чем когда к форели приближается более мелкий партнер. [ 41 ] Хотя и крупные, и мелкие партнеры ведут себя одинаково, поведение мелкого партнера больше влияет на стратегию подопытной рыбы, чем поведение крупного партнера. [ 41 ] Независимо от того, в одиночку или с партнером, который сотрудничает, более крупная рыба приблизится к хищнику ближе, чем более мелкая. [ 41 ] Если партнер отказывает, то фактор состояния колюшки (т.е. ее способность убегать) определяет, насколько близко она приближается к хищнику, а не размер колюшки. [ 41 ] Как стратегия, так и реакция на партнеров разного размера, похоже, зависят от того, сотрудничает ли партнер или отказывается.
Паразиты
[ редактировать ]
Трехиглая колюшка — вторичный промежуточный хозяин гермафродитного паразита Schistocephalus Solidus — ленточного червя рыб и рыбоядных птиц. В колюшек ленточный червь переходит через своих первых промежуточных хозяев — веслоногих циклопоидов, когда они поедаются рыбой. В брюшке колюшки паразит созревает до третьей личиночной стадии — плероцеркоида. Зараженные колюшки впоследствии поедаются рыбоядными птицами, которые служат окончательным хозяином ленточного червя. [ 42 ] [ 43 ]
Другой распространенный паразит трехиглой колюшки — микроспоридий Glugea anomala . [ 44 ] Естественно, инфицированные G. anomala теряют вес по сравнению с неинфицированными людьми. [ 45 ] но не вызывают различий в размерах между особями. [ 44 ] Glugea anomala также коррелирует с поведенческими изменениями, такими как увеличение обмеления, [ 45 ] повышается общительность и активность, снижается смелость. [ 44 ] [ 46 ] Неизвестно, обусловлены ли эти различия в поведении определенными чертами личности, предрасполагающими людей к инфекциям, или же инфекции меняют поведение .
Генетика
[ редактировать ]Трехиглая колюшка в последнее время стала основным исследовательским организмом для биологов-эволюционистов, пытающихся понять генетические изменения, связанные с адаптацией к новой среде обитания. Полный геном самки рыбы из озера Медвежья Лапа на Аляске был недавно секвенирован Институтом Броуда , и теперь доступны многие другие генетические ресурсы. [ 47 ] Эта популяция находится под угрозой из-за присутствия интродуцированной северной щуки в близлежащем озере. Трехиглую колюшку также используют для исследования экспрессии генов в мозге в зависимости от пола. Родители, подвергшиеся воздействию моделей хищников, произвели потомство с различной экспрессией генов по сравнению с теми, которые не подвергались воздействию хищников. На неперекрывающиеся гены, по-видимому, сильно влияет пол родителя, причем гены по-разному экспрессируются у потомства в зависимости от того, подвергался ли родитель мужского или женского пола воздействию хищников. [ 48 ]
Экоэволюционная динамика
[ редактировать ]Исследования трехиглой колюшки занимают центральное место в области экоэволюционной динамики. [ 49 ] [ 50 ] Экоэволюционная динамика — это область исследования, изучающая, как экологические процессы (например, динамика популяций , взаимодействие сообществ и круговорот питательных веществ ) влияют на эволюцию популяций и, в свою очередь, как эти модели эволюции влияют на экологические процессы. [ 51 ] [ 52 ] Важно отметить, что эта динамика возникает, когда существенные эволюционные изменения происходят в том же временном масштабе, что и экологические изменения (т.е. менее 1000 поколений). [ 51 ] [ 53 ] [ 54 ] Трехиглая колюшка особенно полезна для изучения эколого-эволюционной динамики, поскольку после колонизации новых сред многочисленные популяции развивались быстро и по предсказуемым, повторяющимся закономерностям. [ 49 ] [ 55 ] Эти повторяющиеся закономерности эволюции позволяют ученым оценить, воспроизводимо ли воздействие эволюции колюшки на экологические процессы.
Эколого-эволюционная основа использовалась для изучения множества аспектов биологии колюшки. Примечательно, что это исследование было сосредоточено на том, как популяции трехиглой колюшки разошлись, занимая разные экологические ниши (процесс, называемый адаптивной радиацией ), и как колюшки эволюционировали вместе со своими паразитами. [ 49 ] [ 50 ]
Экоэволюционная динамика адаптивной радиации
[ редактировать ]Большинство исследований эколого-эволюционной динамики колюшек сосредоточено на том, как адаптивная радиация различных экотипов влияет на экологические процессы. [ 49 ] [ 50 ] [ 56 ] [ 57 ] [ 58 ] Экотипы представляют собой генетически и морфологически узнаваемые популяции, занимающие четкие экологические ниши . [ 55 ] [ 56 ] [ 57 ] [ 59 ] [ 60 ] У трехиглой колюшки дивергентные экотипы часто встречаются в виде симпатрических (т. е. совместно встречающихся) или парапатрических (т. е. частично перекрывающихся, но в основном изолированных) пар видов, в том числе бентосно - лимнетических пар, [ 56 ] пресноводные — анадромные пары, [ 59 ] и пары озеро-ручей. [ 57 ] [ 60 ] Пары экотипов колюшки разошлись во времени от 10 000 лет до нескольких десятилетий назад. [ 55 ] [ 59 ]
Различные комбинации экотипов колюшки по-разному влияют на экосистемные процессы. Например, совместное присутствие специализированных донных и пресноводных колюшек оказывает иное влияние на разнообразие и численность видов-жертв по сравнению с присутствием только универсального предкового экотипа колюшки. [ 56 ] Примечательно, что этот эффект, по-видимому, обусловлен специализацией пресноводных колюшек на добыче зоопланктона , а не увеличением числа одновременно встречающихся видов колюшек. [ 56 ] Воздействие экотипической специализации на сообщества-жертвы может даже повлиять на численность водорослей и цианобактерий , которые не взаимодействуют напрямую с колюшками, а также на аспекты абиотической среды. [ 56 ] [ 57 ] например, количество окружающего света, доступного для фотосинтеза [ 56 ] и уровень растворенного кислорода, [ 56 ] углерод, [ 58 ] и фосфор. [ 57 ] Эти разнообразные изменения в экосистемных процессах могут сохраняться и влиять на естественный отбор в последующих поколениях колюшки. [ 57 ] потенциально формируя то, как популяции колюшки будут развиваться в будущем. Поскольку присутствие специализированных, а не универсальных экотипов может влиять на экосистемы таким образом, что, в свою очередь, влияет на отбор будущих поколений колюшки, адаптивное излучение специализированных экотипов может запускать экоэволюционные петли обратной связи в природных популяциях. [ 57 ]
Эколого-эволюционная динамика взаимоотношений хозяина и паразита.
[ редактировать ]Колюшки также изучались с целью изучения экоэволюционной динамики коэволюции хозяина и паразита . [ 61 ] [ 62 ] [ 63 ] Трехиглая колюшка может быть хозяином различных паразитов (например, Schistocephalus Solidus , обычного цепня рыб и рыбоядных птиц). [ 42 ] ). На разнообразие видов паразитов внутри отдельной колюшки влияют пищевая ниша человека и иммунный ответ. [ 63 ] Эта ковариация между заражением паразитами и характеристиками хозяина, вероятно, является следствием экоэволюционной обратной связи, в результате которой эволюция особенностей питания и устойчивости к паразитам у колюшек изменяет размножение паразитов и уровень заражения, что, в свою очередь, влияет на воздействие паразитов и отбор на устойчивость к паразитам у колюшек. [ 62 ] Эти обратные связи могут также выходить за рамки взаимодействия колюшки и паразита и изменять экосистемные процессы. [ 61 ] В частности, различия в устойчивости и уровне заражения среди экотипов колюшки могут изменить то, как колюшки влияют на численность видов добычи и уровень растворенных питательных веществ и кислорода. [ 61 ] Эти воздействия на экосистему могут дополнительно повлиять на отбор колюшек в последующих поколениях, что предполагает сложную петлю обратной связи между эволюцией взаимодействия хозяина и паразита, составом сообщества и абиотическими условиями. [ 61 ]
Общие методы
[ редактировать ]Многие исследователи использовали эксперименты на мезокосме , чтобы проверить, как адаптивное излучение экотипов колюшки и взаимодействие колюшки и паразита могут повлиять на экологические процессы. [ 49 ] [ 56 ] [ 57 ] [ 58 ] [ 61 ] В этих экспериментах исследователи моделируют естественную среду обитания колюшек в закрытых резервуарах, включая естественные сообщества растений и беспозвоночных, а также пресноводные экологические зоны . [ 56 ] [ 57 ] [ 58 ] [ 61 ] Затем они систематически манипулировали независимой переменной (например, какие экотипы колюшки присутствовали или наличие паразитов) и измеряли различия в биотических и абиотических аспектах экосистем при различных обработках колюшки. [ 50 ] [ 57 ] [ 58 ] [ 61 ] В некоторых случаях исследователи затем проверяли потенциальные петли обратной связи между эволюцией экотипов и экологическими изменениями, удаляя взрослых колюшек из мезокосмов и заменяя их молодью разных экотипов. [ 57 ] [ 61 ] Таким образом, исследователи смогут затем измерить, как воздействие взрослых колюшек на их экосистемы влияет на общую приспособленность молоди (например, на выживаемость и темпы роста) и на различия в приспособленности между молодью разных экотипов. [ 57 ] [ 61 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б ПриродаСерв. (2019). «Gasterosteus aculeatus» . Красный список исчезающих видов МСОП . 2019 : e.T8951A58295405. doi : 10.2305/IUCN.UK.2019-2.RLTS.T8951A58295405.en . Проверено 28 ноября 2022 г.
- ^ Фанг, Бохао; Мерила, Юха; Рибейро, Филипе; Александр, Карлос М.; Момильяно, Паоло (2018). «Всемирная филогения трехиглой колюшки» . Молекулярная филогенетика и эволюция . 127 : 613–625. дои : 10.1016/j.ympev.2018.06.008 . ПМИД 29906607 . S2CID 49231567 .
- ^ Перейти обратно: а б Фрёзе, Райнер; Поли, Дэниел (ред.) (2022). « Gasterosteus aculeatus » в FishBase . Версия от июня 2022 г.
- ^ Барбер, Иэн (2013). «Колюшки как модельные хозяева в экологической и эволюционной паразитологии». Тенденции в паразитологии . 29 (11): 556–566. дои : 10.1016/j.pt.2013.09.004 . ПМИД 24145060 .
- ^ Реймхен, Т.Е. (1989). «Потеря брачного окраса у трехиглой колюшки ( Gasterosteus aculeatus )». Эволюция . 43 (2): 450–460. дои : 10.2307/2409219 . JSTOR 2409219 . ПМИД 28568546 .
- ^ Хаглунд, ТР; Бут, Д.Г.; Блоу, DM (1990). «Аллозимная вариация и признание «белой колюшки» ». Биохимическая систематика и экология . 18 (7–8): 559–563. дои : 10.1016/0305-1978(90)90129-4 .
- ^ Нур, Мохамед А.Ф. (1999). «Подкрепление и другие последствия симпатии» . Наследственность . 83 (5). Общество генетиков ( Природа ): 503–508. дои : 10.1038/sj.hdy.6886320 . ISSN 0018-067X . ПМИД 10620021 . ( ОРЦИД : 0000-0002-5400-4408 . GS : 5nkhrpUAAAAJ ).
- ^ «Рыбы Дарвина: трехиглая колюшка северо-запада Тихого океана» . Сиэтл Таймс . 15 февраля 2009 г. Проверено 11 июня 2022 г.
- ^ «Рыба озера Вашингтон, эволюционирующая в обратном направлении, дает ключ к разгадке генетики» . Сиэтл Таймс . 15 мая 2008 г. Проверено 11 июня 2022 г.
- ^ Китано, Дж. (15 мая 2008 г.). «Обратная эволюция панцирных пластин трехиглой колюшки» (PDF) . Современная биология . 18 (10): 769–774. дои : 10.1016/j.cub.2008.04.027 . ПМИД 18485710 . S2CID 7864384 . Архивировано (PDF) из оригинала 12 августа 2017 г. Проверено 11 июня 2022 г.
- ^ «Музей Берка и другие отмечают 200-летие Чарльза Дарвина» . Новости УВ . Проверено 11 июня 2022 г.
- ^ Бем, Дж. Э.; Айвс, Арканзас; Бофман, JW (2010). «Нарушение изоляции после спаривания и распад пары видов в результате гибридизации». Американский натуралист . 175 (1): 11–26. дои : 10.1086/648559 . ПМИД 19916869 . S2CID 15817509 .
- ^ Канада – Закон о видах, находящихся под угрозой . dfo-mpo.gc.ca
- ^ Кэрролл, Шон Б. (2006). Создание наиболее приспособленных: ДНК и окончательная судебно-медицинская экспертиза эволюции . WW Norton & Co., стр. 56–57. ISBN 978-0-393-06163-5 .
- ^ «Gasterosteus aculeatus» . Сеть разнообразия животных .
- ^ Санчес-Эрнандес, Дж. (2012). «Применение анализа биологических признаков («признаков») добычи для изучения пищевого поведения рыб-бентофагов: на примере колюшки ( Gasterosteus Gymnurus Cuvier 1829)». Лимнетика . 31 (1): 59–76.
- ^ Вориски, ФГ; Фитцджеральд, Дж.Дж. (1985). «Секс, каннибализм и колюшки» . Поведенческая экология и социобиология . 18 (1): 15–18. дои : 10.1007/BF00299233 . S2CID 21522305 .
- ^ О'Брайен, Конор С.; Бурдо, Райан; Брэдшоу, Уильям Э.; Хольцапфель, Кристина М.; Креско, Уильям А. (2012). «Сохранение фотопериодической нейроэндокринной оси у позвоночных: данные костистых рыб Gasterosteus aculeatus » . Общая и сравнительная эндокринология . 178 (1): 19–27. дои : 10.1016/j.ygcen.2012.03.010 . ПМЦ 3389224 . ПМИД 22504272 .
- ^ ван Иерсель, JJA (1953). «Анализ родительского поведения самца трехиглой колюшки ( Gasterosteus aculeatus L.)». Дополнение к поведению . 3 (3): 1–159. JSTOR 30039128 .
- ^ Кандолин, У.; Фойгт, Х.-Р. (2001). «Отсутствие влияния паразита на размножение самцов колюшки: лабораторный артефакт?». Паразитология . 122 (4): 457–464. дои : 10.1017/S0031182001007600 . ПМИД 11315179 . S2CID 15544990 .
- ^ Тинберген, Нико (1989). Исследование инстинкта . Оксфорд [Англия]. ISBN 978-0198577225 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Милински, М.; Баккер, ТКМ (1990). «Самки колюшек используют окраску самцов при выборе партнера и, следовательно, избегают самцов, зараженных паразитами» (PDF) . Природа . 344 (6264): 330–333. Бибкод : 1990Natur.344..330M . дои : 10.1038/344330a0 . S2CID 4322443 .
- ^ МакЛеннан, округ Колумбия; Макфэйл, доктор юридических наук (1990). «Экспериментальные исследования эволюционного значения половой диморфной брачной окраски у Gasterosteus aculeatus (L.): взаимосвязь между цветом самца и поведением самки». Канадский журнал зоологии . 68 (3): 482–492. дои : 10.1139/z90-071 .
- ^ Баккер, TCM; Мундвайлер, Б. (1994). «Выбор самки и красная окраска самца в естественной популяции трехиглой колюшки ( Gasterosteus aculeatus )» (PDF) . Поведенческая экология . 5 : 74–80. дои : 10.1093/beheco/5.1.74 .
- ^ Баубе, CL; Роуленд, WJ; Фаулер, Дж. Б. (1995). «Механизмы выбора партнера по цвету у самок трехиглой колюшки: оттенок, контраст и конфигурационные сигналы». Поведение . 132 (13/14): 979–996. дои : 10.1163/156853995x00405 . JSTOR 4535315 .
- ^ Маккиннон, Дж. С. (1995). «Видеобрачные предпочтения самок трехиглой колюшки из популяций с разной окраской самцов». Поведение животных . 50 (6): 1645–1655. дои : 10.1016/0003-3472(95)80018-2 . S2CID 53193561 .
- ^ Брейтуэйт, Виктория А.; Барбер, Иэн (2000). «Ограничения сексуальных предпочтений на основе цвета у трехиглой колюшки ( Gasterosteus aculeatus )». Поведенческая экология и социобиология . 47 (6): 413–416. дои : 10.1007/s002650050684 . S2CID 28383103 .
- ^ Рибс, С.Г.; Вориски-младший, ФГ; Фицджеральд, Дж.Дж. (1984). «Диельские закономерности обмахивания, дыхания яиц и ночного поведения самцов трехиглой колюшки Gasterosteus aculeatus L. (f. trachurus )». Канадский журнал зоологии . 62 (3): 329–334. дои : 10.1139/z84-051 .
- ^ Севентер, П. (1961). «Причинно-следственное исследование смещенной активности (раздувание в Gasterosteus aculeatus L.)». Дополнение к поведению . 9 : 1–170.
- ^ Макдональд, Дж. Ф.; Беккерс, Дж.; МакИсаак, С.М.; Блоу, DM (1995). «Приливное размножение и воздушное развитие эмбрионов колюшки (Gasterosteus)». Поведение . 132 (15): 1183–1206. дои : 10.1163/156853995x00522 . JSTOR 4535330 .
- ^ Макдональд, Дж. Ф.; МакИсаак, С.М.; Беккерс, Дж.; Блоу, DM (1995). «Эксперименты по выживанию эмбрионов, выбору среды обитания и конкурентоспособности колюшки (Gasterosteus), гнездящейся в каменистой приливной зоне». Поведение . 132 (15): 1207–1221. дои : 10.1163/156853995x00531 .
- ^ Джеймисон, штат Индиана; Блоу, Д.М.; Колган, PW (1992). «Полевые наблюдения за репродуктивной биологией недавно обнаруженной колюшки (Gasterosteus)». Канадский журнал зоологии . 70 (5): 1057–1063. дои : 10.1139/z92-148 .
- ^ Сэттлер, Джессика Л.; Бофман, Джанетт В. (2020). «Прогресс в разведении колюшки ( Gasterosteus aculeatus ) для получения двух репродуктивных циклов в год». Журнал биологии рыб . 97 (5): 1576–1581. дои : 10.1111/jfb.14517 . ПМИД 32869321 . S2CID 221402138 .
- ^ Хейнс, Дэвид С.; Бейкер, Джон А. (февраль 2003 г.). «Уменьшение размера яиц в природных популяциях трехиглой колюшки, зараженных цестодным макропаразитом». Журнал паразитологии . 89 (1): 1–6. doi : 10.1645/0022-3395(2003)089[0001:ROESIN]2.0.CO;2 . ПМИД 12659295 . S2CID 24098353 .
- ^ Макфэйл, доктор юридических наук; Пикок, SD (2011). «Некоторые эффекты цестоды ( Schistocephalus Solidus ) на размножение трехиглой колюшки ( Gasterosteus aculeatus ): эволюционные аспекты взаимодействия хозяина и паразита». Канадский журнал зоологии . 61 (4): 901–908. дои : 10.1139/z83-118 .
- ^ Хейнс, Дэвид С; Певица Скарлет С; Бейкер, Джон А. (1999). «Вирулентность цестоды Schistocephalus Solidus и размножение у зараженной трехиглой колюшки Gasterosteus aculeatus ». Канадский журнал зоологии . 77 (12): 1967–1974. дои : 10.1139/z99-180 .
- ^ Николас Б. Дэвис; Джон Р. Кребс; Стюарт А. Уэст (2 апреля 2012 г.). Введение в поведенческую экологию (4-е изд.). Оксфорд: Уайли-Блэквелл. ISBN 978-1-4051-1416-5 .
- ^ Милински, Манфред (1987). «ОТО ЗА ТО в колюшках и эволюция сотрудничества». Природа . 325 (29): 433–435. Бибкод : 1987Natur.325..433M . дои : 10.1038/325433a0 . ПМИД 3808044 . S2CID 4320531 .
- ^ Милински, Манфред; Д. Пфлюгер; Д. Куллинг; Р. Кеттлер (1990). «Колюшки неоднократно взаимодействуют в взаимных парах?». Поведенческая экология и социобиология . 27 (1): 17–21. дои : 10.1007/bf00183308 . S2CID 27982566 .
- ^ Перейти обратно: а б с Хантингфорд, Фелисити; Иоанн Лазарь; Брайан Барри; Салли Уэбб (1994). «Динамический анализ совместной проверки хищниками колюшек». Поведение животных . 47 (2): 413–423. дои : 10.1006/anbe.1994.1055 . S2CID 53156278 .
- ^ Перейти обратно: а б с д и Куллинг, Дэвид; Манфред Милински (1992). «Риск нападения хищников в зависимости от размера и качество партнера при осмотре колюшек хищниками». Поведение животных . 44 (5): 949–955. дои : 10.1016/s0003-3472(05)80590-1 . S2CID 53161426 .
- ^ Перейти обратно: а б ЛоБуэ, КП; Белл, Массачусетс (1993). «Фенотипические манипуляции цестодного паразита Schistocephalus Solidus с его промежуточным хозяином, Gasterosteus aculeatus , трехиглой колюшкой». Американский натуралист . 142 (4): 725–735. дои : 10.1086/285568 .
- ^ Дубинина, Миннесота (1980). Ленточные черви (Cestoda, Ligulidae) фауны СССР .
- ^ Перейти обратно: а б с Петкова И.; Эбби-Ли, RN; Лёвли, Х. (2018). «Паразитарная инфекция и личность хозяина: Glugea трехиглая колюшка, зараженная , более социальна» . Поведенческая экология и социобиология . 72 (11). дои : 10.1007/s00265-018-2586-3 . ПМК 6182751 .
- ^ Перейти обратно: а б Уорд, AJW; Дафф, Эй Джей; Краузе, Дж.; Барбер, И. (2005). «Стайное поведение колюшек, зараженных микроспоридийным паразитом Glugea anomala ». Экологическая биология рыб . 72 (2): 155–160. дои : 10.1007/s10641-004-9078-1 .
- ^ Эбби-Ли, RN; Крещенко А.; Фернандес Сала, X.; Петкова И.; Лёвли, Х. (2019). «Влияние манипуляций с моноамином на личность и экспрессию генов трехиглой колюшки». Журнал экспериментальной биологии . 222 (20). дои : 10.1242/jeb.211888 .
- ^ «Геном колюшки в ЭНСЕМБЛ» .
- ^ Хеллманн, Дженнифер; Бухари, Сайед; Дено, Джек; Белл, Элисон (15 ноября 2020 г.). «Полоспецифическая пластичность в поколениях I: материнское и отцовское влияние на сыновей и дочерей» . Журнал экологии животных . 89 (12): 2788–2799. дои : 10.1111/1365-2656.13364 . ПМЦ 7902357 . ПМИД 33191518 .
- ^ Перейти обратно: а б с д и Хендри, А. П., К. Л. Пайчел, Б. Мэтьюз, Дж. В. Бофман и П. Носил. 2013. Исследование колюшки: настоящее и будущее. Исследования в области эволюционной экологии 15:111–141.
- ^ Перейти обратно: а б с д Беккерман, А. П., Д. З. Чайлдс и А. О. Бергланд. 2016. Экоэволюционная биология: петли питания и обратной связи. Текущая биология 26: R161–R164.
- ^ Перейти обратно: а б Шонер, Т.В. 2011. Новейший синтез: понимание экологической динамики. Наука 331: 426–429.
- ^ Бруннер, Ф.С., Дж. А. Дир, М. Эгас, К. Эйсагирре и Дж. Э. Реймакерс. 2019. Разнообразие эколого-эволюционной динамики: сравнение обратных связей между экологией и эволюцией в разных масштабах. Функциональная экология 33:7–12.
- ^ Фуссманн, Г. Ф., М. Лоро и П. А. Абрамс. 2007. Эколого-эволюционная динамика сообществ и экосистем. Функциональная экология 21: 465–477.
- ^ Эллнер, SP 2013. Быстрая эволюция: от генов к сообществам и обратно? Функциональная экология 27: 1087–1099.
- ^ Перейти обратно: а б с Белл, Массачусетс, и МЫ Агирре. 2013. Современная эволюция, аллельный рециклинг и адаптивная радиация трехиглой колюшки. Исследования в области эволюционной экологии 15:377–411.
- ^ Перейти обратно: а б с д и ж г час я дж Хармон, Л. Дж., Б. Мэтьюз, С. Де Рош, Дж. М. Чейз, Дж. Б. Шурин и Д. Шлютер. 2009. Эволюционная диверсификация колюшки влияет на функционирование экосистемы. Природа 458: 1167–1170.
- ^ Перейти обратно: а б с д и ж г час я дж к л Мэтьюз Б., Т. Эбишер, К.Е. Саллам, Б. Лундсгаард-Хансен и О. Зеехаузен. 2016. Экспериментальные доказательства экоэволюционной обратной связи во время адаптивной дивергенции. Текущая биология 26: 483–489.
- ^ Перейти обратно: а б с д и Рудман С.М. и Д. Шлютер. 2016. Экологические последствия обратного видообразования трехиглой колюшки. Текущая биология 26: 490–495.
- ^ Перейти обратно: а б с Фур, К.Г., Ф.А. фон Хиппель и М.А. Белл. 2012. Частичная репродуктивная изоляция недавно выведенной резидентной пресноводной популяции трехвидной колюшки (Gasterosteus aculeatus) от ее предполагаемого анадромного предка. Эволюция 66:3277–3286.
- ^ Перейти обратно: а б Бернер Д., М. Рёсти, А. П. Хендри и В. Зальцбургер. 2010. Ограничения видообразования, предложенные путем сравнения дивергенции озерно-ручьевой колюшки на двух континентах. Молекулярная экология 19: 4963–4978.
- ^ Перейти обратно: а б с д и ж г час я Бруннер, Ф.С., Дж. М. Анайя-Рохас, Б. Мэтьюз и К. Эйсагирре. 2017. Экспериментальные доказательства того, что паразиты вызывают экоэволюционную обратную связь. Труды Национальной академии наук Соединенных Штатов Америки 114: 3678–3683.
- ^ Перейти обратно: а б Вебер, Дж. Н., М. Кальбе, К. К. Шим, Н. И. Эрин, Н. К. Стайнель, Л. Ма и Д. И. Болник. 2017. Сопротивляйтесь глобально, заражайте локально: трансконтинентальный тест адаптации колюшки и ее паразита-ленточного червя. Американский натуралист 189:43–57.
- ^ Перейти обратно: а б Больник, Д.И., Э.Дж. Ресетаритс, К. Балларе, Ю.Е. Стюарт и В.Е. Штутц. 2020. Зависимое от масштаба влияние особенностей участка хозяина на видовой состав метасообщества паразитов колюшки. Экология 101:1–16.
Дальнейшее чтение
[ редактировать ]- Вуттон, Р.Дж. 1976. Биология колюшек. Академическое издательство, Лондон.
- Белл, М.А. и Фостер, С.А. (ред.) 1994. Эволюционная биология трехиглой колюшки. Издательство Оксфордского университета, Нью-Йорк.
- Остлунд-Нильссон С., Майер И. и Хантингфорд Ф.А. (ред.) 2007. Биология трехиглой колюшки. CRC Press, Бока-Ратон.
Внешние ссылки
[ редактировать ]- «Gasterosteus aculeatus» . Интегрированная таксономическая информационная система . Проверено 19 марта 2006 г.
- Раздел Канадского закона о видах, находящихся под угрозой, посвященный парам видов.
- Посмотреть геном колюшки в Ensembl
| Базы данных органов управления : Национальные |
|---|
- Виды, находящиеся в Красном списке МСОП, вызывающие наименьшее беспокойство
- Гастеростеус
- Рыбы северной части Тихого океана
- Рыба Северной Атлантики
- Рыба Великих озер
- Пресноводные рыбы Европы
- Фауна района залива Сан-Франциско
- Прибрежная фауна западной части Северной Америки
- Фауна Атлантической Канады
- Фауна северо-востока США
- Рыба описана в 1758 году.
- Таксоны, названные Карлом Линнеем
- Рыба Балтийского моря