Анатомия рыбы
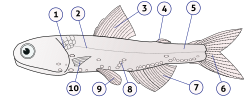

Анатомия рыб это изучение формы или морфологии рыб — . Ее можно противопоставить физиологии рыб , которая изучает, как составные части рыбы функционируют вместе в живой рыбе. [1] На практике анатомия и физиология рыб дополняют друг друга: первая касается структуры рыбы, ее органов или составных частей, а также того, как они соединяются друг с другом, что можно наблюдать на препаровальном столе или под микроскопом, а последний касается того, как эти компоненты функционируют вместе в живой рыбе.
Анатомия рыб часто определяется физическими характеристиками воды — среды, в которой живут рыбы. Вода намного плотнее рыбы, содержит относительно небольшое количество растворенного кислорода и поглощает больше света, чем воздух. Тело рыбы разделено на голову, туловище и хвост, хотя разделения между ними не всегда заметны внешне. Скелет, образующий опорную структуру внутри рыбы, состоит либо из хрящей ( хрящевые рыбы ), либо из костей ( костные рыбы ). Основным элементом скелета является позвоночный столб , состоящий из сочленяющихся позвонков , легких, но прочных. Ребра прикрепляются к позвоночнику, конечностей и поясов конечностей нет. Основные внешние особенности рыбы — плавники — состоят либо из костных, либо из мягких шипов, называемых лучами , которые, за исключением хвостовых плавников , не имеют прямой связи с позвоночником. Их поддерживают мышцы, составляющие основную часть туловища. [2] Сердце имеет две камеры и перекачивает кровь через дыхательные поверхности жабр , а затем по всему телу в едином контуре кровообращения. [3] Глаза приспособлены для видения под водой и обладают только местным зрением. [ необходимо определение ] Есть внутреннее ухо , но нет наружного или среднего уха . Низкочастотные колебания улавливаются системой органов чувств боковой линии , проходящей по длине боков рыб, которая реагирует на близлежащие движения и на изменение давления воды. [2]
Акулы и скаты — базальные рыбы с многочисленными примитивными анатомическими особенностями, сходными с таковыми у древних рыб, включая хрящевой скелет. Их тела, как правило, уплощены в дорсо-вентральном направлении, обычно имеют пять пар жаберных щелей и большой рот, расположенный на нижней стороне головы. Дерма . покрыта отдельными дермальными чешуйками плакоидными У них имеется клоака , в которую открываются мочевой и половой ходы, но нет плавательного пузыря . Хрящевые рыбы откладывают небольшое количество крупных желточных икринок. Некоторые виды яйцеживородящие , у которых молодь развивается внутри, но другие яйцекладущие , и личинки развиваются снаружи в яйцеклетках . [4]
Линия костистых рыб демонстрирует более производные анатомические черты, часто с серьезными эволюционными изменениями по сравнению с чертами древних рыб. У них костный скелет, они обычно уплощены с боков, имеют пять пар жабр, защищенных крышечкой , и рот на кончике морды или рядом с ней. Дерма покрыта перекрывающимися чешуйками . У костных рыб есть плавательный пузырь, который помогает им поддерживать постоянную глубину в толще воды , но нет клоаки. Чаще всего они выметывают большое количество мелких икринок с небольшим количеством желтка, которые разбрасывают в толщу воды. [4]
Тело
[ редактировать ]
Во многих отношениях анатомия рыб отличается от анатомии млекопитающих . Тем не менее, он по-прежнему имеет тот же базовый план тела , из которого все позвоночные произошли : хорда , рудиментарные позвонки, а также четко выраженные голова и хвост. [5] [6]
Рыбы имеют множество различных строений тела. На самом широком уровне их тело разделено на голову, туловище и хвост, хотя эти подразделения не всегда видны снаружи. Тело часто веретенообразное , обтекаемое строение тела часто встречается у быстродвижущихся рыб. Некоторые виды могут быть нитевидными ( угревидными ) или червеобразными ( червеобразными ). Рыбы часто либо сжаты ( с боков тонкие и высокие ), либо вдавлены (сплющены в дорсо-вентральном направлении ).
Скелет
[ редактировать ]
Существует два разных типа скелета: экзоскелет , который представляет собой стабильную внешнюю оболочку организма, и эндоскелет , который образует опорную структуру внутри тела. Скелет рыб состоит либо из хрящей (хрящевые рыбы), либо из костей (костные рыбы). Эндоскелет рыб состоит из двух основных компонентов: осевого скелета, состоящего из черепа и позвоночного столба, и добавочного скелета, поддерживающего плавники. [7] Плавники состоят из костных плавниковых лучей и, за исключением хвостового плавника, не имеют прямой связи с позвоночником. Их поддерживают только мышцы. Ребра прикрепляются к позвоночнику.
Кости — это жесткие органы , составляющие часть эндоскелета позвоночных. Они функционируют для перемещения, поддержки и защиты различных органов тела, производства красных и белых кровяных телец и хранения минералов . Костная ткань представляет собой разновидность плотной соединительной ткани . Кости имеют разнообразную форму и сложную внутреннюю и внешнюю структуру. Они легкие, но при этом прочные и твердые, а также выполняют множество других биологических функций .
Позвонки
[ редактировать ]



Рыбы – позвоночные животные. Все позвоночные животные построены по основному строению тела хордовых : жесткий стержень, проходящий по всей длине животного (позвоночный столб или хорда), [8] с полой трубкой нервной ткани ( спинной мозг ) над ним и желудочно-кишечным трактом под ним. У всех позвоночных рот находится на переднем конце животного или прямо под ним, а анус открывается наружу перед концом тела. Оставшаяся часть тела за анусом образует хвост с позвонками и спинным мозгом, но не имеет кишечника. [9]
Определяющей характеристикой позвоночного является позвоночный столб, в котором хорда (жесткий стержень однородного состава), обнаруженный у всех хордовых, заменена сегментированной серией более жестких элементов (позвонков), разделенных подвижными суставами ( межпозвонковыми дисками) , возникшими эмбрионально. и эволюционно от хорды). Однако некоторые рыбы вторично [ нужны разъяснения ] потеряли эту анатомию, сохранив хорду и во взрослом возрасте, как, например, осетр . [10]
Позвоночный столб состоит из центра (центрального тела или позвоночника позвонка), позвоночных дуг , которые выступают сверху и снизу центра, и различных отростков , которые выступают из центра или дуг. Дуга, идущая от вершины центра, называется нервной дугой , а гемальная дуга или шеврон находится под центром в хвостовых позвонках рыб. Центр рыбы обычно вогнут на каждом конце (амфицельный), что ограничивает движение рыбы. Напротив, центр млекопитающих плоский на каждом конце (ацелозный), форма, которая может поддерживать и распределять сжимающие силы.
Позвонки лопастных рыб состоят из трех отдельных костных элементов. Дуга позвонка окружает спинной мозг и по форме во многом похожа на ту, что встречается у большинства других позвоночных. Прямо под аркой находится небольшой пластинчатый плевроцентр, который защищает верхнюю поверхность хорды. Ниже этого более крупный арочный промежуточный центр защищает нижнюю границу. Обе эти структуры встроены в единую цилиндрическую массу хряща. Подобное расположение было обнаружено у примитивных четвероногих , но в эволюционной линии, которая привела к рептилиям , млекопитающим и птицам, межцентрум частично или полностью заменился увеличенным плевроцентром, который, в свою очередь, стал костным телом позвонка. [11]
У большинства лучепёрых рыб , включая все костистые , эти две структуры слиты и встроены в твердый кусок кости, внешне напоминающий тело позвонка млекопитающих. У современных земноводных ниже дуги позвонка имеется просто цилиндрический кусок кости, без следов отдельных элементов, присутствовавших у ранних четвероногих. [11]
У хрящевых рыб, таких как акулы , позвонки состоят из двух хрящевых трубок. Верхняя трубка формируется из дуг позвонков, но включает также дополнительные хрящевые структуры, заполняющие промежутки между позвонками, заключающие спинной мозг в практически сплошную оболочку. Нижняя трубка окружает хорду и имеет сложную структуру, часто включающую несколько слоев кальцификации . [11]
У миног есть дуги позвонков, но нет ничего похожего на тела позвонков, встречающиеся у всех высших позвоночных . Даже дуги прерывистые, состоящие из отдельных кусочков дугообразных хрящей вокруг спинного мозга на большей части тела, сменяющихся на длинные полоски хряща сверху и снизу в хвостовой области. У миксин нет настоящего позвоночного столба, но в хвосте имеется несколько крошечных нервных дуг. [11] [12] Однако у миксин есть череп . По этой причине в прошлом миксины иногда исключали из состава Vertebrata и вместо этого помещали в сестринскую группу позвоночных в таксоне « Craniata ». [13] Молекулярный анализ [ указать ] с 1992 года показали, что миксины являются сестринской группой миног клады Cyclostomi и, следовательно, являются позвоночными животными в филогенетическом смысле. [14]
Голова
[ редактировать ]


Голова или череп включает крышу черепа (набор костей, покрывающих мозг, глаза и ноздри), морду (от глаза до самой передней точки верхней челюсти ), жаберную крышку или жаберную крышку (отсутствует у акул и бесчелюстная рыба ) и щека , простирающаяся от глаза до предкрышки . Жаберная крышка и предкрышка могут иметь или не иметь шипы. У акул и некоторых примитивных костистых рыб позади каждого глаза находится дыхальце — небольшое дополнительное жаберное отверстие.
Череп рыб образован из ряда слабо соединенных между собой костей. Бесчелюстные рыбы и акулы обладают только хрящевым эндокраном , причем верхняя и нижняя челюсти хрящевых рыб представляют собой отдельные элементы, не прикрепленные к черепу. Костные рыбы имеют дополнительную кожную кость , образующую более или менее целостную крышу черепа у двоякодышащих и холостовых рыб . Нижняя челюсть определяет подбородок.
У миног рот сформирован в виде ротового диска. Однако у большинства челюстных рыб существуют три основные конфигурации. Рот может находиться на переднем конце головы (конечный), может быть перевернут (верхний), может быть повернут вниз или на нижней части рыбы (субтерминальный или нижний). Рот может быть преобразован в присоску, приспособленную для цепляния за предметы в быстро движущейся воде.
Более простое строение встречается у бесчелюстных рыб, у которых череп представлен корытообразной корзиной из хрящевых элементов, лишь частично охватывающей мозг и связанной с капсулами внутреннего уха и единственной ноздрей. Примечательно, что у этих рыб нет челюстей. [15]
Хрящевые рыбы, такие как акулы, также имеют простую и, по-видимому, примитивную структуру черепа. Череп представляет собой единую структуру, образующую оболочку вокруг мозга, охватывающую нижнюю поверхность и боковые стороны, но всегда хотя бы частично открытую вверху в виде большого родничка . Самая передняя часть черепа включает переднюю хрящевую пластинку, рострум и капсулы, в которых заключены органы обоняния . За ними расположены глазницы, а затем дополнительная пара капсул, заключающих в себе структуры внутреннего уха. Наконец, череп сужается к задней части, где большое затылочное отверстие лежит непосредственно над единственным мыщелком , сочленяющимся с первым позвонком. Меньшие отверстия для черепных нервов можно найти в различных точках черепа. Челюсти состоят из отдельных хрящевых обручей, почти всегда отличных от собственно черепа. [15]
У лучепёрых рыб также произошли значительные изменения по сравнению с примитивным строением. Крыша черепа в целом хорошо сформирована, и хотя точное соотношение ее костей с костями четвероногих неясно, для удобства им обычно дают схожие названия. Однако другие элементы черепа могут быть уменьшены; за увеличенными глазницами имеется небольшая область щек, а между ними мало костей, если они вообще есть. Верхняя челюсть часто формируется в основном из предчелюстной кости , при этом сама верхняя челюсть расположена дальше назад, а дополнительная кость, симплетическая , соединяет челюсть с остальной частью черепа. [15]
Хотя черепа ископаемых лопастных рыб напоминают черепа ранних четвероногих, этого нельзя сказать о черепах современных двоякодышащих рыб. Крыша черепа сформирована не полностью и состоит из множества костей несколько неправильной формы, не имеющих прямого отношения к костям четвероногих. Верхняя челюсть образована только крыловидными костями и сошниками , все из которых несут зубы. Большая часть черепа образована хрящами, а его общая структура редуцирована. [15]
На голове может быть несколько мясистых образований, известных как усики , которые могут быть очень длинными и напоминать усы. Многие виды рыб также имеют на голове разнообразные выступы или шипы. Ноздри глубины почти всех рыб . не соединяются с ротовой полостью, а представляют собой ямки различной формы и
 Череп северной щуки
Череп северной щуки


Внешние органы
[ редактировать ]Челюсть
[ редактировать ]
Челюсти позвоночных, вероятно, первоначально развились в силурийский период и появились у рыб-плакодерм , которые в дальнейшем диверсифицировались в девонском периоде . Считается, что челюсти произошли от глоточных дуг , поддерживающих жабры рыб. Две самые передние из этих дуг, как полагают, являются самой челюстью (см. гиомандибула ) и подъязычной дугой , которая прижимает челюсть к черепной коробке и увеличивает механическую эффективность . Хотя не существует ископаемых доказательств, непосредственно подтверждающих эту теорию, она имеет смысл в свете количества глоточных дуг, видимых у современных челюстных животных ( челюстных ), имеющих семь дуг, и примитивных бесчелюстных позвоночных ( Agnatha ), которые есть девять. [ нужна ссылка ]

| Внешние видео | |
|---|---|
Считается, что первоначальное избирательное преимущество, полученное челюстями, было связано не с питанием, а с повышением эффективности дыхания . Челюсти использовались в щочном насосе (наблюдаемом у современных рыб и амфибий), который перекачивает воду через жабры рыб или воздух в легкие амфибий. С течением времени было выбрано более привычное использование челюстей при питании, которое стало очень важной функцией у позвоночных.
Системы связей широко распространены у животных. Наиболее полный обзор различных типов связей у животных дал М. Мюллер. [16] который также разработал новую систему классификации, которая особенно хорошо подходит для биологических систем. Механизмы связи особенно часты и разнообразны в голове костистых рыб, таких как губаны , у которых развилось множество специализированных механизмов водного питания . Особенно развиты рычажные механизмы выдвижения челюстей . При аспирационном питании система соединенных четырехстержневых рычагов отвечает за согласованное открывание рта и трехмерное расширение полости рта. Другие связи ответственны за выпячивание предчелюстной кости.
Глаза
[ редактировать ]
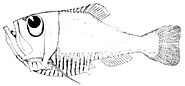
Глаза рыб похожи на глаза наземных позвоночных, таких как птицы и млекопитающие, но имеют более сферическую линзу . Их сетчатка обычно имеет как палочки, так и колбочки (для скотопического и фотопического зрения ), и большинство видов обладают цветовым зрением . Некоторые рыбы могут видеть ультрафиолет , а некоторые — поляризованный свет . Среди бесчелюстных рыб хорошо развиты глаза у миноги, а у миксины лишь примитивные глазные пятна. [17] Предки современных миксин, считавшихся протопозвоночными, [18] очевидно, были вытеснены в очень глубокие темные воды, где они были менее уязвимы для зрячих хищников и где выгодно иметь выпуклое глазное пятно, собирающее больше света, чем плоское или вогнутое. В отличие от людей, рыбы обычно регулируют фокус , перемещая хрусталик ближе к сетчатке или дальше от нее. [19]
Жабры
[ редактировать ]
Жабры, расположенные под жаберной крышкой, являются органом дыхания, предназначенным для извлечения кислорода из воды и выделения углекислого газа. Обычно их не видно, но их можно увидеть у некоторых видов, например у плащеносной акулы . Лабиринтный орган Anabantoidei Clariidae и . используется рыбами для извлечения кислорода из воздуха Жаберные тычинки представляют собой пальцеобразные выступы на жаберной дуге, которые служат фильтраторами для удержания отфильтрованной добычи. Они могут быть костными или хрящевыми.
Кожа
[ редактировать ]Кожа рыб является частью покровной системы , состоящей из двух слоев: эпидермиса и слоя дермы. Эпидермис количеством происходит из эктодермы и становится самым поверхностным слоем, полностью состоящим из живых клеток с минимальным кератина . Обычно он проницаем. Дерма мезодермы происходит из обнаруженных и напоминает небольшую соединительную ткань , состоящую в основном из коллагеновых волокон, у костистых рыб. У некоторых видов рыб чешуя выходит из дермы, проникает в тонкий слой базальной мембраны , лежащей между эпидермисом и дермой, становится видимой снаружи и покрывает слой эпидермиса. [20]
Как правило, кожа также содержит потовые и сальные железы , которые уникальны для млекопитающих, но у рыб встречаются и другие типы кожных желез. В эпидермисе рыбы обычно имеют множество отдельных клеток кожи, секретирующих слизь, называемых бокаловидными клетками , которые вырабатывают слизистое вещество на поверхности кожи. Это способствует изоляции и защите от бактериальной инфекции. [21] [22] Цвет кожи многих млекопитающих часто обусловлен меланином , содержащимся в их эпидермисе. Однако у рыб окраска кожи во многом обусловлена хроматофорами в дерме, которые, помимо меланина, могут содержать гуаниновые или каротиноидные пигменты. Многие виды, например камбала , меняют цвет своей кожи, регулируя относительный размер хроматофоров. У некоторых рыб также могут быть ядовитые железы, фотофоры или клетки, которые производят более водянистую серозную жидкость в дерме. [23] [21] [24]

Весы
[ редактировать ]Также частью покровной системы рыб являются чешуи, покрывающие внешнюю часть тела многих челюстных рыб. Общеизвестные чешуйки происходят из дермы или мезодермы и могут быть похожи по структуре на зубы. Вместо этого некоторые виды покрыты щитками . У других может не быть чешуи, покрывающей внешнее тело.
 Сингулярная циклоидная шкала боуфина
Сингулярная циклоидная шкала боуфина Циклоидная чешуя, покрывающая роху
Циклоидная чешуя, покрывающая роху Циклоидные чешуи Боуфина
Циклоидные чешуи Боуфина



Существует четыре основных типа рыбьей чешуи, происходящей из дермы. [25] [26]
- Плакоидные чешуйки , также называемые дермальными зубчиками, представляют собой заостренные чешуйки. По строению они похожи на зубы , у которых они состоят из дентина и покрыты эмалью . Они типичны для хрящевых рыб (правда, у химер они есть только на ластиках).
- Ганоидные чешуйки представляют собой плоские чешуйки, имеющие вид основания. Производные из плакоидных чешуек, они имеют толстый слой эмали, но без подстилающего слоя дентина. Эти чешуи покрывают тело рыбы, слегка перекрывая друг друга. Они типичны для гара и бичира .
- Циклоидные чешуи — это маленькие чешуйки овальной формы с годичными кольцами, напоминающими кольца дерева. У них отсутствуют эмаль, дентин и сосудистый костный слой. Боуфин и ремора имеют циклоидную чешую.
- Ктеноидные чешуи похожи на циклоидные, также имеют годичные кольца, лишены эмали, дентина и сосудистого костного слоя. Их отличают шипы или выступы по одному краю. У палтуса есть чешуя такого типа.
 Рыбья чешуя: 1. циклоидная чешуя; 2. ктеноидная чешуя; 3. плаккоидная чешуя; 4. ганоидная шкала
Рыбья чешуя: 1. циклоидная чешуя; 2. ктеноидная чешуя; 3. плаккоидная чешуя; 4. ганоидная шкала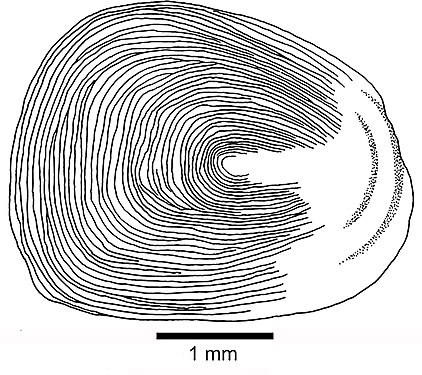 Циклоидная шкала
Циклоидная шкала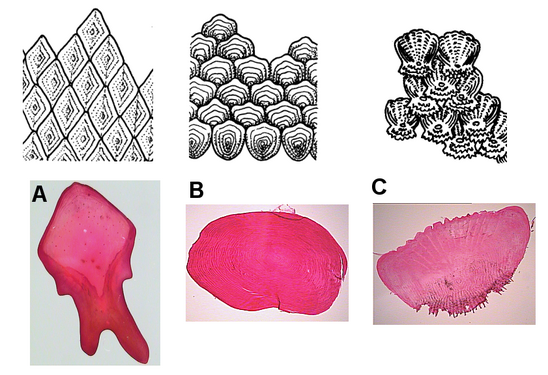 Рыбья чешуя: A. ganoid; Б. циклоидная; С. ктеноид
Рыбья чешуя: A. ganoid; Б. циклоидная; С. ктеноид



Другой менее распространенный тип чешуи — щиток , который может представлять собой внешнюю щитовидную костную пластинку; видоизмененная, утолщенная чешуя, часто килеватая или колючая; или выступающая модифицированная (грубая и сильно ребристая) шкала. Щитки обычно связаны с боковой линией , но могут быть обнаружены на хвостовом стебле (где они образуют хвостовые кили ) или вдоль вентрального профиля. Некоторые рыбы, например , сосновая шишка , полностью или частично покрыты щитками. Их функция предназначена для защиты рыб от воздействия окружающей среды и хищников других видов.
Боковая линия
[ редактировать ]
Боковая линия — это орган чувств , используемый для обнаружения движения и вибрации в окружающей воде. Например, рыбы могут использовать систему боковой линии, чтобы следовать за вихрями, создаваемыми убегающей добычей. У большинства видов он состоит из линии рецепторов, проходящей вдоль каждой стороны рыбы.
Подсвечники
[ редактировать ]Фотофоры — это светоизлучающие органы, которые у некоторых рыб выглядят как светящиеся пятна. Свет может вырабатываться соединениями во время переваривания добычи, специализированными митохондриальными клетками организма, называемыми фотоцитами , или симбиотическими бактериями . Фотофоры используются для привлечения пищи или спугивания хищников.
до
[ редактировать ]
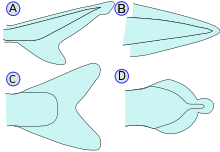


Плавники – самая отличительная черта рыб. Они либо состоят из костных шипов, либо лучей, выступающих из тела, покрытых кожей и соединяющих их вместе, либо в виде перепонок, как у большинства костистых рыб, либо похожих на ласты, как у акул. За исключением хвостового или хвостового плавников, плавники не имеют прямой связи с позвоночником и поддерживаются только мышцами. Их основная функция – помогать рыбе плавать. Плавники также можно использовать для скольжения или ползания, как это видно у летучей рыбы и рыбы-лягушки . Плавники, расположенные в разных местах рыбы, служат разным целям, например, движению вперед, повороту и сохранению вертикального положения. На каждый плавник приходится ряд видов рыб, у которых этот конкретный плавник был утерян в ходе эволюции. [ нужна ссылка ]
Шипы и лучи
[ редактировать ]У костистых рыб большинство плавников могут иметь шипы или лучи. Плавник может содержать только колючие лучи, только мягкие лучи или комбинацию того и другого. Если присутствуют оба, колючие лучи всегда располагаются спереди. Шипы обычно жесткие, острые и несегментированные. Лучи обычно мягкие, гибкие, сегментированные и могут быть разветвленными. Такая сегментация лучей является основным отличием их от шипов; У некоторых видов шипы могут быть гибкими, но никогда не сегментированными.
Шипы имеют разнообразное применение. У сома они используются как средство защиты; многие сомы обладают способностью выворачивать шипы наружу. Спинороги также используют шипы, чтобы запираться в расщелинах и не допускать их вытягивания.
Лепидотрихии представляют собой костные двустороннепарные сегментированные плавниковые лучи, встречающиеся у костистых рыб. Они развиваются вокруг актинотрихий как часть дермального экзоскелета. Лепидотрихии также могут содержать хрящи или кости. На самом деле они сегментированы и выглядят как серия дисков, сложенных друг на друга. Генетической основой формирования плавниковых лучей считаются гены, кодирующие белки актинодин 1 и актинодин 2 . [27]
Типы плавников
[ редактировать ]- Спинные плавники : Спинные плавники, расположенные на спине рыбы, служат для предотвращения перекачивания рыбы и помогают ей совершать резкие повороты и остановки. У большинства рыб один спинной плавник, но у некоторых рыб их два или три. У удильщиков передняя часть спинного плавника видоизменилась в иллиций и эску , биологический эквивалент удочки и приманки . Две-три кости, поддерживающие спинной плавник, называются проксимальным, средним и дистальным птеригиофорами . В остистых плавниках дистальный птеригиофор часто слит со средним или отсутствует вовсе.
- Хвостовой/хвостовой плавники: также называемые хвостовыми плавниками, хвостовые плавники прикреплены к концу хвостового стебля и используются для движения. Хвостовой стебель — узкая часть тела рыбы. Гипуральный сустав – это сустав между хвостовым плавником и последним из позвонков. Гипурал часто имеет веерообразную форму. Хвост может быть гетероцеркальным , обращенным гетероцеркальным, протоцеркальным , дифицеркальным или гомоцеркальным .
- Гетероцеркальный: позвонки доходят до верхней доли хвоста, делая его длиннее (как у акул).
- Перевернутая гетероцеркальность: позвонки заходят в нижнюю долю хвоста, делая его длиннее (как у Anaspida ).
- Протоцеркальный: позвонки доходят до кончика хвоста; хвост симметричен, но не расширен (как у круглоротых , предков позвоночных и ланцетников ).
- Дифицеркальный: позвонки доходят до кончика хвоста; хвост симметричен и расширен (как у бичира, двоякодышащей рыбы, миноги и целаканта ). Большинство палеозойских рыб имели дифицеркальный гетероцеркальный хвост. [28]
- Гомоцеркальный: позвонки простираются на очень короткое расстояние до верхней доли хвоста; хвост по-прежнему выглядит внешне симметричным. У большинства рыб имеется гомоцеркальный хвост, но он может иметь самую различную форму. Хвостовой плавник может быть закругленным на конце, усеченным (почти вертикальный край, как у лосося), раздвоенным (оканчивающимся двумя зубцами), выемчатым (с небольшим изгибом внутрь) или сплошным (с прикрепленными спинным, хвостовым и анальным плавниками). как у угрей).
- Анальные плавники : расположены на брюшной поверхности позади ануса и используются для стабилизации рыбы во время плавания.
- Грудные плавники : расположены парами с каждой стороны, обычно сразу за жаберной крышкой. Грудные плавники гомологичны передним конечностям четвероногих и помогают ходить некоторым видам рыб, например, некоторым удильщикам и илистым прыгунам . Своеобразной функцией грудных плавников, сильно развитой у некоторых рыб, является создание динамической подъемной силы, которая помогает некоторым рыбам, например акулам, сохранять глубину, а также обеспечивает «полет» летучим рыбам. Некоторые лучи грудных плавников могут превращаться в пальцеобразные выступы, как, например, у морских малиновок и летающих гурнардов .
- «Головные плавники»: «рога» скатов манты и их родственников, иногда называемые головными плавниками, на самом деле представляют собой модификацию передней части грудного плавника.
- Брюшные плавники : расположены парами на каждой стороне брюшной части ниже грудных плавников. Брюшные плавники гомологичны задним конечностям четвероногих. Они помогают рыбе подниматься или опускаться по воде, резко поворачиваться и быстро останавливаться. У бычков брюшные плавники часто слиты в один присосочный диск, который можно использовать для прикрепления к предметам.
- Жировой плавник : мягкий мясистый плавник, расположенный на спине позади спинного плавника и прямо перед хвостовым плавником. Он отсутствует во многих семействах рыб, но встречается у лососевых , харациновых и сомов. Его функция остается загадкой, и его часто отрезают, чтобы пометить рыбу, выращенную в инкубатории, хотя данные 2005 года показали, что форель с удаленным жировым плавником имеет на 8% более высокую частоту ударов хвостом. [29] Дополнительные исследования, опубликованные в 2011 году, показали, что плавник может иметь жизненно важное значение для обнаружения и реагирования на такие раздражители, как прикосновение, звук и изменения давления. Канадские исследователи обнаружили в плавнике нейронную сеть, что указывает на то, что она, вероятно, выполняет сенсорную функцию, но до сих пор не уверены, каковы будут последствия ее удаления. [30]
- Хвостовой киль: боковой гребень, обычно состоящий из щитков, на хвостовом стебле прямо перед хвостовым плавником. Встречается у некоторых видов быстро плавающих рыб. Он обеспечивает устойчивость и поддержку хвостового плавника, подобно килю корабля. Киль может быть одиночный парный, по одному с каждой стороны, или две пары сверху и снизу.
- Плавники : Маленькие плавники обычно находятся между спинным и хвостовым плавниками, но могут также находиться между анальным и хвостовым плавниками. У бичиров на спинной поверхности имеются только плавники, спинной плавник отсутствует. У некоторых рыб, таких как тунец или сайра , они лишены лучей, не убираются и находятся между последним спинным или анальным плавником и хвостовым плавником.
Внутренние органы
[ редактировать ]


Кишечник
[ редактировать ]Как и у других позвоночных, кишечник рыб состоит из двух сегментов: тонкого и толстого кишечника . У большинства высших позвоночных тонкая кишка делится на двенадцатиперстную кишку и другие части. У рыб отделы тонкой кишки не столь четкие, и термины «передняя кишка» или «проксимальная кишка» . вместо двенадцатиперстной кишки можно использовать [31] У костистых рыб кишечник относительно короткий, обычно примерно в полтора раза длиннее тела рыбы. Обычно он имеет несколько пилорических придатков , небольших мешообразных структур по всей длине, которые помогают увеличить общую площадь поверхности органа для переваривания пищи. У костистых рыб нет илеоцекального клапана , граница между тонкой кишкой и прямой кишкой обозначена только концом пищеварительного эпителия . [24] У некостистых рыб, таких как акулы, осетры и двоякодышащие рыбы, тонкого кишечника как такового нет. Вместо этого пищеварительная часть кишечника образует спиральную кишку , соединяющую желудок с прямой кишкой. При этом типе кишечника сама кишка относительно прямая, но имеет длинную складку, идущую по внутренней поверхности по спирали, иногда на десятки витков. Эта складка создает клапаноподобную структуру, которая значительно увеличивает как площадь поверхности, так и эффективную длину кишечника. Выстилка спиральной кишки аналогична слизистой оболочке тонкой кишки костистых и четвероногих немлекопитающих. [24] У миног спиральный клапан чрезвычайно мал, возможно, потому, что их диета требует незначительного переваривания. У миксины вообще нет спирального клапана, пищеварение происходит почти на всей длине кишечника, не подразделяющегося на разные отделы. [24]
Пилорический слепой
[ редактировать ]У многих рыб вдоль кишечника имеется ряд небольших отверстий, называемых пилорическими придатками. Цель слепой кишки — увеличить общую площадь поверхности кишечника, тем самым увеличивая всасывание питательных веществ. [32] [33]
Количество пилорических придатков сильно различается у разных видов, а у некоторых видов рыб придатки вообще отсутствуют. Виды с небольшим количеством слепых отростков или без них компенсируют их отсутствие за счет более длинного кишечника или более высоких или более извитых кишечных ворсинок, тем самым достигая аналогичных уровней площади поглощающей поверхности. [32] [33]
Двоякодышащие рыбы также имеют мешочек, расположенный в начале кишечника, который также называется пилорической слепой кишкой , но он имеет другую структуру и функцию, чем пилорический слепой отдел других видов рыб. Слепая кишка двоякодышащих рыб гомологична (из-за общего происхождения) слепой кишке, присутствующей у большинства амниот (четвероногих позвоночных, к которым относятся все млекопитающие, рептилии и птицы). [32] У большинства травоядных слепая кишка получает частично переваренную пищу из тонкой кишки и служит камерой ферментации для расщепления целлюлозы (например, травы или листьев) в рационе. У плотоядных слепая кишка часто сильно редуцирована или отсутствует.
Желудок
[ редактировать ]Как и у других позвоночных, относительное положение отверстий пищевода и двенадцатиперстной кишки по отношению к желудку остается относительно постоянным. В результате желудок всегда несколько изгибается влево, а затем поворачивается назад, чтобы встретиться с пилорическим сфинктером . Однако у миног, миксин, химер , двоякодышащих рыб и некоторых костистых рыб желудок вообще отсутствует, а пищевод открывается непосредственно в кишечник. Эти рыбы потребляют диеты, которые либо требуют небольшого хранения пищи, либо не требуют предварительного переваривания желудочными соками, либо и того, и другого. [34]
Почки
[ редактировать ]Почки рыб обычно представляют собой узкие, удлиненные органы, занимающие значительную часть туловища. Они сходны с мезонефросами высших позвоночных (рептилий, птиц и млекопитающих). Почки содержат группы нефронов , обслуживаемые собирательными трубочками, которые обычно впадают в мезонефральные протоки . Однако ситуация не всегда так проста. У хрящевых рыб имеется также более короткий проток, который дренирует задние (метанефрические) части почки и соединяется с мезонефрическим протоком у мочевого пузыря или клоаки. Действительно, у многих хрящевых рыб передняя часть почки может дегенерировать или вообще перестать функционировать у взрослой особи. [35] Почки миксины и миноги необычайно просты. Они состоят из ряда нефронов, каждый из которых впадает непосредственно в мезонефрический проток. [35] Как и у нильской тилапии, почка некоторых рыб состоит из трех частей; голова, туловище и хвостовые почки. [36] У рыб нет отдельного надпочечника с четко выраженной корой и мозговым слоем, как у млекопитающих. Интерренальные и хромаффинные клетки расположены в головной почке. [36]
Мочевой пузырь
[ редактировать ]Жабры большинства костистых рыб помогают выводить аммиак из организма, и рыбы живут в окружении воды, но у большинства из них все еще есть отдельный мочевой пузырь для хранения отработанной жидкости. Мочевой пузырь костистых рыб проницаем для воды, хотя это менее верно для пресноводных видов, чем для морских видов. [37] : с. 219 У пресноводных рыб мочевой пузырь является ключевым местом всасывания многих основных ионов. [38] у морских рыб моча удерживается в мочевом пузыре в течение длительного времени, чтобы максимизировать поглощение воды. [38] Считается, что мочевые пузыри рыб и четвероногих аналогичны, тогда как плавательные пузыри первых и легкие вторых считаются гомологичными.
У большинства рыб также есть орган, называемый плавательным пузырем , который не связан с мочевым пузырем, за исключением своей мембранной природы. Вьюны относятся к числу немногих видов рыб , , сардины и сельди у которых мочевой пузырь развит слабо. Он самый крупный у тех рыб, у которых отсутствует воздушный пузырь, и расположен перед яйцеводами и позади прямой кишки . [39]Селезенка
[ редактировать ]Селезенка имеется почти у всех позвоночных. Это нежизненно важный орган, по строению похожий на большой лимфатический узел . Он действует в первую очередь как фильтр крови и играет важную роль в отношении эритроцитов и иммунной системы . [40] У хрящевых и костных рыб он состоит в основном из красной мякоти и обычно представляет собой несколько удлиненный орган, поскольку фактически находится внутри серозной оболочки кишечника. [41] Единственные позвоночные, у которых нет селезенки, — это миноги и миксины. Даже у этих животных внутри стенки кишечника имеется диффузный слой кроветворной ткани, имеющий структуру, аналогичную красной пульпе , и предположительно гомологичную селезенке высших позвоночных. [41]
Печень
[ редактировать ]Печень — крупный жизненно важный орган, присутствующий у всех рыб. Он выполняет широкий спектр функций, включая детоксикацию , синтез белка и производство биохимических веществ, необходимых для пищеварения. Он очень восприимчив к загрязнению органическими и неорганическими соединениями, поскольку они могут накапливаться с течением времени и вызывать потенциально опасные для жизни состояния. Благодаря способности печени к детоксикации и хранению вредных компонентов ее часто используют в качестве биомаркера окружающей среды . [42]
Сердце
[ редактировать ]

Сердце у рыб часто называют двухкамерным. [43] состоит из одного предсердия для приема крови и одного желудочка для ее перекачки, [44] в отличие от трехкамерного (два предсердия, один желудочек) сердца земноводных и большинства рептилий и четырехкамерного (два предсердия, два желудочка) сердца млекопитающих и птиц. [43] Однако сердце рыбы имеет входное и выходное отделения, которые можно назвать камерами, поэтому его иногда называют трехкамерным. [44] или четырехкамерный, [45] в зависимости от того, что считать камерой. Предсердие и желудочек иногда считают «настоящими камерами», а остальные — «вспомогательными камерами». [46]
Четыре отсека расположены последовательно:
- Венозный синус : тонкостенный мешок или резервуар с сердечной мышцей, который собирает дезоксигенированную кровь через входящие печеночные и кардинальные вены . [ нужна проверка ] [44]
- Предсердие: толстостенная мышечная камера, которая направляет кровь в желудочек. [44]
- Желудочек: толстостенная мышечная камера, которая перекачивает кровь в четвертую часть — выносящий тракт. [44] Форма желудочка значительно варьирует: обычно трубчатая у рыб с удлиненным телом, пирамидальная с треугольным основанием у других, иногда мешковидная у некоторых морских рыб. [45]
- Путь оттока (OFT): Идет к брюшной части аорты и состоит из артериального трубчатого конуса , артериальной луковицы или обоих. [45] Артериальный конус, обычно встречающийся у более примитивных видов рыб, сокращается, обеспечивая приток крови к аорте, в то время как передняя луковица этого не делает. [46] [47]
Остиальные клапаны, состоящие из лоскутной соединительной ткани, предотвращают обратный ток крови через отсеки. [45] Устьевой клапан между венозным синусом и предсердием называется сино-предсердным клапаном, который закрывается во время сокращения желудочков. [45] Между предсердием и желудочком находится устьевой клапан, называемый атриовентрикулярным клапаном , а между артериальной луковицей и желудочком находится устьевой клапан, называемый бульбо-желудочковым клапаном. [45] Артериальный конус имеет различное количество полулунных клапанов . [46]
Вентральная аорта доставляет кровь к жабрам, где она насыщается кислородом и течет через дорсальную аорту в остальную часть тела. (У четвероногих брюшная аорта разделена на две части; одна половина образует восходящую аорту , а другая — легочную артерию ). [41]
Кровеносная система всех позвоночных животных закрытая . У рыб простейшая кровеносная система, состоящая всего из одного контура, при этом кровь перекачивается через капилляры жабр в капилляры тканей тела. Это известно как одноцикловая циркуляция. [48]
У взрослой рыбы четыре отсека расположены не в прямой ряд, а образуют S-образную форму, причем два последних отсека лежат над двумя первыми. Этот относительно более простой образец встречается у хрящевых рыб и лучепёрых рыб. У костистых рыб артериальный конус очень мал, и его точнее можно назвать частью аорты, а не собственно сердца. Артериальный конус отсутствует ни у каких амниот, предположительно он был поглощен желудочками в ходе эволюции. Точно так же, хотя у некоторых рептилий и птиц венозный синус присутствует в виде рудиментарной структуры, в противном случае он поглощен правым предсердием и больше не различим. [41]
Плавательный пузырь
[ редактировать ]
Плавательный пузырь или газовый пузырь — это внутренний орган, который способствует способности рыбы контролировать свою плавучесть и, таким образом, оставаться на текущей глубине воды, подниматься или опускаться, не тратя энергию на плавание. Мочевой пузырь имеется только у костистых рыб. У более примитивных групп, таких как некоторые Leuciscinae , bichirs и двоякодышащие рыбы, мочевой пузырь открыт пищеводу и выполняет функцию легкого . Он часто отсутствует у быстроплавающих рыб, таких как семейства тунцов и скумбрии. Рыбы с открытым пузырем, ведущим в пищевод, называются физостомами , а рыбы с закрытым пузырем — физоклистами . В последнем газообмен в мочевом пузыре контролируется посредством сети мирабилис — сети кровеносных сосудов, влияющих на газообмен между мочевым пузырем и кровью. [49]
Веберовский аппарат
[ редактировать ]Рыбы надотряда Ostariophys обладают структурой, называемой веберовским аппаратом , модификацией, которая позволяет им лучше слышать. Эта способность может объяснить заметный успех остариофизических рыб. [50] Аппарат состоит из набора костей, известных как веберовы косточки , цепочки мелких костей, которые соединяют слуховую систему с плавательным пузырем рыб. [51] Слуховые косточки соединяют стенку газового пузыря с Y-образным лимфатическим синусом , расположенным рядом с заполненным лимфой поперечным каналом, соединяющим мешочки правого и левого уха. Это позволяет передавать вибрации во внутреннее ухо. Полностью функционирующий веберовский аппарат состоит из плавательного пузыря, веберовских косточек, части переднего отдела позвоночника, а также некоторых мышц и связок. [51]
Репродуктивные органы
[ редактировать ]
К репродуктивным органам рыб относятся семенники и яичники . У большинства видов гонады представляют собой парные органы одинакового размера, которые могут частично или полностью слиться. [52] Также может существовать ряд вторичных органов, повышающих репродуктивную способность . Генитальный сосочек представляет собой небольшую мясистую трубку позади ануса у некоторых рыб, из которой выделяются сперматозоиды или яйца; пол рыбы часто можно определить по форме ее сосочка . [ нужна ссылка ] За определением пола у рыб, которое зависит от внутренних генетических факторов, следует половая дифференциация посредством экспрессии генов механизмов обратной связи, которые обеспечивают стабильность уровней определенных гормонов и клеточного профиля. Однако виды -гермафродиты являются исключением: они способны изменять ход половой дифференциации, чтобы максимизировать свою приспособленность. Существуют различные механизмы определения пола гонад у рыб и процессы, которые способствуют развитию функции гонад. На пол гонад влияет ряд факторов, включая клеточно-автономные генетические механизмы , эндокринные , паракринные , поведенческие или сигналы окружающей среды. Это приводит к тому, что первичные зародышевые клетки (ПГК) способны интерпретировать внутренние или внешние стимулы и развиваться в сперматогонии или оогонии . [53] Сперматогенез в семенниках — это процесс, при котором сперматогонии дифференцируются в сперматоциты посредством митоза и мейоза , в результате чего число хромосом уменьшается вдвое , образуя гаплоидные сперматиды . Во время спермиогенеза , последней стадии сперматогенеза, гаплоидные сперматиды развиваются в сперматозоиды . [54] В яичниках оогонии также подвергаются митозу и мейозу во время оогенеза , что приводит к образованию первичных ооцитов, а затем, в конечном итоге, к яйцеклетке . Первичный ооцит делится и дает вторичный ооцит, а также полярное тельце , прежде чем вторичный ооцит разовьется в гаплоидную отиду . [55]

Яички
[ редактировать ]
У большинства самцов рыб есть два семенника одинакового размера. У акул яичко на правой стороне обычно больше. У примитивных бесчелюстных рыб имеется лишь один семенник, расположенный по средней линии тела, хотя и он образуется в результате слияния парных структур в зародыше. [41]
Белая оболочка, под прочной перепончатой оболочкой семенник некоторых костистых рыб, содержит очень тонкие спиральные трубки, называемые семенными канальцами . Канальцы выстланы слоем клеток ( зародышевых клеток ), которые от полового созревания до старости развиваются в сперматозоиды (также известные как сперматозоиды или мужские гаметы ). Развивающиеся сперматозоиды проходят через семенные канальцы к сети яичка , расположенной в средостении яичка , к выносящим протокам , а затем к придатку яичка , где созревают вновь созданные сперматозоиды (см. Сперматогенез ). Сперматозоиды попадают в семявыносящий проток и в конечном итоге выбрасываются через уретру и из отверстия уретры посредством мышечных сокращений.
Однако у большинства рыб семенные канальцы отсутствуют. Вместо этого сперматозоиды производятся в сферических структурах, называемых ампулами сперматозоидов . Это сезонные структуры, выделяющие свое содержимое в период размножения, а затем реабсорбируемые организмом. Перед следующим сезоном размножения начинают формироваться и созревать новые ампулы сперматозоидов. В остальном ампулы по существу идентичны семенным канальцам высших позвоночных и включают тот же диапазон типов клеток. [57]
По распределению сперматогоний строение костистых семенников имеет два типа: у наиболее частых сперматогонии располагаются по всему ходу семенных канальцев, а у Atherinomorpha они приурочены к дистальной части этих структур. У рыб может наблюдаться кистозный или полукистозный сперматогенез. [ необходимо определение ] в отношении фазы выхода половых клеток кисты в просвет семенных канальцев. [52]
Яичники
[ редактировать ]Многие особенности яичников являются общими для всех позвоночных, включая наличие фолликулярных клеток и белочной оболочки. В яичнике рыбы в любой момент времени могут присутствовать сотни или даже миллионы оплодотворенных яиц. Свежие яйца могут развиваться из зародышевого эпителия на протяжении всей жизни. Желтые тела встречаются только у млекопитающих и у некоторых пластиножаберных рыб; у других видов остатки фолликула быстро рассасываются яичником. [57] Яичник костистых рыб часто содержит полое, заполненное лимфой пространство, которое открывается в яйцевод и куда откладываются яйца. [57] Большинство нормальных самок рыб имеют два яичника. У некоторых пластиножаберных полностью развивается только правый яичник. У примитивных бесчелюстных рыб и некоторых костистых рыб имеется только один яичник, образовавшийся в результате слияния парных органов в зародыше. [57]
Яичники рыб могут быть трех типов: гимнновариальные, вторично-гимновариальные и цистовариальные. При первом типе ооциты выходят непосредственно в целомическую полость, затем поступают в устье , затем через яйцевод и элиминируются. Вторичные гимноварианские яичники выделяют яйцеклетки в целом, откуда они попадают непосредственно в яйцевод. При третьем типе ооциты выводятся наружу через яйцевод. [58] Гимноварии — это примитивное состояние, встречающееся у двоякодышащих рыб, осетров и боуфинов . Кистовариумы характерны для большинства костистых рыб, у которых просвет яичника неразрывно связан с яйцеводом. [52] Вторичные гимноварии встречаются у лососевых и некоторых других костистых рыб.
Нервная система
[ редактировать ]
Центральная нервная система
[ редактировать ]У рыб обычно довольно маленький мозг по сравнению с размером тела по сравнению с другими позвоночными, обычно он составляет одну пятнадцатую массы мозга птицы или млекопитающего такого же размера. [59] Однако у некоторых рыб мозг относительно крупный, особенно у мормирид и акул, у которых мозг примерно такой же массивный по отношению к массе тела, как у птиц и сумчатых . [60]
Рыбий мозг разделен на несколько отделов. Спереди расположены обонятельные доли , пара структур, которые получают и обрабатывают сигналы от ноздрей через два обонятельных нерва . [59] Подобно тому, как люди чувствуют запах химикатов в воздухе, рыбы чувствуют запах химикатов в воде, пробуя их на вкус. Обонятельные доли очень велики у рыб, которые охотятся преимущественно по запаху, таких как миксины, акулы и сомы. За обонятельными долями находится двухлопастный конечный мозг , структурный эквивалент головного мозга у высших позвоночных. У рыб конечный мозг отвечает главным образом за обоняние . [59] Вместе эти структуры образуют передний мозг .
Передний мозг соединяется со средним мозгом через промежуточный мозг (на схеме эта структура находится ниже зрительных долей и поэтому не видна). Промежуточный мозг выполняет функции, связанные с гормонами и гомеостазом . [59] лежит Шишковидное тело чуть выше промежуточного мозга. Эта структура обнаруживает свет, поддерживает циркадные ритмы и контролирует изменения цвета. [59] Средний мозг или средний мозг содержит две зрительные доли . Они очень крупные у видов, которые охотятся визуально, таких как радужная форель и цихлиды . [59]
мозг Задний или метэнцефалон особенно участвует в плавании и равновесии. [59] Мозжечок представляет собой однодольную структуру, которая обычно является самой большой частью мозга. [59] Микины и миноги имеют относительно небольшой мозжечок, в то время как мозжечок мормирид массивен и, по-видимому, участвует в их электрическом чувстве . [59]
Ствол мозга или продолговатый мозг — это задняя часть мозга. [59] Помимо контроля над некоторыми мышцами и органами тела, по крайней мере у костистых рыб, ствол мозга управляет дыханием и осморегуляцией . [59]
Позвоночные животные — единственная группа хордовых, обладающая полноценным мозгом. У ланцетника наблюдается небольшое вздутие переднего конца спинного нервного канатика , однако у него отсутствуют глаза и другие сложные органы чувств, сравнимые с таковыми у позвоночных. У других хордовых не наблюдается тенденции к цефализации . [9] Центральная нервная система основана на полой нервной трубке, проходящей по всей длине животного, от которой периферическая нервная система, разветвляется иннервирующая различные системы. Передний конец нервной трубки расширен за счет утолщения стенок и расширения центрального канала спинного мозга на три первичных мозговых пузырька; проэнцефалон ромбенцефалон (передний мозг), средний мозг (средний мозг) и ( задний мозг) затем далее дифференцировались в различных группах позвоночных. [61] Два латерально расположенных глаза формируются вокруг выростов среднего мозга, за исключением миксины, хотя это может быть вторичной потерей. [62] [63] Передний мозг хорошо развит и подразделен у большинства четвероногих, тогда как средний мозг доминирует у многих рыб и некоторых саламандр . Везикулы переднего мозга обычно парные, образуя полушария, подобные полушариям головного мозга у млекопитающих. [61] Полученная в результате анатомия центральной нервной системы с единственным полым вентральным нервным тяжем, увенчанным рядом (часто парных) пузырьков, уникальна для позвоночных. [9]
Мозжечок
[ редактировать ]Цепи мозжечка одинаковы у всех классов позвоночных, включая рыб, рептилий, птиц и млекопитающих. [64] Аналогичное строение мозга имеется и у головоногих моллюсков с хорошо развитым мозгом, например у осьминогов . [65] Это было воспринято как доказательство того, что мозжечок выполняет функции, важные для всех видов животных, имеющих мозг.
У разных видов позвоночных существуют значительные различия в размерах и форме мозжечка. У земноводных, миног и миксин мозжечок развит слабо; в последних двух группах он едва отличим от ствола мозга. Хотя спино-мозжечок в этих группах присутствует , первичными структурами являются небольшие парные ядра, соответствующие вестибуло-мозжечку . [57]
Мозжечок хрящевых и костных рыб необычайно велик и сложен. По крайней мере, в одном важном отношении он отличается по внутренней структуре от мозжечка млекопитающих: мозжечок рыб не содержит дискретных глубоких ядер мозжечка . Вместо этого основной мишенью клеток Пуркинье являются клетки особого типа, распределенные по коре мозжечка, тип, не встречающийся у млекопитающих. У мормирид (семейство слабоэлектрочувствительных пресноводных рыб) мозжечок значительно больше, чем весь остальной мозг вместе взятый. Самая большая его часть представляет собой специальную структуру, называемую клапаном , которая имеет необычно правильную архитектуру и получает большую часть информации от электросенсорной системы. [66]
У большинства видов рыб и амфибий есть система боковой линии, которая улавливает волны давления в воде. Одна из областей мозга, которая получает первичную информацию от органа боковой линии, медиальное октаволатеральное ядро, имеет структуру, подобную мозжечку, с зернистыми клетками и параллельными волокнами. У электрочувствительных рыб сигнал от электросенсорной системы поступает в дорсальное октаволатеральное ядро, которое также имеет структуру, подобную мозжечку. У лучепёрых рыб (самая большая группа) в зрительной тектуме имеется слой — маргинальный слой, похожий на мозжечок. [64]
Идентифицированные нейроны
[ редактировать ]Нейрон считается «идентифицированным», если он обладает свойствами, которые отличают его от любого другого нейрона того же животного — такими свойствами, как местоположение, нейромедиатор , характер экспрессии генов и связность — и если каждый отдельный организм, принадлежащий к тому же виду, имеет один и только один нейрон с одинаковым набором свойств. [67] В нервных системах позвоночных в этом смысле «идентифицировано» очень мало нейронов (считается, что у человека их нет). Таким образом, в более простых нервных системах некоторые или все нейроны могут быть уникальными. [68]
У позвоночных наиболее известными нейронами являются гигантские клетки Маутнера рыб. [69] У каждой рыбы есть две клетки Маутнера, расположенные в нижней части ствола мозга: одна слева и одна справа. Каждая клетка Маутнера имеет аксон , который пересекается, иннервируя нейроны на одном и том же уровне мозга, а затем перемещаясь вниз через спинной мозг, образуя при этом многочисленные связи. Синапсы , генерируемые клеткой Маутнера, настолько мощны, что один-единственный потенциал действия вызывает серьезную поведенческую реакцию: в течение миллисекунд рыба изгибает свое тело в форме буквы С , затем выпрямляется, тем самым быстро продвигаясь вперед. Функционально это реакция быстрого побега , которую легче всего вызвать сильная звуковая волна или волна давления, воздействующая на орган боковой линии рыбы. Клетки Маутнера — не единственные идентифицированные нейроны у рыб: существует еще около 20 типов, включая пары «аналогов клеток Маутнера» в каждом сегментарном ядре спинного мозга. Хотя клетка Маутнера способна сама по себе вызывать реакцию бегства, в контексте обычного поведения другие типы клеток обычно способствуют формированию амплитуды и направления реакции.
Клетки Маутнера были описаны как командные нейроны . Командный нейрон — это особый тип идентифицированного нейрона, определяемый как нейрон, который способен самостоятельно управлять определенным поведением. [70] Такие нейроны чаще всего появляются в системах быстрого ускользания у различных видов: гигантский аксон кальмара и гигантский синапс кальмара , используемые для новаторских экспериментов в нейрофизиологии из-за их огромного размера, оба участвуют в цепи быстрого ускользания кальмара. Однако концепция командного нейрона стала спорной из-за исследований, показавших, что некоторые нейроны, которые изначально подходили под это описание, на самом деле были способны вызывать реакцию только в ограниченном наборе обстоятельств. [71]
Иммунная система
[ редактировать ]Иммунные органы различаются в зависимости от вида рыб. [72] У бесчелюстных рыб (миноги и миксины) настоящие лимфоидные органы отсутствуют. Эти рыбы полагаются на участки лимфоидной ткани в других органах для производства иммунных клеток. Например, эритроциты , макрофаги и плазматические клетки производятся в передней части почки (или пронефросе ) и некоторых участках кишечника (где созревают гранулоциты ). Они напоминают примитивный костный мозг миксины.
Хрящевые рыбы (акулы и скаты) имеют более развитую иммунную систему. У них есть три специализированных органа, уникальные для хондрихтиев ; эпигональные органы (лимфоидные ткани, похожие на кости млекопитающих), окружающие гонады, орган Лейдига в стенках пищевода и спиральный клапан в кишечнике. В этих органах находятся типичные иммунные клетки (гранулоциты, лимфоциты и плазматические клетки). У них также есть узнаваемый тимус и хорошо развитая селезенка (их самый важный иммунный орган), где различные лимфоциты развиваются и хранятся , плазматические клетки и макрофаги.
Хондростальные рыбы (осетры, веслоносы и бичиры) обладают основным местом производства гранулоцитов в массе, которая связана с мозговыми оболочками , мембранами, окружающими центральную нервную систему. Их сердце часто покрыто тканью, содержащей лимфоциты, ретикулярные клетки и небольшое количество макрофагов. Хрящевая почка — важный кроветворный орган; именно здесь развиваются эритроциты, гранулоциты, лимфоциты и макрофаги.
Как и у хондростеевых рыб, основные иммунные ткани костистых рыб ( костистых рыб ) включают почки (особенно переднюю почку), в которых находится множество различных иммунных клеток. [73] Кроме того, костистые рыбы обладают тимусом, селезенкой и рассеянными иммунными участками в тканях слизистых оболочек (например, в коже, жабрах, кишечнике и половых железах). Как и в случае с иммунной системой млекопитающих, костистые эритроциты, нейтрофилы и гранулоциты, как полагают, находятся в селезенке, тогда как лимфоциты являются основным типом клеток, обнаруженных в тимусе. [74] [75] В 2006 году лимфатическая система, аналогичная лимфатической системе млекопитающих, была описана у одного вида костистых рыб — данио . Хотя это еще не подтверждено, в этой системе, по-видимому, будут накапливаться нестимулированные наивные Т-клетки в ожидании встречи с антигеном . [76]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Проссер, К. Лэдд (18 марта 1991 г.). «Введение: Определение сравнительной физиологии: теория адаптации». В Проссере, К. Лэдд (ред.). Экологическая и метаболическая физиология животных . Нью-Йорк: Вили-Лисс. стр. 1–2. ISBN 0-471-85767-Х . ОСЛК 22906165 . Архивировано из оригинала 3 июля 2021 года . Проверено 3 июля 2021 г.
- ^ Jump up to: а б Дорит, РЛ; Уокер, ВФ; Барнс, Р.Д. (1991). Зоология . Издательство Колледжа Сондерса. стр. 816–818 . ISBN 978-0-03-030504-7 .
- ^ «Рыбье сердце» . ThinkQuest . Оракул. Архивировано из оригинала 28 апреля 2012 года . Проверено 27 июня 2013 г.
- ^ Jump up to: а б Котпал, Р.Л. (2010). Современный учебник зоологии: Позвоночные животные . Публикации Растоги. п. 193. ИСБН 9788171338917 . Архивировано из оригинала 22 апреля 2016 года.
- ^ МакГиннис, Сэмюэл М. (2006). Полевой справочник по пресноводным рыбам Калифорнии. Архивировано 1 августа 2020 г. на странице 45 Wayback Machine , University of California Press. ISBN 9780520936966
- ^ Ваггонер, Бен. «Позвоночные животные: летопись окаменелостей» . УКМП. Архивировано из оригинала 29 июня 2011 года . Проверено 15 июля 2011 г.
- ^ Бертон, Дерек; Бертон, Маргарет (21 декабря 2017 г.). Основная биология рыб . Том. 1. Издательство Оксфордского университета. дои : 10.1093/oso/9780198785552.001.0001 . ISBN 978-0-19-878555-2 .
- ^ Ваггонер, Бен. «Позвоночные: еще о морфологии» . УКМП. Архивировано из оригинала 6 августа 2012 года . Проверено 13 июля 2011 г.
- ^ Jump up to: а б с Ромер, А.С. (1949): Тело позвоночного. У. Б. Сондерс, Филадельфия. (2-е изд. 1955 г.; 3-е изд. 1962 г.; 4-е изд. 1970 г.)
- ^ Лием, Карел Ф.; Уоррен Франклин Уокер (2001). Функциональная анатомия позвоночных: эволюционная перспектива . Издательство Харкорт-колледжа. п. 277. ИСБН 978-0-03-022369-3 .
- ^ Jump up to: а б с д Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 161–170. ISBN 978-0-03-910284-5 .
- ^ Кураку; Хошияма, Д; Като, К; Шуга, Х; Мията, Т; и др. (декабрь 1999 г.). «Монофилия миног и миксин, поддерживаемая генами, закодированными в ядерной ДНК». Журнал молекулярной эволюции . 49 (6): 729–35. Бибкод : 1999JMolE..49..729K . дои : 10.1007/PL00006595 . ПМИД 10594174 . S2CID 5613153 .
- ^ Николлс, Генри (10 сентября 2009 г.). «Из уст в уста» . Природа . 461 (7261): 164–166. дои : 10.1038/461164а . ПМИД 19741680 .
- ^ Сток, Дэвид; Уитт Г.С. (7 августа 1992 г.). «Свидетельства на основе последовательностей рибосомальной РНК 18S о том, что миноги и миксины образуют естественную группу». Наука . 257 (5071): 787–9. Бибкод : 1992Sci...257..787S . дои : 10.1126/science.1496398 . ПМИД 1496398 .
- ^ Jump up to: а б с д Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 173–177. ISBN 978-0-03-910284-5 .
- ^ Мюллер, М. (1996). «Новая классификация плоских четырехстержневых связей и ее применение к механическому анализу систем животных» (PDF) . Фил. Пер. Р. Сок. Лонд. Б. 351 (1340): 689–720. Бибкод : 1996РСТБ.351..689М . дои : 10.1098/rstb.1996.0065 . ПМИД 8927640 . Архивировано (PDF) из оригинала 27 сентября 2011 года . Проверено 11 января 2011 г.
- ^ Н. А. Кэмпбелл и Дж. Б. Рис (2005). Биология , седьмое издание. Бенджамин Каммингс, Сан-Франциско, Калифорния.
- ^ Тревор Д. Лэмб; Шон П. Коллин; Эдвард Н. Пью младший (2007). «Эволюция глаза позвоночных: опсины, фоторецепторы, сетчатка и глазная чашечка» . Обзоры природы Неврология . 8 (12): 960–976. дои : 10.1038/nrn2283 . ПМК 3143066 . ПМИД 18026166 .
- ^ Хелфман, Коллетт, Фейси и Боуэн, 2009, Разнообразие рыб: биология, эволюция и экология, стр. 84–87.
- ^ «Покровы — Членистоногие | Британника» . www.britanica.com . Проверено 6 мая 2022 г.
- ^ Jump up to: а б Кардонг, Кеннет (2018). Сравнительная анатомия, функции, эволюция позвоночных . Нью-Йорк, Нью-Йорк: McGraw-Hill Education. стр. 212–219. ISBN 978-1-259-70091-0 .
- ^ Рейкерс, Себастьян; Геберт, Марина; Уппалапати, Сай; Мейер, Уилфрид; Мэдерсон, Пол; Селл, Энн Ф.; Крузе, Чарли; Паус, Ральф (2010). « Рыба имеет значение: актуальность биологии рыбьей кожи для исследований в дерматологии» . Экспериментальная дерматология . 19 (4): 313–324. дои : 10.1111/j.1600-0625.2009.01059.x . ПМИД 20158518 . S2CID 20223479 .
- ^ «Покровы - Производные кожи и придатки | Британника» . www.britanica.com . Проверено 20 апреля 2022 г.
- ^ Jump up to: а б с д Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 129–145. ISBN 978-0-03-910284-5 .
- ^ "Покровы - Рыбы | Британика" . www.britanica.com . Проверено 20 апреля 2022 г.
- ^ Кардонг, Кеннет (2018). Сравнительная анатомия, функции, эволюция позвоночных . Нью-Йорк, Нью-Йорк: McGraw-Hill Education. стр. 212–219. ISBN 978-1-259-70091-0 .
- ^ Чжан, Дж.; Ваг, П.; Гуай, Д.; Санчес-Пулидо, Л.; Падхи, Британская Колумбия; Корж, В.; Андраде-Наварро, Массачусетс; Акименко, М.А. (2010). «Потеря белков актинотрихии рыб и переход от плавника к конечностям». Природа . 466 (7303): 234–237. Бибкод : 2010Natur.466..234Z . дои : 10.1038/nature09137 . ПМИД 20574421 . S2CID 205221027 .
- ^ фон Зиттель К.А., Вудворд А.С. и Шлозер М. (1932) Учебник палеонтологии, том 2, Macmillan and Company. Страница 13.
- ^ Тителл, Э. (2005). «Таинственный маленький толстый плавник» . Журнал экспериментальной биологии . 208 : v – vi. дои : 10.1242/jeb.01391 . Архивировано из оригинала 26 июля 2008 года . Проверено 8 февраля 2011 г.
- ^ «Удаление форели, плавник лосося задевает нерв» . Архивировано из оригинала 20 июля 2011 года.
- ^ Гийом, Жан; Издательство Праксис; Садасивам Кошик; Пьер Берго; Роберт Метайллер (2001). Питание и кормление рыб и ракообразных . Спрингер. п. 31. ISBN 978-1-85233-241-9 . Архивировано из оригинала 14 апреля 2021 года . Проверено 9 января 2009 г.
- ^ Jump up to: а б с Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 353–354. ISBN 978-0-03-910284-5 .
- ^ Jump up to: а б Баддингтон, РК; Даймонд, Дж. М. (1986). «Возвращение к Аристотелю: функция пилорических придатков у рыб» (PDF) . Учеб. Натл. акад. наук. США . 83 (20): 8012–8014. Бибкод : 1986PNAS...83.8012B . дои : 10.1073/pnas.83.20.8012 . ПМЦ 386855 . ПМИД 3464017 . Архивировано (PDF) из оригинала 23 мая 2018 года . Проверено 22 мая 2018 г.
- ^ Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 345–349. ISBN 978-0-03-910284-5 .
- ^ Jump up to: а б Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 367–376. ISBN 978-0-03-910284-5 .
- ^ Jump up to: а б Габер и Абдель-Масуд, Вафаа и Фатма (2019). «Интерренальная ткань, хромаффинные клетки и тельца станния нильского тилапии (Oreochromis niloticus)» . Микроскопия (Оксфорд, Англия) . 68 (3): 195–206. doi : 10.1093/jmicro/dfy146 . ПМИД 30805608 . Архивировано из оригинала 26 апреля 2021 года . Проверено 26 апреля 2021 г.
- ^ Пи Джей Бентли (14 марта 2013 г.). Эндокринная система и осморегуляция: сравнительный анализ позвоночных . Springer Science & Business Media. ISBN 978-3-662-05014-9 .
- ^ Jump up to: а б Таквам, Мариус; Вуд, Крис М.; Криви, Х.; Нильсен, Том О. (29 июня 2023 г.). «Роль почек в регуляции кислотно-щелочного баланса и выделении аммиака у пресноводных и морских рыб: последствия нефрокальциноза» . Границы в физиологии . 14 . дои : 10.3389/fphys.2023.1226068 . ISSN 1664-042X . ПМЦ 10339814 . ПМИД 37457024 .
- ^ Оуэн, Ричард (1843). Лекции по сравнительной анатомии и физиологии беспозвоночных животных . Лондон: Лонгман, Браун, Грин и Лонгманс. стр. 283–284.
- ^ Селезенка. Архивировано 31 мая 2019 г. в Wayback Machine , Интернет-энциклопедия науки.
- ^ Jump up to: а б с д и Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия: Холт-Сондерс Интернэшнл. стр. 410–411. ISBN 978-0-03-910284-5 .
- ^ Стори, Э.М.; Роча, MLCF; Диас, Дж. Ф.; дос Сантос, CEI; де Соуза, Коннектикут; Амарал, Л.; Диас, Дж. Ф. (1 января 2014 г.). «Элементарная характеристика повреждений печени рыб» . Ядерные приборы и методы в физических исследованиях. Раздел B: Взаимодействие пучков с материалами и атомами . 318 : 83–87. Бибкод : 2014НИМПБ.318...83С . дои : 10.1016/j.nimb.2013.05.109 . ISSN 0168-583X . Архивировано из оригинала 3 июля 2021 года . Проверено 3 июля 2021 г. - через Elsevier Science Direct.
- ^ Jump up to: а б Джурд, Ричард Дэвид (январь 2004 г.). Мгновенные заметки по биологии животных . Гирляндная наука. п. 134. ИСБН 978-1-85996-325-8 . Архивировано из оригинала 6 декабря 2016 года . Проверено 13 марта 2016 г.
- ^ Jump up to: а б с д и Острандер, Гэри Кент (2000). Лабораторная рыбка . Эльзевир. стр. 154–155. ISBN 978-0-12-529650-2 . Архивировано из оригинала 6 декабря 2016 года . Проверено 13 марта 2016 г.
- ^ Jump up to: а б с д и ж Фаррелл, Энтони П., изд. (1 июня 2011 г.). Энциклопедия физиологии рыб: от генома к окружающей среде . Стивенс, Э. Дон; Чех-младший, Джозеф Дж; Ричардс, Джеффри Г. Academic Press. п. 2315. ИСБН 978-0-08-092323-9 . Архивировано из оригинала 6 декабря 2016 года . Проверено 13 марта 2016 г.
- ^ Jump up to: а б с Шукла, JP Fish & Fisheries . Публикации Растоги. стр. 154–155. ISBN 978-81-7133-800-9 . Архивировано из оригинала 6 декабря 2016 года . Проверено 13 марта 2016 г.
- ^ Икардо, Хосе М. (2006). «Артериальный конус костистого сердца: исключен, но не пропущен» . Анатомические записи, часть A: открытия в молекулярной, клеточной и эволюционной биологии . 288А (8): 900–908. дои : 10.1002/ar.a.20361 . ISSN 1552-4884 . ПМИД 16835938 . S2CID 9676359 .
- ^ Гилберт, Скотт Ф. (1994). Биология развития (4-е изд.). Сандерленд, Массачусетс: Sinauer Associates, Inc., стр. 781 . ISBN 978-0-87893-249-8 .
- ^ Кардонг, К. (2008). Позвоночные животные: Сравнительная анатомия, функции, эволюция (5-е изд.). Бостон: МакГроу-Хилл. ISBN 978-0-07-304058-5 .
- ^ Бриггс, Джон К. (2005). «Биогеография рыб-отофизов (Ostariophy: Otophys): новая оценка». Журнал биогеографии . 32 (2): 287–294. Бибкод : 2005JBiog..32..287B . дои : 10.1111/j.1365-2699.2004.01170.x . S2CID 84010604 .
- ^ Jump up to: а б Нельсон, Джозеф, С. (2006). Рыбы мира . Джон Уайли и сыновья, Inc. ISBN 978-0-471-25031-9 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б с Гимарайнш-Крус, Родриго Х., Родриго Х.; Сантос, Хосе Э. дос; Сантос, Гилмар Б. (2005). «Структура гонад и гаметогенез Loricaria lentiginosa Isbrücker (Pisces, Teleostei, Siluriformes)» . Преподобный Брас. Зоол . 22 (3): 556–564. дои : 10.1590/S0101-81752005000300005 . ISSN 0101-8175 .
- ^ Девлин, Роберт Х.; Нагахама, Ёситака (21 июня 2002 г.). «Определение пола и дифференциация пола у рыб: обзор генетических, физиологических и экологических влияний» . Аквакультура . Определение пола и половая дифференциация у рыб. 208 (3): 191–364. Бибкод : 2002Aquac.208..191D . дои : 10.1016/S0044-8486(02)00057-1 . ISSN 0044-8486 .
- ^ Нисимура, Хитоши; Л'Эрно, Стивен В. (25 сентября 2017 г.). «Сперматогенез» . Современная биология . 27 (18): Р988–Р994. Бибкод : 2017CBio...27.R988N . дои : 10.1016/j.cub.2017.07.067 . ISSN 0960-9822 . ПМИД 28950090 . S2CID 235311767 .
- ^ Санчес, Флор; Смитц, Йохан (1 декабря 2012 г.). «Молекулярный контроль оогенеза» . Biochimica et Biophysica Acta (BBA) - Молекулярные основы болезней . Молекулярная генетика репродуктивной недостаточности человека. 1822 (12): 1896–1912. дои : 10.1016/j.bbadis.2012.05.013 . ISSN 0925-4439 . ПМИД 22634430 .
- ^ Араужо, Андреа Соарес де; ду Насименто, Уоллес Силва; Ямамото, Мария Эмилия; Челлаппа, Сатьябама (2012). «Временная динамика размножения неотропических рыб Crenicichla menezesi (Perciformes: Cichlidae)» . Научный мировой журнал . 2012 . Хиндави Лимитед: 1–10. дои : 10.1100/2012/579051 . ISSN 1537-744X . ПМК 3415153 . ПМИД 22919339 .
- ^ Jump up to: а б с д и Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 385–386. ISBN 978-0-03-910284-5 .
- ^ Брито, МФГ; Базцоли, Н. (2003). «Размножение сома-сурубима (Pisces, Pimelodidae) в реке Сан-Франциско, регион Пирапора, Минас-Жерайс, Бразилия» . Бразильский архив ветеринарной медицины и зоотехники . 55 (5): 624–633. дои : 10.1590/S0102-09352003000500018 . ISSN 0102-0935 .
- ^ Jump up to: а б с д и ж г час я дж к Хелфман, Коллетт и Фейси, 1997 , стр. 48–49.
- ^ Хелфман, Коллетт и Фейси 1997 , стр. 191
- ^ Jump up to: а б Хильдебранд М. и Гонслоу Г. (2001): Анализ структуры позвоночных. 5-е издание. Джон Уайли и сыновья, Inc. Нью-Йорк
- ^ «Следить за эволюцией» . PhysOrg.com . 3 декабря 2007 г. Архивировано из оригинала 15 марта 2012 г. Проверено 4 декабря 2007 г.
- ^ «Гиперотрети – Микины» . Архивировано из оригинала 6 февраля 2013 года . Проверено 14 декабря 2012 г.
- ^ Jump up to: а б Bell CC, Хан В., Sawtell NB (2008). «Структуры, подобные мозжечку, и их значение для функции мозжечка». Анну. Преподобный Нейроски . 31 : 1–24. дои : 10.1146/annurev.neuro.30.051606.094225 . ПМИД 18275284 .
- ^ Вудхэмс PL (1977). «Ультраструктура аналога мозжечка у осьминога». Джей Комп Нейрол . 174 (2): 329–45. дои : 10.1002/cne.901740209 . ПМИД 864041 . S2CID 43112389 .
- ^ Ши З, Чжан Ю, Мик Дж, Цяо Дж, Хан ВЗ (2008). «Нейрональная организация уникальной специализации мозжечка: клапан мозжечка мормиридовой рыбы» . Дж. Комп. Нейрол . 509 (5): 449–73. дои : 10.1002/cne.21735 . ПМЦ 5884697 . ПМИД 18537139 .
- ^ Хойл Дж., Виерсма, Калифорния (1977). Идентифицированные нейроны и поведение членистоногих . Пленум Пресс. ISBN 978-0-306-31001-0 .
- ^ «Червячная книга: Спецификация нервной системы» . Архивировано из оригинала 17 июля 2011 года . Проверено 14 декабря 2012 г.
- ^ Штейн ПСЖ (1999). Нейроны, сети и двигательное поведение . МТИ Пресс. стр. 38–44. ISBN 978-0-262-69227-4 .
- ^ Штейн, с. 112
- ^ Симмонс П.Дж., Янг Д. (1999). Нервные клетки и поведение животных . Издательство Кембриджского университета. п. 43. ИСБН 978-0-521-62726-9 .
- ^ Сапата, Агустин Г.; Чиба, Акира; Варас, Альберто (1996). «Клетки и ткани иммунной системы рыб». Организм, патоген и окружающая среда . Физиология рыб. Том. 15. стр. 1–62. дои : 10.1016/s1546-5098(08)60271-x . ISBN 9780123504395 .
- ^ Д. П. Андерсон. Иммунология рыб . (С. Ф. Снешко и Х. Р. Аксельрод, ред.), Гонконг: TFH Publications, Inc. Ltd., 1977.
- ^ Чильмончик, С. (1992). «Тимус рыб: развитие и возможная функция в иммунном ответе». Ежегодный обзор болезней рыб . 2 : 181–200. дои : 10.1016/0959-8030(92)90063-4 .
- ^ Хансен, доктор юридических наук; Сапата, АГ (1998). «Развитие лимфоцитов у рыб и земноводных». Иммунологические обзоры . 166 : 199–220. дои : 10.1111/j.1600-065x.1998.tb01264.x . ПМИД 9914914 . S2CID 7965762 .
- ^ Кучер; и др. (2006). «Развитие лимфатической системы рыбок данио требует передачи сигналов VegFc» . Современная биология . 16 (12): 1244–1248. Бибкод : 2006CBio...16.1244K . дои : 10.1016/j.cub.2006.05.026 . ПМИД 16782017 . S2CID 428224 .
Цитируемые работы
[ редактировать ]- Хельфман, Г.; Коллетт, Б.; Фейси, Д. (1997). Разнообразие рыб (1-е изд.). Уайли-Блэквелл. ISBN 978-0-86542-256-8 .
Внешние ссылки
[ редактировать ]- Mongabay.com Анатомия рыб Mongabay
- Потрясающие рентгеновские снимки рыб , выставка Смитсоновского института, LiveScience , 13 июня 2011 г.
| Поля |
|  |
|---|---|---|
| Бактерии и грибы | ||
| Протисты | ||
| Растения | ||
| Беспозвоночные | ||
| Млекопитающие | ||
| Другие позвоночные | ||
| Глоссарии | ||
| Связанные темы | ||
